Мембранные каналы и переносчики как ферменты
До сих пор мы рассматривали работу мембранных транспортных белков на микроскопическом молекулярном уровне. Оказывается, что применение прямо противоположного, феноменологического, способа описания транспортных мембранных процессов позволяет экспериментально определять многие параметры транспортных белков, даже не проводя микроскопические исследования методами молекулярной биологии.
Кинетическую теорию переходного состояния Эйринга и Поляни, используемую в ферментативном катализе, успешно применяют для описания различных транспортных систем, в том числе для количественного описания работы молекул-переносчиков и ионных каналов.
В основе этого подхода лежит предположение о том, что система может находиться в нескольких дискретных состояниях, каждому из которых соответствует стандартное значение электрохимического потенциала. При этом взаимные переходы между двумя состояниями происходят с преодолением энергетических барьеров и константы скоростей переходов зависят от высоты этих барьеров.
На рисунке 99 представлена схема ионного канала и соответствующие профили свободной энергии.
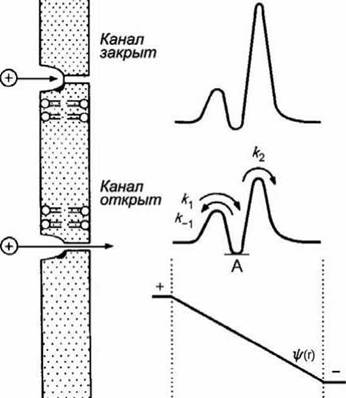
Рисунок 99 - Изменение профиля свободной энергии иона в ионном канале при деполяризации мембраны
Эти профили соответствуют случаю, когда в канале имеется единственное место связывания, локализованное где-то вблизи от входа в канал. Глубина впадины (точка А) отвечает слабому связыванию.
При наличии трансмембранной разности потенциалов на профиль свободной энергии накладывается профиль электрического потенциала щ(г). В этом случае высота энергетических барьеров, соответствующих к, и к2, будет меньше, и скорость транспорта катионов через канал увеличится. Минимумы на кривых изменения свободной энергии соответствуют местам связывания транспортируемых веществ.
Можно предположить, что канал или переносчик имеет одно или несколько мест связывания переносимых веществ.При достаточно высоких концентрациях переносимого вещества все эти места оказываются занятыми, и скорость переноса достигает своего максимального значения wmax, равного максимальной скорости работы фермента.
Примем для простоты, что ионы изначально присутствуют только с одной стороны мембраны, внутри канала имеется единственное место связывания, и что ионы не могут свободно проникать внутрь канала или покидать его.
Случай, когда внутри канала имеется много мест связывания, по которым ион может последовательно передаваться, формально сводится к ситуации, когда в канале есть единственное место связывания, и кинетика работы такого фермента сводятся к стационарной кинетике Михаэлиса- Ментен [9].
Если принять, что мембранный потенциал срм равен нулю, то такую "ферментативную реакцию" можно представить следующим образом 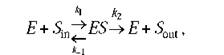
где £ - канальный белок (фермент) в "открытом" состоянии; 51п и Sout - транспортируемое вещество (субстрат) внутри и снаружи мембраны; ES - комплекс субстрата с местом связывания внутри канала.
На рисунке 99 показаны профили свободной энергии для закрытого и открытого каналов.
Еще по теме Мембранные каналы и переносчики как ферменты:
- Мембранные переносчики.
- Бло:каторы Na + - ионных каналов возбудимых мембран
- Блохи как переносчики чумы.
- Белок пресенилин-1 работает как канал «утечки» кальция из эндоплазматического ретикулума
- Роль функциональной активности кальций-зависимых калиевых каналов большой проводимости и фоновых 2Р калиевых каналов в нейропротектирующем действии ИЛ-10
- Потенциалочувствительные натриевые каналы
- Потенциалочувствительные калиевые каналы
- Оценка передвижений и контактов носителей и переносчиков возбудителя чумы.
- А. Специфическая передача личинок паразитических червей насекомы ми-переносчикам и [30] [31]
- Передача паразитических червей через посредство насекомых-переносчиков.
- Нерегулируемые калиевые каналы
- 9.0.5. Биологическая регуляция ферментов
- 3.2. Способы оценки численности носителей и переносчиков возбудителя чумы
- Ферменты пищеварительного тракта
- Ферменты пищеварительного тракта
- Амилоид-деградирующие ферменты
- 9.0. Ферменты
- 9.0.6. Ферменты и лекарственные вещества