Поступление воды в организм
Одни пассивные механизмы предотвращения потерь воды не могут обеспечить длительное выживание клещей даже при умеренных дефицитах насыщения. Тем не менее многие виды способны переживать в подобных неблагоприятных условиях по несколько месяцев, а у голодающих взрослых особей D.
reticulatus или А. ате- гісапит даже более 2 лет (табл. 10).Источниками поступления воды в организм являются жидкая пища, процессы метаболизма, поглощение капельной воды в результате питья и адсорбция водяных паров из атмосферы. По объему наибольшее количество воды поступает во время питания, но этот источник ограничен лишь 3 приемами пищи, которые разделены в жизненном цикле неделями или месяцами жизни вне организма прокормителя.
Метаболическая вода играет наибольшую роль в поддержании водного баланса у напитавшихся клещей между аполизисом и собственно линькой, а также у готовящихся к яйцекладке самок (Kahl, 1989). Так, у неперелинявших самок /. п- cinus, находившихся при 76 % влажности и температуре 20 °С, содержание воды в организме оставалось постоянным в течение нескольких недель до завершения линьки. У готовящихся к яйцекладке самок /. ricinus метаболическая вода компенсировала около 80 % потерь при 95 % влажности и температуре 15 °С, а при 34 % влажности и 15 °С — около 33 %.
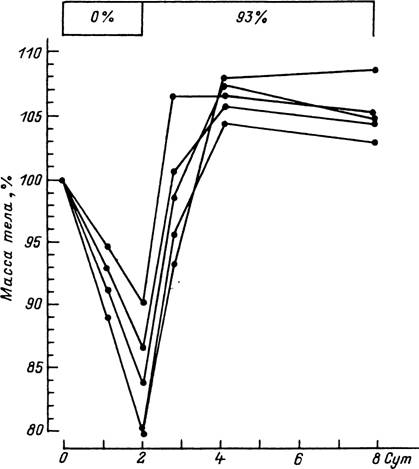
Рис. 76. Изменения массы тела у голодных нимф Hyalomma anatolicum, находившихся первые 2 сут опыта при 0 % относительной влажности и затем перенесенных в среду с относительной влажностью 93 %. Температура опыта 20 °С (по: Kahl, 1989).
Вопрос о способности клещей пить капельную воду и значении этого источника в их водном балансе требует дальнейших специальных исследований. Эта способность доказана только для ограниченного числа видов. В лабораторных условиях обнаружено активное питье воды личинками R.
evertsi, R. appendiculatus, A. heb- raeum, В. decoloratus, обитающими в ксерофильных ландшафтах Южной Африки (Londt, Whitehead, 1972), и В. microplus (Wilkinson, 1953). В других опытах с личинками D. variabilis и A. cajennense способность пить воду не обнаружена (Knulle, 1966).Главным источником воды для голодающих клещей остаются водяные пары, поглощение которых возможно при достаточно высокой относительной влажности воздуха. Впервые этот феномен был установлен для голодных взрослых I. ricinus (Lees, 1946), а в дальнейшем и для всех других, изученных в этом отношении видов (Knulle, Rudolph, 1982; Needham, Teel, 1991). Для каждого вида существует свое определенное значение относительной влажности воздуха (критическая влажность). В условиях более низкой влажности клещ непрерывно теряет воду, а при более высокой восполняет потери и поддерживает достаточный уровень содержания воды в организме за счет поглощения водяных паров (рис. 76). Величина критической влажности отражает экологические особенности вида и в первую очередь его требования к степени увлаженности воздуха в местообитаниях (табл. 14). У имаго наиболее гигрофильного I. ricinus этот показатель близок к 86—96 % относительной влажности, у A. cajennense — 90—92, у A. maculatum — 86—90, А. атегісапит — 80—88, у умеренно ксерофильных видов Dermacentor и Rhipi- cephalus — около 85—90 и у ксерофильных видов рода Hyalomma — 75—80 %. Величина критической влажности может отличаться у разных фаз развития одного вида, меняться в зависимости от физиологического состояния и возраста клещей.
Первоначально полагали, что напитавшиеся особи неспособны к поглощению влаги из воздуха, но недавно это явление было обнаружено у личинок и нимф /. ricinus, D. marginatus и H. punctata. У напитавшихся личинок и нимф вплоть до аполизиса (отделения гиподермы от старой кутикулы) происходит увеличение массы тела за счет поглощения водяных паров. После аполизиса на фаратной стадии масса тела убывает вплоть до освобождения от старой кутикулы (экдизиса).
Утрата способности к адсорбции воды у фаратных особей связана с дезинтеграцией старых слюнных желез, включая альвеолы I типа, и незавершенностью формирования новых альвеол (КаЫ, 1989).Поглощение водяных паров является физиологическим процессом, сопряжено с затратами энергии и возможно при температуре не ниже определенного порога, который для разных стадий I. ricinus, /. scapularis и Н. punctata при относительной влажности 95 % составляет 0—5 °С, а для голодных взрослых /. ricinus — даже -1 °С (КаЫ, 1989). С увеличением относительной влажности воздуха выше критической скорость адсорбции водяных паров возрастает до определенных пределов и далее сохраняется на одном уровне. Клещи, находящиеся в условиях высокой и постоянной влажности, также претерпевают циклические изменения содержания воды в организме. Например, у голодных самок A. variegatum 8—14-суточные периоды постепенного снижения содержания воды в организме сменялись 2—6- суточными периодами активного восстановления водного баланса, и при этом содержание воды в организме варьировало в пределах ±2 % (Rudolph, Knulle, 1978). У голодных имаго Н. asiaticum, утративших за счет испарения до 14 % своей первоначальной массы, после помещения в среду с влажностью 80 % водный баланс восстанавливался в первые 14 сут; а при 95 % влажности — за 1 сут. Общая масса воды, поглощенной клещами за это время, достигала 12 % от массы тела подсушенных особей (Балашов, 1960).
Таблица 14
Значения критической равновесной влажности некоторых видов иксодид
| Вид | Фаза | Критическая равновесная влажность (% относительной влажности) | Литературный источник |
| Ixodes ricinus | И | 86—96 | Lees, 1946 |
| Amblyomma атегісапит | И | >88 | Needham, Teel, 1991 |
| А. cajennense | Л | 80—85 | Knulle, 1966 |
| » » | И | 90—92 | Lees, 1946 |
| A. hebraeum | и | 80—86 | Osman, 1978 |
| A. lipidum | и | 84—91 | To же |
| A. limbatum | н | 80—83 | Bull, Smyth, 1973 |
| A. maculatum | и | >86 | Needham, Teel, 1991 |
| A. variegatum | и | 80—85 | Rudolph, Knulle, 1979 |
| Аропотта hydrosauri | н | 81—83 | Bull, Smyth, 1973 |
| Dermacentor andersoni | л | 80—85 | Knulle, 1966 |
| » » | и | 86—90 | Lees, 1946 |
| D. reticulatus | и | 86—88 | To же |
| D. variabilis | л | 80—85 | Knulle, 1966 |
| » » | и | 84—86 | Hair et al., 1975 |
| Haemaphysalis leporispalustris | л | 85 | Camin, Drenner, 1978 |
| Hyalomma asiaticum | и | 80 | Балашов, 1960 |
| H. dromedarii | л | 75 | Hafez et al., 1970 |
| н, и | 75—84 | ||
| H. schulzei | и | 80—85 | Knulle, Rudolph, 1982 |
| Rhipicephalus appendiculatus | л | 70 | Londt, Whitehead, 1972 |
| » » | н, и | 75—85 | Mongi, 1978 |
| R. bursa | н | 80—85 | Knulle, Rudolph, 1982 |
| R. pulchellus | н, и | 75—85 | Mongi, 1978 |
| R. sanguineus | и | 84—90 | Lees, 1946 |
Как мы уже отметили, голодные клещи, находящиеся на растительности в их естественных местообитаниях, совершают регулярные миграции. Они перемещаются из зоны с высоким дефицитом насыщения на траве или над поверхностью почвы в листовой опад и трещины почвы, где сохраняется более высокая влажность. Подобные миграции связаны с необходимостью восстановления водного баланса организма путем поглощения водяных паров. Лабораторными опытами было показано, что способность к переживанию повторных циклов обезвоживания—поглощения водяных паров различна у разных видов. Наибольшая пластичность отмечена у А. cajennense. При дегидратации при 0 % влажности до состояния обезвоживания и последующей гидратации при 96 % влажности самки этого вида переживали в среднем 4.4 ±1.28 цикла и максимально 12 циклов. У самцов отмечено до И циклов и в среднем 4 ±0.87 (Needham, Teel, 1991). У взрослых /. ricinus в природе в условиях Швейцарии в зависимости от микроклимата отмечено от 1 до 8 циклов вертикальной миграции (рис. 66) (Gigon, 1985).
Адсорбция водяных паров осуществляется внутри предротовой полости, в которую выделяются капли гигроскопичной слюны.
Эта слюна активно впитывает водяные пары из воздуха при значениях относительной влажности выше критической. Затем слюна повторно заглатывается или всасывается через специализированные участки кутикулы внутри ротовой полости (Rudolph, Knulle,1978). Микроосмотическими методами и по точке замерзания осмотический механизм адсорбции водяных паров был подтвержден для голодных самок А. атегісапит. Жидкое содержимое слюнного резервуара и латеральных выводных протоков у этих клещей было гиперосмотичным по отношению к гемолимфе (Sigal et al., 1991).Местом образования гиперосмотичной слюны служат альвеолы I типа в слюнных железах клеща (табл. XVI, 2; XIX, 4). По своему строению они сходны с осморегуляторными органами других животных. Цитоплазма их клеток разделена глубокими впячиваниями наружной цитоплазматической мембраны и множеством камер, внутри которых заключены многочисленные митохондрии. В процессе обезвоживания—гидратации организма ультраструктура митохондрий претерпевает характерные изменения, отражающие высокий уровень метаболической активности в процессе секреции гиперосмотичной слюны (Needham et al., 1990).
Еще по теме Поступление воды в организм:
- Пути поступления ядов в организм
- Поступление, распределение и выведение радиоактивных веществ из организма.
- Эффекты при повторном поступлении БАВ в организм
- Мероприятия, направленные на прекращение дальнейшего поступления ТХВ в организм
- Сохранение воды в организме
- Избыточное накопление воды в организме
- Потери воды организмом
- Алгоритмы обработки информации канала ультразвукового зондирования толщи воды (УЗК) и оптического локатора поверхности воды.
- Механизмы заражения воды в водопроводах; микрофлора водопроводной воды.
- Эффекты при совместном поступлении БАВ
- №3 Организм человека как единая биологическая система. Уровни организации человеческого организма: молекулярный, клеточный, тканевой, органный, системный, организменный.
- Пути поступления и токсичность