«Сборка» многоклеточных организмов.
Архейская жизнь предполагала сложную систему отношений между одноклеточными организмами. Они «общались» между собой с помощью сигнальных молекул, известных нам сегодня под общими названиями хемокинов и лимфокинов.
Их рецепторы воспринимали химические сигналы, поступившие из окружающей среды, а белки трансдукции сигналов управляли клеточными функциями. Ни ЦНС, ни «мыслительной деятельности» не требовалось для существования таких организмов в течение сотен миллионов лет. Но такого количества жизни, которое существовало в конце архея и в протерозое, на планете никогда больше не было, чему есть следующее подтверждение.В 1990-е гг. на архипелаге Шпицберген открыта уникальная по полноте последовательность позднепротерозойских осадков, отлагавшихся в период с 850 до 600 млн лет назад. Исследовав эти осадки на предмет изотопного отношения 12С/13С, Э. Нолль (1996) установил, что на протяжении всего этого времени темпы захоронения органического углерода оставались самыми высокими за всю историю Земли (цит. по работе Еськова К. Ю., 2004), Несомненно то, что жизнь именно тогда пережила свой период количественного «расцвета».
И такая жизнь могла бы быть вполне самодостаточной, если бы в ее организацию не вмешивалась ретровирусная эволюция (см. раэд. 1.1). Ан- тиэнтропийные процессы наращивания и усложнения генома одноклеточных организмов, вызванные увеличением числа копий их генов и появлением новых экзонов, продолжались (их механизмы см. в разд. 1.2) — эволюционный маховик раскручивался; 840-740 млн лет назад появились первые эволюционные пробы многоклеточности. Ими были бесскелетные организмы, названные по месту обнаружения хайнаньской биотой — первый акт творения многоклеточной жизни. Для них характерно членистое строение, отражающее дупликацию генетического материала, осуществляемую ретроэлементами.
Однако сложные многоклеточные формы жизни, как и миллиарды лет до этого (в нашем ощущении времени), все еще не закреплялись естественным отбором.
Демону Дарвина не хватало механизма поддержания «многоклеточности», предполагающего соединение в единое целое «своих» и исключение «не своих» клеток. Этот механизм мог сформироваться на основе молекул специфической адгезии. Но для того чтобы такая «специфичность» появилась у поверхностных белков сотен разных клеток, формирующих целостный организм, естественный отбор должен был «перепробовать» миллионы различных вариантов адгезионных молекул. Выбирать же Демону Дарвина было пока не из чего, о чем свидетельствуют окаменелые останки так называемой эдиа каре кой фауны, сменившие хайнаньскую

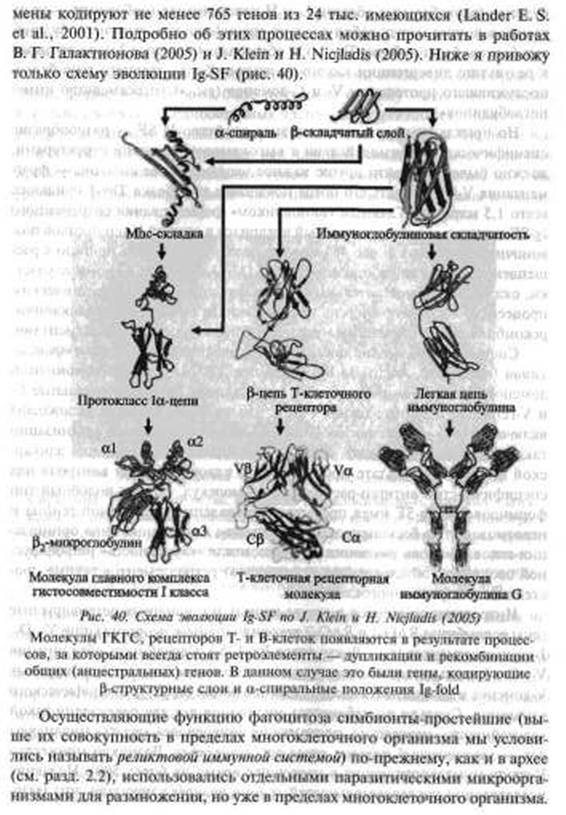

и доступ к рецепторам клеток, в которых происходит их персистирование и размножение, т. е. в фагоцитирующих клетках, теперь уже специализирующихся в роли «мусорщиков». Что и произошло в отношении других паразитов свободноживущих простейших, например, тех, которых мы знаем как возбудителей чумы, сибирской язвы, туляремии, ортопоксви- русных инфекций и др., не способных интегрироваться с геномом хозяина. Случайно попав в организм многоклеточного животного, они либо вызывают его гибель, либо элиминируются клетками иммунной системы, осуществляющими функцию «присмотра» за их потенциальными хозяевами — фагоцитирующими клетками (см. ниже «Иммунная система»). Использовать его для собственного поддержания в природе они не могут.
Во-вторых, «унаследованные» многоклеточными организмами ретро- вирусы и другие ретроэлементы без механизма их обмена с другими многоклеточными организмами, подверглись бы нейтральной эволюции и генетико-автоматическим процессам.
Оба этих явления, разумеется, имели место в ходе эволюции многоклеточных форм жизни постоянно, но они компенсировались горизонтальной передачей экзогенных ретровирусов и их последующей (а иногда повторной) эндогенизацией (см.
разд. 1.2). А вот ответ на вопрос — почему естественный отбор закрепил горизонтальную передачу ретровирусов за многоклеточными, может быть следующим. Он не мог отобрать другие формы многоклеточной жизни — без ретровирусов они не «собрались» в единое целое и не размножались.Эндогенные ретровирусы активны на всех этапах репродукции многоклеточных организмов: сперматогенез и овуляция, регуляция экспрессии генов плаценты, трофоэктодермальный рост и дифференциация (trophecto- derm growth and differentiation), взаимодействие между плодом и матерью, подавление иммунных реакций со стороны организма материи в отношении плода, дифференциация тканей плода и др. (см. работы Arimi М. М. et. аі., 2006; Prudhomme S. et al., 2005). Уже у кольчатых червей (Annelida, сегментированные черви) многочисленные фагоцитирующие клетки не способны поглощать живые сперматозоиды своего вида, но активно устраняют погибшие клетки и сперматозоиды других видов (Купер Э., 1980).
Показателен и сам механизм участия эндогенных ретровирусов в формировании плода. В плаценте оболочечные белки ретровирусов выполняют ту же роль белков слияния, что и при формировании протоклеточных образований в архее, и при переходе одноклеточных форм жизни в многоклеточные во времена так называемой эдиакарской фауны (см. рис. 39). Они экспрессируются в синцитиотрофобластном слое (syncytiotrophoblast layer), образованном посредством слияния мононуклеарных цитотрофо- бластов, и образуют участки синцития в тех участках плаценты, где начинается взаимодействие матери и плода (Dunlap К. et аі., 2006). Уже эти данные свидетельствуют о том, что информация, определяющая целостное развитие многоклеточного.организма, хотя и содержится в зиготе, но только как некая потенция, которая не реализуется без участия ретровирусов матери.
Путь экзогенным ретровирусам в многоклеточный организм прокладывается и подстраховывается самим многоклеточным организмом посредством полового процесса. Феномен хорошо изучен у людей. Семенная жидкость содержит большое количество различных иммуносупрессивных агентов.
Они подавляют иммунные реакции на аллогенные белки, представленные на поверхности сперматозоидов и в семенной жидкости, со стороны слизистых поверхностей нижних половых путей женщины. Это увеличивает продолжительность жизни сперматозоидов и гарантирует зачатие (Kelly R.W. et аі., 1991).Но одновременно семенная жидкость препятствует устранению инфицированных вирусами макрофагов посредством киллерной активности Т-клеток и натуральных киллерных клеток, так как действие цитокинов, вызывающих такие реакции, переключается микроорганизмом «с точностью наоборот». Активность этих клеток стимулируется IL-12 и подавляется IL-10, однако в нижних родовых путях женщины активность IL-12 подавлена, a IL-10 стимулирована (Kelly R. et аі., 1997).
Сами сперматозоиды к передаче экзогенных ретровирусов между особями вида Homo sapiens отношения не имеют. По данным A. J. Quayle et аі. (1997), в 75 % исследованных им образцов спермы ВИЧ-серопозитив- HbLx людей ВИЧ были инфицированы Т-клетки, и в 38 % образцов — макрофаги. Но ими ни в одном эксперименте не была обнаружена ДНК ВИЧ в подвижных сперматозоидах. По данным же Р. L. Vemazza et аі. (1997), количество РНК ВИЧ крови коррелирует с количеством РНК ВИЧ семенной жидкости, т. е. по отношению к ВИЧ кровь и семенная жидкость являются «сообщающимися сосудами».
Система иммунитета, которую привычно называют «врожденной» (фагоцитоз, комплемент), также преследует интересы ВИЧ, а не человека. По данным Н. Bouchlal et аі. (2002), оба, Х5- и RS-тропные штаммы вируса проникают через слизистую и инфицируют эпителий половых путей благодаря активации факторов комплемента, содержащихся в семенной жидкости. Благодаря опсонизации они инфицируют клетки, имеющие рецептор комплемента на своей поверхности.
Еще более надежно «подстраховывается» иммунной системой человека передача ВИЧ от матери к плоду. Его транспорт через синцитиотрофобла- стный слой осуществляется с помощью антител к gpl20 и gp41 по обоим механизмам, объединенным под названием «феномен антителозависимого усиления инфекции» (см.
Toth F. D. et аі., 1994; и разд. 3.3, «Феномен антителозависимого усиления инфекции»).Третий акт творения — «кембрийский взрыв». Теперь, когда иммунная система первых позвоночных «научилась» поддерживать их целостную структуру и одновременно стала средой размножения ретровирусов, вращение маховика эволюции еще более ускорилось. Палеонтологи отмечают, что 490-435 млн лет назад (кембрий) на планете появились многоклеточные организмы современного типа, начался их выход из морей на материки и освоение ими всей поверхности Земли — этот феномен назван учеными кембрийским взрывом.
В триасе (триасовый период, 230-190 млн лет назад) вращение маховика эволюции достигло своего максимума. В эволюции таксонов дупликации отдельных генов уступают их кластерной организации, теперь уже кластеры генов дуплицируются ретроэлементами, что периодически проявляется «неадаптивными признаками» у стремительно появляющихся новых многоклеточных видов. По суше затоптались животные причудливых форм и огромных размеров, само существование которых трудно объяснить «дарвиновской приспособленностью»: диплодки — нелепые травоядные гиганты с маленькой головой на тонкой длинной шее из десятков повторяющихся позвонков, поворачивающихся только в горизонтальном направлении. При наличии длинной шеи тяжелому диплодку пришлось миллионы лет питаться низкорослой растительностью на болотах; стегозавры — массивные травоядные ящеры с двумя рядами асимметрично повторяющихся тонких треугольных костяных пластин на спине, предназначение которых до сих пор вызывает споры среди ученых.
В конце юра (юрский период, 190-136 млн лет назад) рыбу в морях ловили летающие ящеры-птеранодоны с размахом крыльев до 18 метров. И ответ на вопрос, кто тут к кому «приспосабливался» — вид к «неадаптивному признаку» или «неадаптивный признак» к виду, а вместе — к условиям окружающей среды, никак однозначно не следует из дарвиновских представлений об «эволюции без скачков». Но такое буйство разнообразия жизни должно было когда-то кончиться.
Оно и кончилось — маховик эволюции «пошел в разнос» в меловом периоде (136-66 млн лет назад), что проявилось, прежде всего, вымиранием рептилий. Им на смену в высвободившиеся экологические ниши пришли млекопитающие (палеогеновый период, 66-25 млн лет назад). Но их «золотой век» оказался еще короче, они стали угасать в неогене (неогеновый период, 25-1,8 млн лет назад). Ныне живущие слоны, носороги, лошадиные лишь жалкие остатки палеогеновой фауны млекопитающих.Торможение эволюции. Разумеется, некоторые палеонтологи объясняют массовые вымирания видов (кризисы биоразнообразия) тектоническими катастрофами и климатическими изменениями, и, в частности похолоданием климата. При этом они как бы не замечают того, что эти «вымирания» растягивались на десятки миллионов лет. Продолжительность же ледниковых эр на Земле составляет не менее трети всего времени ее эволюции за последние 2,5 млрд лет. А если учесть длительные начальные фазы зарождения оледенения и его постепенной деградации, то эпохи оледенения займут почти столько же времени, сколько и теплые, безледные условия. Интересно то, что великое оледенение пермского периода (закончилось 270 млн лет назад) не привело к принципиальному изменению фауны (более подробно о ледниковых периодах в истории Земли см. в кн. Джона Б. с соавт., 1982).
Сегодня (кайнозой) условия для существования жизни лучше, чем в конце архея и в протерозое, однако биологическое разнообразие на планете снижается. И тому должны быть более серьезные основания, чем деятельность человека, так как и ему самому она не дает идеальной адаптации к факторам внешней среды. За последние 30 тыс. лет из трех видов «мыслящих» приматов, вымерли два. Но и они уже не отражали того многообразия «мыслящей жизни», которое породили среди приматов толчки ретровирусной эволюции 5-2 млн лет назад (см, разд. 1.2 и рис. 6).
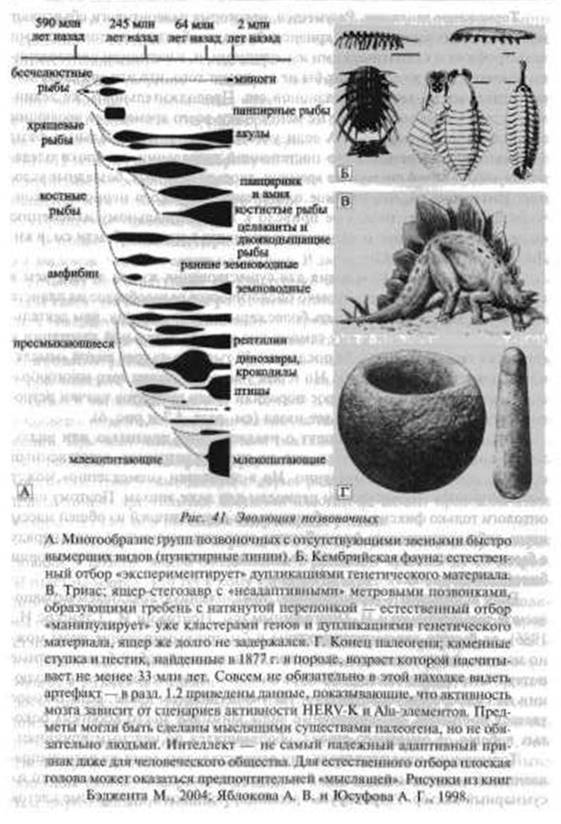
Когда эволюционисты пишут о неадаптивных признаках или видах, то обычно подразумевается то, что их должен устранить естественный отбор и, естественно, немедленно. Но в эволюции «немедленно» может соответствовать геологическим периодам или даже эпохам. Поэтому палеонтологи только фиксируют выборку видов-долгожителей из общей массы когда-либо возникавших форм. Быстро эволюционирующие виды наряду с быстро вымирающими видами не оставляют заметных следов в истории биосферы (рис. 41).
Если использовать терминологию, предложенную создателем неравновесной термодинамики И. Пригожиным (см. Пригожин И., Стенгерс И,, 1986), то быстро эволюционирующие и быстро вымирающие виды можно назвать диссипативными, т. е. они представляют собой безвозвратные потери энергии (диссипации), затраченной в ходе ретровирусной эволюции на самоорганизацию живого. На эволюционном древе, если таковое удается построить, диссипативные виды занимают место коротких боковых побегов от основного ствола, оборвавшихся по неясным причинам.
Торможение маховика эволюции осуществлялось теми же разнонаправленными механизмами, какими он раскручивался, поменялся только их суммарный вектор. Ретровирусы, реплицирующиеся в цитоплазме клеток

Псевдогены у человека н грызунов*
Таблица 1}
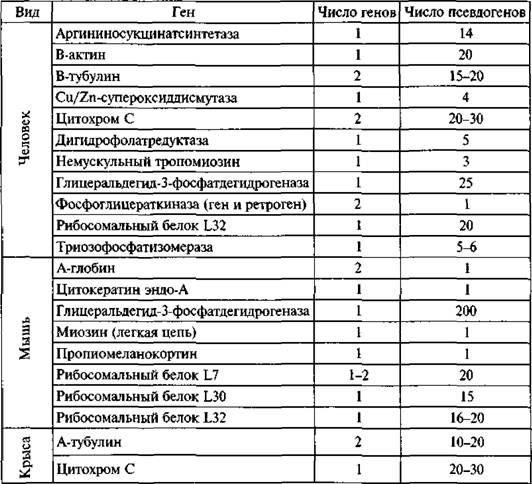
* C сайта: http://humbio.ru/humbio/molevol/(W0fda4a.htm.
свидетельствуют псевдоген и зация эволюционно наиболее активных их генов и возраст наиболее распространенных ретроэлементов (см. разд. 1.2); но по этим же причинам палеонтологи имеют возможность фиксировать вымирание отдельных видов как часть эволюционного процесса.
Еще по теме «Сборка» многоклеточных организмов.:
- Простейшие — симбионты многоклеточных
- №3 Организм человека как единая биологическая система. Уровни организации человеческого организма: молекулярный, клеточный, тканевой, органный, системный, организменный.
- Взаимоотношения опухоли и организма
- Реактивность организма
- Влияние опухоли на организм
- Глава 1. Необходимость очищения организма
- Уровни организации человеческого организма
- Роль организма в опухолевом процессе
- Очищение коллоидных растворов организма
- Элиминация веществ из организма
- Роль металлов в жизнедеятельности организмов
- 1. Представления о процессе адаптации организма.
- Воздействие на организм низких температур
- Значение воспаления для организма
- Глава 2. Основные методы очищения организма
- ВОЗДЕЙСТВИЕ НА ОРГАНИЗМ ТЕРМИЧЕСКИХ ФАКТОРОВ
- Сохранение воды в организме
- Поступление воды в организм
- Факторы, влияющие на реактивность и резистентность организма