Гуморальная и клеточная иммунные системы.
Простейшие — фагоцитирующие эндосимбионты многоклеточных образований не могли при дальнейшем усложнении многоклеточной жизни защитить ее от случайного проникновения отдельных прокариотов и одноклеточных эукариотов.
Отработанные в течение миллиардов лет совместной эволюции механизмы лиганд-рецепторного узнавания между этими формами жизни продолжали действовать и в новых условиях. Благодаря им реликтовая иммунная система играла роль Троянского коня для многоклеточного орга-низма. Поэтому преимущество под давлением естественного отбора стали получать многоклеточные виды, у которых формировались взаимосвязанные системы по узнаванию и элиминации инфицированных фагоцитирующих клеток. У позвоночных они сложились еще в одну систему иммунитета, которую ученые условно делят на гуморальную и клеточную.
Паразит (микоплазма, бактерия или вирус), попав в организм позвоночного животного, встречает там уже не только своих «старых знакомых» по архейской эре — фагоцитирующие клетки, но и нечто качественно иное — иммунную систему — т. е. комплекс взаимосвязанных реакций органов и тканей макроорганизма на такое проникновение. И если судить по работе М. Marketon et аі. (2005), он «не узнает» клетки иммунной системы, эволюционировавшие уже в многоклеточных организмах. В селезенке мыши при экспериментальной чумной инфекции практически не поражаются T- и В-клетки, составляющие там большинство. Зато «в полном составе» гибнут все фагоцитирующие клетки. «В упор не замечает» лимфоциты и возбудитель натуральной оспы (McFadden G., 2004).
Паразит, сумевший «незамеченным» проникнуть в фагоцитирующую клетку, становится заметным иммунной системе, начав размножаться. Синтезирующиеся вирусные белки либо ЛПС бактерий образуют комплексы с поверхностными антигенами клетки (антигены гистосовместимости или трансплантационные антигены), а те распознаются Т-лимфоцитами.
Активировавшиеся Т-лимфоциты связываются своими рецепторами с инфицированной фагоцитирующей клеткой и убивают ее посредством апоптоза и особых литических белков — перфоринов и цитолизинов, поэтому такое звено иммунитета называют клеточным.Микроорганизм, сумевший размножиться в макрофаге, в дальнейшем имеет дело с неизвестными ему из архейской жизни клонами узкоспециализированных В-лимфоцитов, запускающих синтез антител, специфичных к его поверхностным белкам. Сам по себе В-лимфоцит для такого паразита безвреден, но связанный антителом, синтезированным В-лим- фоцитом, он легко становится «добычей» тех, кого привык использовать для собственного обитания, питания и размножения. Эту часть иммунных реакций (включая активизацию системы комплемента) исследователи традиционно относят к гуморальному иммунитету.
Общим для генетических систем, кодирующих антитела, рецепторы B- и Т-клеток и трансплантационные антигены, является их сложная интрон- экзонная организация и выраженная полигенность. Репертуар антител ученые затрудняются определить, считая, что он «насчитывает миллионы специфичностей». Ни одна другая генетическая система организма не имеет такого количества аллельных форм определенного гена, как транс-
плантационные антигены. В систему входят около 400 генов, обнаружено 3,6 млн строительных блоков ДНК, общее число фенотипов HLA составляет около 20 млрд. Системы не дублируют друг друга по специфичности распознавания антигенов. Сложная интрон-экзонная организация генов — это обычный результат деятельности ретроэлементов (см. рис. 7, 13 и 15). В следующей главе мы рассмотрим «ответы» иммунной системы на ретровирусы, подтверждающие причастность ретровирусов к созданию иммунной системы многоклеточных организмов вообще, и человека в частности. Но прежде чем перейти к столь непопулярной у разработчиков ВИЧ-вакцин теме, я хочу обратить внимание читателя на роль в инфекционных и эпидемических процессах, вызываемых ретровирусами, так называемых антиретровирусных систем (факторов) клеток.
Белки AID/APOBEC. Семейство AID/APOBEC включает белки, способные деаминировать цитозин на урацил у однонитевых полинуклеотидов. Они выполняют разные физиологические функции в клетке. АРОВЕС1 (обнаружен первым) редактирует посредством этого механизма мРНК аполипопротеина В (АроВ), внедряя преждевременный стоп-кодон и индуцируя продукцию «обрезанного» АроВ, который имеет уже другую функцию, Роль АРОВЕС2 остается неясной, но эволюционно он считается «родительским» для всего семейства (Cullen В. G., 2006).
Лктивационно-индуцируемая деаминаза (activation-induced deaminase, AID) также имеет древнее происхождение. Она функционирует в активированных В-клетках, где случайным образом редактирует дезоксицигидин (deoxycytidine) в дезоксиуридин (deoxyuridine) в локусах иммуноглобулиновых генов и является триггером активации процесса соматической гипермутации (somatic hypermutation, SHM) lg-вариабельного региона генов, кодирующих антиген-связывающие сайты. Если активность AID оказывается направленной на не Ig-гены, то это приводит к злокачественному перерождению В-клеток, но механизм, отвечающий за «нацеливание» SHM на Ig-гены, неизвестен (Ronai D. et аі., 2005).
Ген APOBEC 3G кодирует белок, который упаковывается в ретровирусные частицы, где дезаминирует цитозин на урацил в минус-цепи вирусной ДНК в процессе обратной транскрипции. В результате чего в плюс-цепи кДНК гуанозин заменяется аденином, и репликация ВИЧ останавливается. Этот механизм распространяется только на ВИЧ, лишенные гена белка Vif (фактор инфекционное™ вируса), и не влияет на продукцию вируса с полноценным Vif, Ген APOBEC 3G функционирует одинаковым образом как в Т-клетках-хелперах, так и в макрофагах (Wahl S. et аі., 2006).
Геном человека и других приматов кодирует по крайней мере пять, а возможно, и семь таких белков, сгруппированных в отдельный генный кла-
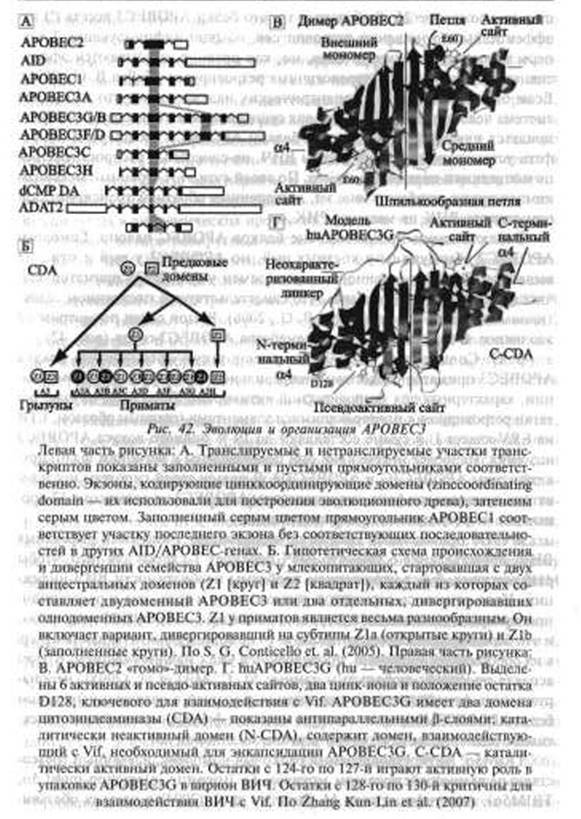
стер на хромосоме 22. Любопытно то, что белки АРОВЕСЗ всегда (!) неэффективны в отношении ретровирусов, обычно инфицирующих Т-хелперы и макрофаги данного вида, но, как правило, оказываются эффективными в отношении гетерологичных ретровирусов (Cullen В.
G., 2006). Если отказаться от антропоцентрических надежа на то, что иммунная система человека предназначена для существования только его самого, то придется признать, что благодаря белкам АРОВЕСЗ Т-хелперы и макрофаги устраняют мутантные формы ВИЧ, не способные распространяться по популяциям вида Homo sapiens. По своей сути этот странный механизм компенсирует ошибки в гене vif, допущенные обратной транскриптазой при синтезе ДНК на матрице РНК.Эволюционное происхождение белков APOBEC неясно. Гомологи АРОВЕС2 обнаружены у костных рыб, но АРОВЕСЗ у рыб и птиц не выявлено. Скорость несинонимичных замен у АРОВЕСЗ приматов значительно выше синонимичных, что свидетельствует о постоянном селективном давлении на них (Cullen В. G., 2006). В этой связи посмотрим на эволюцию и организацию всего семейства АРОВЕСЗ-генов (рис. 42).
S. G. Conticello et. аі. (2005) считают такую организацию локуса АРОВЕСЗ приматов следствием неправильного кроссовера и рекомбинации, характерных для ретровирусной активности. Во всяком случае, остатки ретровирусов с повторяющимися элементами (главным образом, LTR из ERV класса I) в сумме составляют до 19 % полного локуса АРОВЕСЗ человека. Наиболее интенсивно они представлены в регионах, фланкирующих гены APOBEC3G и АРОВЕСЗН. В пользу своей гипотезы исследователи приводят тот факт, что псевдогены АРОВЕСЗ на хромосоме 12 происходят от ретротранспозиционных актов.
В свете приведенных данных по элиминации Т-клетками дефектных ВИЧ, представление о том, что ВИЧ поражает Т-клетки для того, чтобы размножаться, будет упрощением проблемы необратимости ВИЧ-инфекции. Надо отнестись серьезно к возможности и того, что в эволюционном прошлом позвоночных ретровирусы «создали» АРОВЕСЗ «под себя», и эти белки продолжают играть какую-то не совсем понятную для нас роль в их собственной эволюции, происходящей уже с нашим участием. В этом аспекте проблемы любопытны данные М. С. Mikl et аі. (2005), необнаруживших фенотипические отклонения у мышей с разрушенным геном белка АРОВЕСЗ.
К тому же остается неясным фактор, оказывающий селективное давление на эти гены в течение миллионов лет.TRIMSa. Антиретровирусный белковый комплекс, названный трехсоставным взаимодействующим мотивом 5a (tripartite interaction motif 5α, TRlMSa), идентифицирован М. Stremlau et al. (2004) в клетках обезьян-

его взаимодействием с протеасомой и внести «сигналы*» для протеолиза посредством RING-зависимой аутоубиквитинилации (см. выше). Формирование комплекса «вирус — TRIM 5а» блокирует этап «раздевания» вируса и перенос его нуклеиновой кислоты в ядро клетки. Инфекция блокируется еще до обратной транскрипции вирусной РНК. Исследование TRIM5a у близкородственных приматов показало, что различия между ними, определяющие антивирусную специфичность, обнаруживаются в участках ВЗО.2-домена. Судя по высокому соотношению dN/dS, эти участки белка, соответствующие его поверхностным петлям, в ходе эволюции приматов подверглись позитивной селекции (Towers G. J., 2007).
Протеиновое семейство TRIM возникло в результате дупликации последовательности предкового гена, что характерно для активности ретроэлементов. Ген TRIM5 человека расположен на хромосоме 11 в пределах группы тесно связанных TRIM, включающих TRIMS, 6, 34 и 22. Эти TRIM либо не имеют, либо имеют относительно слабую антиретровирусную активность против близких ретровирусов (ВИЧ-1, ВИЧ-2, SIVmac, EIAV и MLV). Трудно сказать, почему это происходит. Возможно, они имеют какую-то другую, альтернативную функцию, или они просто неактивны против этой «выборки» ретровирусов. Во всяком случае, TRIM6, 22 и 34, непохожие на TRIM5, не имеют В30.2 доменов, что предполагает в эволюционном прошлом отсутствие на них того селективного давления, которому подвергался TRIM5 (Zhang F. et al., 2006; Li X. et al., 2007),
Взаимодействие между ВИЧ-1 и TRIMSa в клетках приматов Старого Света возможно благодаря циклофилину А (cyclophilm A, CypA).
CypA — пептидилпролилизомераза (peptidyl prolyl isomerase), выполняющая цис/ транс-изомеризацию (cis/trans isomerisation) пролиновых пептидных групп (proline peptide bonds) белков. CypA взаимодействует с молекулами gag лентивируса в инфицированных клетках, приводя их к рекрутингу в «нарождающиеся» вирионы (Thali М. et аі., 1994). CypA также взаимодействует с проникшим в клетку кором ВИЧ-1. И такое взаимодействие более важно для поддержания инфекционное™ вируса, чем взаимодействие циклофе- лина с «собирающимся» кором (Towers G. J. et al., 2003). CypA выполняет цис/транс-изомеризацию пептидных групп G89-P90 на наружной поверхности капсида ВИЧ. Эта активность пополняет цис-конформацию капсида и вовлекает капсид в рестрицирующийся комплекс с TRIM5a. Если активность CypA в клетке-мишени снижена, то TRIMSa не может взаимодействовать с капсидом, по большей части находящимся в лнрдке-конфор- мации (Towers G. J., 2007).Тот факт, что гены TRIM10, 15, 26, 27, 31, 38, 39 и 40 ассоциированы с главным комплексом гистосовместимости на хромосоме 6 и во время гриппозной инфекции их экспрессия усиливается, позволяет предположить возможность их участия в работе системы гуморального и клеточного иммунитета (Nisole S. et аі., 2005). Однако активность CypA не связана с активностью TRIM5a. Поэтому Z. Keckesova et аі. (2005) считают антиретровирусную активность TRIMSa не зависящей от реакций иммунной системы человека.
Оба известных антиретровирусных фактора, TRIMSa в комплексе с CypA и АРОВЕСЗ, защищают клетки только от «чужеродных» для них ретровирусов. В отношении же «своих» ретровирусов они являются фильтрами, гарантирующими виду, что другие экзогенные ретровирусы не могут составить им конкуренцию. Для вида Homo sapiens антиретровирусные факторы TRIMSa и АРОВЕСЗ являются гарантами успешности его инфицирования ВИЧ-1. C антропоцентрической точки зрения их существование с такой функцией бессмысленно. Однако если посмотреть на распространение ретровирусов в эволюционном аспекте проблемы, то смысл в поддержании естественным отбором TRIM5a и АРОВЕСЗ все же есть. Они обеспечивают условия для эндогенизации в зародышевую линию хозяина экзогенных ретровирусов, что происходит в ходе процесса, который мы понимаем как пандемический. Чем более масштабный характер принимает пандемия, тем больше генетическое разнообразие как самих ретровирусов, так и взаимодействующих с ними хозяев. Эндогенизация происходит случайно, но это такая же случайность, как и выстрел из револьвера при игре в «русскую рулетку», когда заполнение патронами барабана и нажатие на курок происходят со все возрастающей скоростью.
* * Ht
Эволюционное прошлое иммунной системы многоклеточных организмов свидетельствует о закреплении за ней естественным отбором резервуарной роли по отношению к ретровирусам. Поэтому можно утверждать, что эволюционный процесс среди многоклеточных организмов (по крайней мере, из царства животных) реализуется через клетки, которые мы относим к иммунной системе. В нашем восприятии времени мы видим только одну их функцию — защиту организма от антиген но-чуже родных веществ. Но благодаря клеткам иммунной системы происходит размножение и накопление экзогенных ретровирусов до какой-то критической массы, которая позволяет некоторым из них эндогенизироваться в зародышевой линии отдельных особей инфицированного вида, и в дальнейшем передаваться вертикально, меняя его эволюционную траекторию по механизму аллопатрического или симпатрического видообразования. ВИЧ/ СПИД-пандемия среди вида Homo sapiens — частное проявление этого процесса в эволюции таксона приматов.
Еще по теме Гуморальная и клеточная иммунные системы.:
- Методы изучения клеточного и гуморального иммунитета
- Клеточный и гуморальный иммунитет при послойной кератопластике
- Исследование циркулирующих иммунных комплексов гуморального звена иммунитета
- Оценка взаимосвязей между показателями системного и местного клеточного н гуморального иммунитета у часто болеющих детей с патологией ЛОР-органов.
- Нарушения систем, функционально сопряженных с иммунной системой
- Архейские предки иммунной системы человека
- Иммунная система
- ТЕМА 3: Органы иммунной системы.
- Экспериментальное воспроизведение гипофункции иммунной системы.
- 14.1. Иммунная система и опухолевый рост.
- I. Органы иммунной системы.
- Общие закономерности нарушений иммунной системы
- Глава 6. Патологическая физиология иммунной системы