Потенциалочувствительные натриевые каналы
Потенциалочувствительные натриевые каналы закрыты, когда мембрана находится в состоянии покоя, мембранный потенциал в таком состоянии называется потенциалом покоя.
Небольшая начальная точечная деполяризация мембраны стимулирует такое конформационное изменение в близлежащих канальных белках, при котором натриевые каналы открываются, и по этим открытым каналам ионы Na+ перемещаются внутрь клетки, тем самым ещё больше увеличивая деполяризацию (положительно заряжая цитозольную сторону мембраны).
Чем больше становится деполяризация, тем больше открывается натриевых каналов, и тем больше ионов Na+ входит в клетку. По мере того как всё большее число ионов Na+ переходит в цитозоль зона избыточного положительного заряда на цитозольной стороне мембраны, и соответствующая зона отрицательного заряда на экзоплазматической стороне мембраны (зона деполяризации) распространяется от места первоначальной деполяризации.
Такое распространение зоны деполяризации захватывает всё большую площадь мембраны, в пределах которой натриевые каналы открываются, и поток ионов Na+ внутрь клетки всё возрастает.
Процесс развивается лавинообразно, и за долю миллисекунды открывается настолько много натриевых каналов, что суммарный поток ионов Na+ внутрь клетки становится намного большим, чем встречный поток ионов К+ наружу через калиевые каналы.
Когда мембранный потенциал достигает равновесного значения для ионов натрия, то движение ионов Na+ внутрь клетки под действием градиента концентрации уравновешивается движением этих ионов наружу под действием избыточного положительного потенциала тех ионов Na+, которые уже успели войти в цитозоль, и дальнейший рост амплитуды мембранного потенциала останавливается. Значение мембранного потенциала в таком состоянии называется потенциалом реверсии.
На рисунке 128 показаны основные структурные конформации натриевого канала.
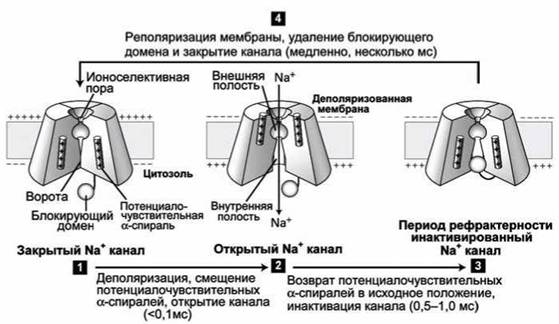
Рисунок 128 - Цикл работы потенциалочувствительного натриевого канала
В состоянии покоя "ворота" канала закрыты (рисунок 128(1)). Деполяризация мембраны приводит к смещению потенциалочувствительных а-спиралей (2) (на рисунке они смещаются вверх) к экзоплазматической стороне мембраны, что вызывает конформационное изменение структуры белка, и ворота открываются, позволяя ионам перемещаться внутрь клетки. Через 1 мс канал механически блокируется цитозольным доменом, который как пробка просто "затыкает" канал (3). Пока мембрана деполяризована - канал остаётся заблокированным. Через несколько миллисекунд после реполяризации и возвращения мембранного потенциала к значению потенциала покоя сместившиеся вниз (в исходное состояние) заряженные а-спирали "выдавливают пробку" блокирующего домена из канала, и канал возвращается в исходное состояние (рисунок 128(4)).
13.4.
Еще по теме Потенциалочувствительные натриевые каналы:
- Натриевые каналы
- Строение потенциалочувствительных ионных каналов
- Потенциалочувствительные калиевые каналы
- Роль функциональной активности кальций-зависимых калиевых каналов большой проводимости и фоновых 2Р калиевых каналов в нейропротектирующем действии ИЛ-10
- Нерегулируемые калиевые каналы
- Фильтры селективности ионных каналов.
- 3.1.2. Канал GPRS
- Рецепторы - ионные каналы.
- Временное разделение каналов (ВРК)
- Основные понятия и характеристики каналов связи
- 2.4.1. Частотное разделение каналов (ЧРК)
- Каналы связи и порты ввода-вывода
- Блокаторы кальциевых каналов.
- Разделение каналов по форме (РКФ)
- Глава 11 Блокаторы кальциевых каналов
- 3. Использование каналов распространения товара
- Белок пресенилин-1 работает как канал «утечки» кальция из эндоплазматического ретикулума
- Ионные каналы в модели Михаэлиса-Ментен.