РОЛЬ КОЖИ И ЛЕГКИХ В АДАПТАЦИИ К ПУСТЫНЕ
Весьма важное место в ограничении потерь воды у пустынных организмов занимают адаптивные особенности отдачи воды кожей и легкими при дыхании. Эти потери составляют значительную часть так называемых незаметных потерь веса (perspiratio insensibilis), которые издавна являются одним из путей изучения общего обмена веществ.
Кожно-легочные потери складываются из потерь воды при дыхании путем почти полного насыщения водяными парами вдыхаемого воздуха, испарения воды с поверхности (потоотделение и кожная перспирация) и, наконец, из разности веса поглощаемого кислорода и выделяемой при кожном и легочном дыхании углекислоты. Эта разность учитывается при определении кожно-легочных потерь воды путем одновременного определения газообмена и незаметных потерь веса.Кожно-легочный механизм экономии воды у пустынных организмов был впервые обнаружен А. Й. Щегловой (1949). Кожно-легочные потери воды у большой песчанки значительно ограничены по сравнению с лабораторной белой крысой. Важно и то, что при повышении температуры среды у белой крысы кожно-легочные потерн воды резко возрастают, а у пустынного вида — большой песчанки это возрастание незначительно. В итоге при высоких температурах среды (30°С) кожно-легочные потери воды у белой крысы почти в 2 раза превышают таковые у большой песчанки (рис. 70). Песчанка лишь в минимальных количествах расходует воду на терморегуляцию. А так как химическая терморегуляция у большой песчанки также выражена слабо, то, очевидно, существование этого вида в пустыне обусловлено своеобразной формой поведения — роющей деятельностью, обеспечивающей в глубоких норах такой термический режим, который не нарушает постоянства температуры тела (Щеглова, 1949).
Ограничение водного обмена было затем подтверждено на желтом суслике, тушканчике Северцева (Филатова, 1944). У этих видов оказатся более низкий водный
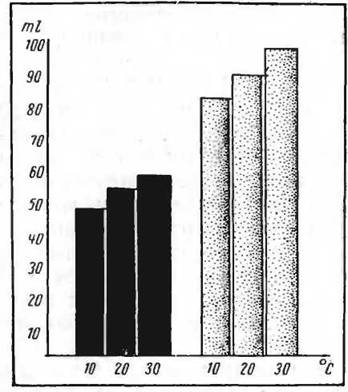
Рис.
70. Кожно-легочные потерн воды у большой песчанки и белой крысы при температурах среды 10°, 20°, 30° С. Черные столбики — большая песчанкаобмен и более низкие экстраренальные потери воды, нежели у гребенщиковой песчанки — обитателя орошаемых земель и белой крысы. В дальнейшем установлено (Щеглова, 1949, 1952а, 19526, 1959), что ограничение водного обмена можно наблюдать у многих пустынных грызунов — тонкопалого суслика, малоазиатской песчанки, малого суслика. При сравнении трех видов сусликов — тонкопалого, желтого и малого, самый низкий уровень водного обмена оказался у тонкопалого суслика, несколько больший у желтого и самый высокий — у малого (Щеглова, 19526). Различия могут быть объяснены не только особенностями расселения этих видов, но и сроками залегания в спячку желтого и малого сусликов. Первый залегает в спячку очень рано, когда содержание воды в растениях пустыни снижается. Малый суслик залегает в спячку поздно и долго питается содержащей воду степной растительностью.
Подобные же отношения у пустынных видов грызунов Нового Света (Schmidt-Nielsen a. Schmidt-Nielsen, 1950, 1952). Потери воды кожей и легкими у пустынных видов составляют около 2/3 потерь воды у лабораторных животных. Обращают на себя внимание низкие величины потерь воды у диких видов грызунов, особенно домовой мыши, которая может находиться без воды неопределенно долгое время (Bing a. Mendel, 1931). Этот факт интересен как пример приспособления к недостатку воды в условиях, весьма далеких от пустынных.
Ограничение кожно-легочных (экстраренальных) потерь воды у обитателей пустыни определяется особенностями кожного покрова и испарения воды легкими. Так как грызуны не обладают потовыми железами, то роль кожи в кожно-легочных потерях воды невелика. Выдыхаемый воздух у непустынных млекопитающих насыщен водяными парами практически на 100%- Небольшое недонасыщение может быть лишь при очень поверхностном дыхании.
Дефицит насыщения выдыхаемого воздуха был впервые обнаружен у верблюда в Южном Казахстане (Бах- тиозина, 1948), а затем тщательно изучен как в условиях нормального снабжения водой, так и при водном голодании (Бахтиозина, 1960а, 19606).
Обнаружилось, что верблюды при достаточном снабжении водой в жаркие летние месяцы способны выдыхать воздух с относительной влажностью, не превышающей 69—86% при исходной величине 88—95%. После прекращения водного голодания относительная влажность выдыхаемого воздуха возвращалась к исходной на 5—6-й день. Относительная влажность вдыхаемого воздуха была в этих опытах от 26,8 до 55% (табл. 28). Недонасыщение выдыхаемого воздуха водяными парами при водном голодании было ярко показано на курдючных овцах (в Казахстане в летний период). При низкой относительной влажности, не превышающей 45—47%, и температуре около 31—33°С относительная влажность выдыхаемого воздуха до водного голодания превышала 90 %• При 4-дневном водном голодании относительная влажность выдыхаемого воздуха снизилась с 78 до 62 %.При редком водопое (1 раз в три дня) у каракульских овец наблюдается снижение потерь воды легкими (Падучева, Ковальский и Шраер, 1957). По сравнению с ежедневным водопоем снижение потерь воды через легкие может достигать в апреле 1,4%, а в июле 23,2%. Авторы объясняют это уменьшением легочной вентиляции и падением окислительных процессов.
Было высказано предположение, что длинные носовые ходы некоторых пустынных животных, смачиваемые выделяемой слизистой жидкостью, могут благодаря ее испарению охлаждать область морды и таким образом конденсировать водяные пары. Однако это соображение лишено какой-либо энергетической основы, так как количество испаренной с поверхности жидкости должно быть равно количеству удержанной, т. е. не вызывать
Таблица 28
Относительная влажность вдыхаемого и выдыхаемого верблюдом воздуха при водном голодании (Бахтиазина, 1960а)
| День опыта | Температур і окружаю ш го воздуха (°С) | Относительная влажность воздуха | |
| вдыхаемого (%) | выхыхаемог» (%) | ||
| 1 | Нормаль 25,7 | ные условия 55,0 | 95,0 |
| 2 | 25,6 | 38,0 | 90,0 |
| 3 | 24,2 | 54,2 | 93,0 |
| 4 | 22,4 | 52,4 | 91,8 |
| 5 | 24,8 | 42,6 | 88,8 |
| 1 | Период водь 28,3 | юго голодания 31,2 | 82,2 |
| 2 | 29,9 | 32,4 | 79,8 |
| 3 | 27,6 | 36,8 | 86,2 |
| 4 | 26,6 | 39,5 | 74,6 |
| 5 | 24,7 | 38,1 | 79,8 |
| 6 | 26,0 | 40,8 | 72,7 |
| 7 | 25,4 | 34,0 | 78,3 |
| 8 | 25,7 | 35,0 | 73.0 |
| 9 | 25,7 | 36,5 | 69,5 |
| 10 | 23,3 | 41,6 | 70,4 |
| 11 | 25,6 | 26,8 | 69,3 |
| 1 | Восстановит 26,0 | пельный период 37,7 | 73,0 |
| 2 | 26,2 | 35,0 | 83,8 |
| 3 | 24,1 | 37,7 | 79,8 |
| 4 | 23,8 | 40,1 | 82,4 |
| 5 | 24,9 | 29,5 | 71,3 |
| 6 | 25,7 | 32,7 | 91,6 |
| 7 | 25,0 | 32,0 | 91,0 |
какого-либо энергетического эффекта. Было высказано и предположение (Слоним, 1962), что здесь может иметь место активное удержание водяных паров слизью, скапливающейся по ходу верхних дыхательных путей.
Последнее требовало для подтверждения изучения ферментативных механизмов, изменяющих физико-химические свойства муцинов. Были необходимы и специальные патофизиологические исследования легочной ткани и верхних дыхательных путей, в том числе и межклеточных образований, играющих (Гинецииский, 1962) важную роль в перемещениях воды в организме.Специальные исследования активности тканевой гиа- луронидазы в легочной ткани и ткани почек (Григоренко, 1967, 1968), показали, что у впадающего в зимнюю спячку краснощекого суслика (Citellus erythrogenys) и у большой песчанки наблюдается взаимно противоположно направленная динамика активности этого фермента. У песчанки наибольшая активность в засушливый период года, у суслика — в период впадения в зимнюю спячку и сразу после пробуждения. В почках наблюдаются те же взаимоотношения. Эти фактические данные позволяют думать, что проницаемость межклеточных образований для воды и состояние кислых мукополисаха- ридов могут играть существенную роль в динамике удержания воды в легких. Введение антидиуретического гормона вызывает однозначное изменение активности фермента в легких и почках.
С другой стороны, гистохимические, ультрамикроско- пические и гистологические методы исследования (Соболева, 1967, 1968) осветили и морфологическую картину в легких пустынных животных. Оказалось, что у верблюда и большой песчанки строение легких отличается значительной длиной воздухоносных путей. Эпителий, выстилающий бронхиоли, цилиндрический, лишь в самых конечных отделах переходит в плоский — альвеолярный. Такое строение легочной ткани обеспечивает меньшие потери воды при поддержании высокого напряжения водяных паров в альвеолах.
Особенную роль приходится отвести большим клеткам альвеолярного эпителия у всех млекопитающих, принимающим на себя функцию фагоцитов-макрофагов. Превращение этих клеток в макрофаги сопровождается изменениями, регистрируемыми гистохимически: исчезает активность неспецифической эстеразы и проявляется накопление липазы.
Такое явление сопровождает дедифференцировку клеток и превращение их в макрофаги.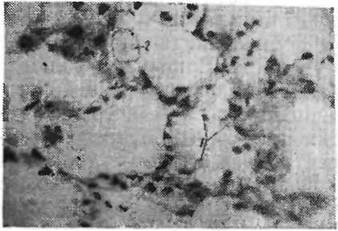
Рис. 71. Микрофотография легкого большой песчанки после дегидратации.
1 — клетки на разных стадиях дедифферен- цировки. 2 — в просвете альвеолы пнно- цит (по Соболевой, 1964)
После содержания больших песчанок на безводной диете большие клетки альвеолярного эпителия берут на себя новую функцию — пнноцитов, т. е. клеток, задерживающих воду. При этом клетки оказываются бедными липидами, белком и мукополисахаридамн. Цитоплазма содержит большое количество неокрашивающихся вакуолей. Клетки эти часто наблюдаются не только в стенке, но и в просвете альвеол. У белых крыс при содержании на безводной диете на 6—7-й день также наступают начальные стадии дедифференцировки альвеолярного эпителия, но до образования пиноцитов дело, по-видимому, не доходит (рис. 71). У верблюда после 20 дней лишения воды также появляются большие комплексы ли- пофагов, в которых можно предполагать начальные стадии развития пиноцитов.
Кроме того, в этих же исследованиях были обнаружены значительные изменения в жировой ткани, расположенной подплеврально, заключающиеся в нарушении структуры жировой ткани и накоплении кислых мукополисахаридов. При дальнейшем водном голодании жир почти исчезает и заменяется новой тканью, состоящей из неправильных многоугольных и округлых клеток с уплощенными ядрами, передвигающимися к периферии. В цитоплазме этих клеток мало белка, нет липидов и мукополисахаридов и содержится большое количество вакуолей. Это позволяет думать о наличии системы пиноцитов, расположенных в подплевральной области и удерживающих воду.
Изложенные данные позволяют выявить пути удержания воды легкими при дыхании и понять интимные механизмы этого явления. Окончательное решение этих вопросов требует еще дальнейших исследований.
Еще по теме РОЛЬ КОЖИ И ЛЕГКИХ В АДАПТАЦИИ К ПУСТЫНЕ:
- Роль сердечно-сосудистой системы в процессах адаптации к гипоксии
- 18. Профессиональная адаптация – роль психолога в этом процессе.
- 1.2.3. Роль функциональной асимметрии полушарий в процессах адаптации
- 1.2.3. Роль функциональной асимметрии полушарий в процессах адаптации
- 1.2.2. Роль эмоций в процессе адаптации к умственной деятельности
- 1.2.2. Роль эмоций в процессе адаптации к умственной деятельности
- 17. Четыре онтогенетические жизненные позиции человека(по Э.Берну, Д.Харрису) их влияние на психологическое здоровье и социальную адаптацию, роль семьи
- Жаркий климат пустынь
- РОЛЬ ХИРУРГИИ В ЛЕЧЕНИИ БОЛЬНЫХ С МЕТАСТАЗАМИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ В ЛЕГКИХ
- Тема 6. Дифференциальная диагностика очаговых заболеваний легких. Инфильтративный туберкулез легких, пневмония, рак легких
- Особенности пищеварения и водного обмена пустынных млекопитающих
- Высокогорье и горно-пустынная местность
- Тема занятия. ОБСТРУКТИВНЫЕ И РЕСТРИКТИВНЫЕ ЗАБОЛЕВАНИЯ ЛЕГКИХ. РАК ЛЕГКИХ
- Тема 7. Дифференциальная диагностика очаговых заболеваний легких. ТЭЛА. Эозинофильное поражение легких