Мішені дії антивірусних агентів
З розвитком сучасної молекулярної вірусології та ДНК- рекомбінантних технологій метод сліпого скринінгу активних сполук був частково відкинутий. Важливі для вірусного росту гени були клоновані та експресовані в транскрипційно- здатних організмах, тому їх продукти можуть бути проаналізовані на молекулярному рівні.
Життєвий цикл багатьох вірусів добре вивчений, що виявило багато мішеней для дії антивірусних сполук (табл. 5.5). За допомогою вірусних маркерів і ДНК-рекомбінантних технологій інгібітори важливих вірусологічних процесів можуть бути знайдені, навіть якщо вірус не реплікується в культурі тканин.Таблиця 5.5
Основні мішені дії антивірусних препаратів
| Мішень | Прототип препарату |
| Інтактний віріон | Оксалін |
| Взаємодія з клітинними рецепторами Злиття мембран | Аналоги рецепторів Енфувіртид |
| Депротеїнізація вірусної НК Первинний синтез РНК у РНК-вірусів Зворотна транскрипція | Амантадин Інгібітори РНК-залежної РНК-полімерази Зидовудин |
| Інтеграція | Ралтегравір |
| Регуляція синтезу РНК | ВІЛ-/Щ-інгібітори |
| Процесинг РНК-транскриптів | Рибавірін |
| Трансляція вірусної РНК | Інтерферон |
| Саквінавір | |
| Реплікація ДНК-вірусів | Ацикловір |
| Реплікація РНК-вірусів | РНК-залежна РНК-полімераза, інгібітори хеліказ |
У результаті багаторічного пошуку антивірусних хіміопрепаратів установлено, що найкращі з них або безпосередньо селективно інгібують окремі етапи вірусного репродукування в клітинах, або діють опосередковано через клітину-хазяїна.
Препарати першого типу впливають на проникнення вірусу в клітину та його депротеїнізацію, транскрипцію вірусного геному, синтез або функціонування вірусних ДНК, РНК та білків. У другому випадку йдеться про речовини, що впливають на структури клітини, які відіграють важливу роль при репродукуванні вірусу. До них також можна віднести речовини, що діють на заражені вірусом клітини й викликають їх елімінацію з організму.
Інгібітори проникнення вірусів до клітини. На перших кроках реплікації вірус є привабливою мішенню для антивірус - ної дії, оскільки його взаємодія з клітинним рецептором високо- специфічна. Сполуки, які можуть інтерферувати в процесі взаємодії вірусу з клітинним рецептором, не повинні впливати на нормальне функціонування клітинних рецепторів. Стадія взаємодії вірусу з клітиною може бути заблокована двома шляхами (рис. 5.9, 5.10):
1) використанням агентів, які імітують вірусасоційовані протеїни та специфічно зв'язуються з клітинними рецепторами (антитіла, антиідіотипові антитіла, природні й синтетичні ліганди);
2) використанням агентів, що імітують рецептор і специфічно взаємодіють із вірусасоційованими протеїнами (антитіла до вірусних білків, антиідіотипові антитіла, синтетичні рецептори та аналоги рецепторів).
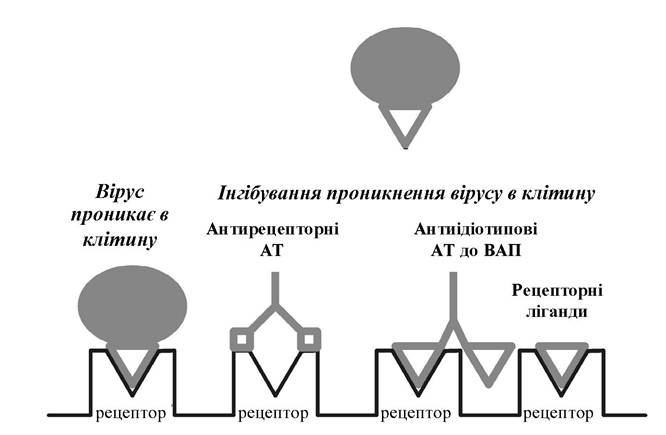
Рис. 5.9. Способи інгібування взаємодії вірусу з клітиною за допомогою агентів, які впливають на клітинний рецептор (АТ - антитіла; ВАП - вірусасоційовані протеїни)
Для багатьох вірусів, зокрема герпесвірусів та ВІЛ, розроблені рецептори, які блокують взаємодію вірус-рецептор. Для вірусів, які мають суперкапсидну оболонку, злиття мембран є наступною мішенню антивірусної дії. За допомогою комп'ютерного моделювання були ідентифіковані маленькі молекули, що можуть зв'язуватися з гідрофобним карманом гемаглютиніну вірусу грипу, тим самим перешкоджаючи конформаційним змінам гемаглютиніну, які передують злиттю мембран. Із серії запропонованих сполук деякі похідні бензохінону та гідрохінону виявили інгібуючу активність на процес злиття мембран.
Механізм дії таких сполук залишається невідомим, але дослідники схиляються до думки, що ці речовини діють як молекулярний клей, який перешкоджає перебудові фузійного пептиду.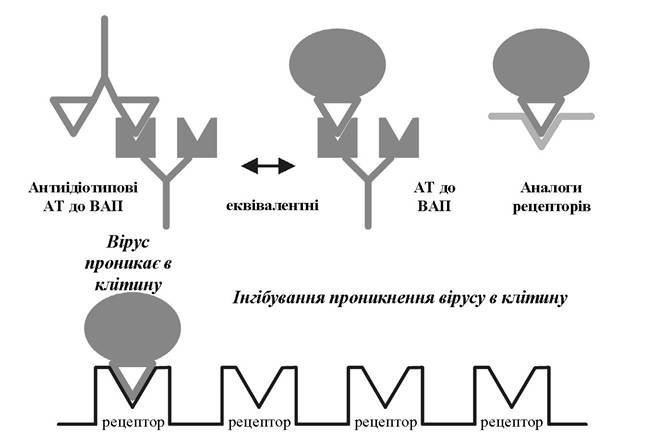
Рис. 5.10. Способи інгібування взаємодії вірусу з клітиною за допомогою агентів, які впливають на вірусасоційовані протеїни (АТ - антитіла; ВАП - вірусасоційовані протеїни)
Розроблено два типи інгібіторів проникнення ВІЛ у клітину. Так, препарат енфувіртид (Fuzeon) взаємодіє з поверхневим вірусним білком gp41 і перешкоджає його конформаційним змінам, необхідним для ініціації злиття вірусної та клітинної мембран, а інший препарат, маравірок, приєднується до хемокі- нового рецептора CCR5, блокуючи його від взаємодії з вірусним глікопротеїном gp41.
Іншим прикладом інгібіторів злиття мембран є препарат палівізумаб (СИНАГІС), до складу якого входять гуманізовані моноклональні антитіла IgG1is, які взаємодіють з епітопом А антигену F-білка респіраторного синцитіального вірусу. Молекула палівізумабу складається з людських (95 %) і мишачих (5 %) послідовностей.
Інгібітори стадії депротеїнізації. Існує обмежена кількість препаратів, що впливають на стадію депротеїнізації вірусу. Це пов'язано з двома причинами: по-перше, ця стадії порівняно погано досліджена, по-друге, депротеїнізація вірусів зазвичай здійснюється клітинними ферментами. Найвідомішими препаратами з цієї серії є похідні адамантану - амантадин і ремантадин (активні проти вірусу грипу А).
Мішенню дії амантадину є білок М2, який здійснює транспорт протонів усередину вірусу. За рахунок цього відбувається дисоціація вірусного рибонуклеопротеїнового комплексу. Крім того, білок М2 відіграє певну роль при транспортуванні пост- трансляційних вірусних глікопротеїнів на клітинну поверхню. На рис. 5.11 зображено можливий механізм взаємодії амантадину з М2-білком вірусу грипу типу А.
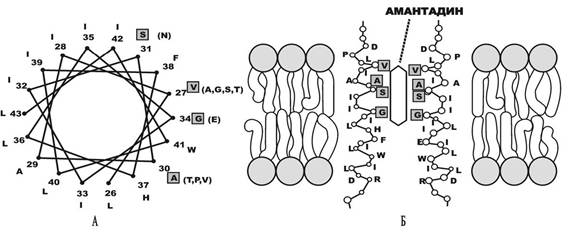
Рис.
5.11. Взаємодія амантадину з М2-протеїном вірусу грипу АБілок М2 є тетрамером з порою всередині чотирьох субодиниць. На рисунку він схематично зображений у вигляді колеса зі спіральною симетрією (А). Амінокислоти (Asp24-Arg45) позначені однією літерою. На чотирьох позначених локусах у дужках наведено амінокислоти, які найчастіше виявляються та можуть замінювати нормальні, що приводить до подальшої селекції амантадинрезистетних штамів. На (Б) зображено схему взаємодії амантадину з двома діагонально розташованими d-спіралями білка М2 в біліпідному шарі. Полярний бік спіралей формує внутрішню поверхню іонного каналу.
Інгібітори синтезу вірусної нуклеїнової кислоти та процесингу. Вірусні ферменти, які відповідають за реплікацію вірусу, є привабливими мішенями для дії антивірусних препаратів. Одними з найефективніших інгібіторів вірусних поліме- раз є аномальні нуклеозиди. Зазвичай ці препарати потрапляють у клітину у вигляді пропрепаратів, які, перш ніж стати активними, мають фосфорилуватися вірусним ферментом. Саме в цьому полягає селективна дія аномальних нуклеозидів.
Прототипом аномальних нуклеозидів є препарат ацикло- вір - ациклічний аналог нуклеозиду. Його дія заснована на фосфорилуванні тимідинкіназою вірусу герпесу. У зв'язку з тим, що ферменти клітини-хазяїна не фосфорилюють ацик- ловір з достатньою ефективністю, ацикловірмонофосфат утворюється переважно в клітинах, заражених вірусом герпесу, у яких присутній необхідний вірусний фермент. Ацикловірмонофосфат перетворюється клітинними ферментами на трифосфат, який інтерферує із синтезом вірусної ДНК, пригнічуючи вірусну ДНК-полімеразу та діючи як термінатор синтезу ДНК у випадку включення в зростаючий ланцюг вірусної ДНК (рис. 5.12).
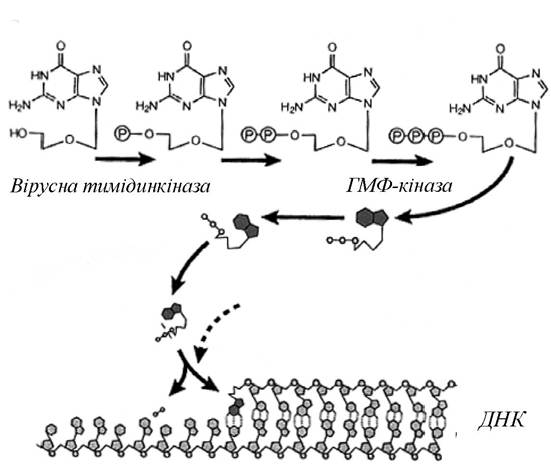
Рис. 5.12. Механізм дії ацикловіру
Відомо три класи антиретровірусних препаратів, які інгібують активність зворотної транскриптази: нуклеозидні інгібітори зворотної транскриптази (азидотимідин, ламівудин, абакавір), нуклеотидні інгібітори зворотної транскриптази (тенофовір), ненуклеозидні інгібітори зворотної транскриптази (невірапін, делавірдин, ефавіренз).
Дія препаратів перших двох класів подібна до дії ацикловіру. Азидотимідин фосфорилюється клітинною тимідинкіназою в інфікованих та неінфікованих клітинах і метаболізує послідовно до моно-, ди- та трифосфату. Вбудовуючись у ланцюг ДНК, що формується, трифосфат азидотиміди- ну стає ефективним інгібітором зворотної транскриптази вірусу. З'-азидогрупа запобігає формуванню 5'- та З'-фосфодиефірних зв'язків і блокує синтез кінцевих ланцюгів вірусної ДНК. Ці препарати неконкурентно зв'язуються з ферментом безпосередньо близько до ділянки приєднання нуклеотидів. У результаті фермент взаємодіє з меншою кількістю нуклеотидів, і процес полімеризації ДНК суттєво гальмується.РНК -залежні РНК-полімерази РНК-вмісних вірусів є унікальними ферментами, оскільки тваринні клітини їх не мають. Ці полімерази виконують різні вірусоспецифічні функції, включаючи синтез праймерів і сар-структур, розпізнавання вторинної структури вірусного геному. Крім того, перспективними мішенями для антивірусної дії є хелікази, що кодуються багатьма РНК-вірусами. Нині ведеться інтенсивний пошук інгібіторів унікальних вірусних хеліказ і РНК-залежних РНК-полімераз, але активні сполуки поки що не знайдені.
Інгібітори протеаз. Інфекційна активність вірусу грипу пов'язана з протеолітичним розщепленням гемаглютиніну на легкий і важкий ланцюги аргінін-специфічними протеазами невірусного походження. З погляду на це як можливий антивірусний препарат досліджувався камостат - інгібітор серинових протеаз. Камос- тат показав антигрипозну активність як in vitro, так і in vivo. Крім того, в умовах in vitro камостат пригнічував розмноження амантадинрезистентних штамів вірусів грипу А та В. Багато вірусів потребують специфічних протеаз для розщеплення полі- протеїнів на функціональні субодиниці під час їх самозбірки. Усі герпесвіруси кодують серинову протеазу, яка є необхідною для формування віріона. Перспективність цього ферменту як мішені для антивірусної дії полягає в незвичній третинній структурі. Крім того, усі герпесвіруси кодують аналоги даної протеази.
Детальне знання структури й механізму дії таких протеаз відкриває нові горизонти для пошуку антигерпетичних препаратів.Вірус гепатиту С кодує як найменше дві протеази, які відповідають за процесинг вірусних протеїнів. NSS-протеїн є привабливою антивірусною мішенню, оскільки він необхідний для реплікації вірусу. Хоч даний вірус не розмножується в культурі клітин, що значно ускладнює процес пошуку антивірусних агентів, досліди зі спорідненими флавівірусами свідчать про антиві- русну ефективність інгібіторів NSS-протеази.
Інгібітори регуляторних протеїнів. Багато вірусів контролюють свій реплікативний цикл за допомогою унікальних регуляторних протеїнів. Найчастіше ці білки виконують свою функцію, зв'язуючись зі специфічними послідовностями нуклеїнової кислоти або інших білків вірусу. Регуляторні протеїни виконують важливу роль для реплікації вірусу й тому є ідеальними мішенями для пошуку антивірусних препаратів.
Інгібітори асамблювання та виходу з клітини. Відомо небагато препаратів, що пригнічують останні стадії реплікації вірусу. Серед них метисазон - інгібітор поксвірусів та інгібітори нейрамінідази вірусу грипу оселтамівір і замамівір. Цінність даних препаратів полягає в тому, що, оскільки вони пригнічують останні стадії життєвого циклу вірусу, то проблеми, пов'язані із селекцією резистентних штамів, зводяться до мінімуму.
5.4.
Еще по теме Мішені дії антивірусних агентів:
- Резистентність до антивірусних препаратів
- Нові напрями у створенні антивірусних препаратів
- Класифікація антивірусних хіміопрепаратів
- Молекулярні основи дії ІНФ на клітину
- Дії у випадку загрози виникнення радіаційної небезпеки:
- Дії у випадку раптового виникнення радіаційної небезпеки:
- ЗАПИТАННЯ ДЛЯ САМОКОНТРОЛЮ
- Тетрацикліни
- Одним з важливих теоретичних питань розвитку епідеміології як науки є проблеми інтеграції та диференціації.
- Імуностимулятори та імуномодулятори
- Хіміотерапія вірусних інфекцій
- Фактори неспецифічної резистентності у хворих та стан імунної системи при пневмонії в різні періоди розвитку захворювання в залежності від величини запального процесу в легенях