юПУЕИЯЙХЕ ОПЕДЙХ ХЛЛСММНИ ЯХЯРЕЛШ ВЕКНБЕЙЮ
кНЙСЯ ЦЕМНБ Rag]/2. яСОЕПЯЕЛЕИЯРБН ХЛЛСМНЦКНАСКХМНБ (Ig-SF). пЕКХЙРНБЮЪ ХЛЛСММЮЪ ЯХЯРЕЛЮ ВЕКНБЕЙЮ. ╚яАНПЙЮ╩ ЛМНЦНЙКЕРНВМШУ НПЦЮМХГЛНБ. рПЕРХИ ЮЙР РБНПЕМХЪ ≈ *ЙЕЛАПХИЯЙХИ БГПШБ╩. рНПЛНФЕМХЕ ЩБНКЧЖХХ.
цСЛНПЮКЭМЮЪ Х ЙКЕРНВМЮЪ ХЛЛСММШЕ ЯХЯРЕЛШ. аЕДЙХ AIDfAPOBEC. TRIMSa..нАШВМН Б ЛНМНЦПЮТХЪУ, НОХЯШБЮЧЫХУ ТСМЙЖХНМХПНБЮМХЕ ХЛЛСММНИ ЯХЯРЕЛШ ВЕКНБЕЙЮ (ЯЛ., МЮОПХЛЕП, НАЯРНЪРЕКЭМСЧ ПЮАНРС аЕКНЖЙНЦН я. л. Х юБРЮКХНМЮ 3. 3,, 2006), ЕЕ НПЦЮМХГЮЖХЪ ОПЕДЯРЮБКЪЕРЯЪ ГЮЙНМВЕММНИ Х ОПНДСЛЮММНИ ЯУЕЛНИ, Б ЯННРБЕРЯРБХХ Я ЙНРНПНИ БЯЕ ЕЕ ГБЕМЭЪ ДПСФМН ОПНРХБНЯРНЪР БПЮФДЕАМНЛС ОПНМХЙМНБЕМХЧ НОЮЯМШУ ЛХЙПННПЦЮМХГЛНБ. оНМХЛЮМХЕ ТСМЙЖХНМХПНБЮМХЪ ХЛЛСММНИ ЯХЯРЕЛШ ХЛЛСМНКНЦЮЛХ ОПЕДОНКЮЦЮЕР ХЯЙКЧВХРЕКЭМН ДНЯКНБМНЕ РНКЙНБЮМХЕ КЮР. РЕПЛХМЮ immunitas ≈ МЕБНЯОПХХЛВХБНЯРЭ НПЦЮМХГЛЮ ОН НРМНЬЕМХЧ Й БНГАСДХРЕКЪЛ ГЮПЮГМШУ АНКЕГМЕИ. оНДПЮГСЛЕБЮЕРЯЪ, ВРН ЦДЕ ХМТЕЙЖХЪ, РЮЛ Х ХЛЛСМХРЕР. нРЯЧДЮ ЯКЕДСЕР САЕФДЕМХЕ ЛМНЦХУ СВЕМШУ Б РНЛ, ВРН ХЛЛСММЮЪ ЯХЯРЕЛЮ ВЕКНБЕЙЮ АНПЕРЯЪ Я бхв-ХМТЕЙЖХЕИ, Х МЮДН РНКЭЙН ЕИ ОНЛНВЭ Б ЩРНИ АНПЭАЕ. нДМЮЙН ОПЮЙРХВЕЯЙХ ЯДЕКЮРЭ ЩРН МЕ СДЮЕРЯЪ. дКЪ ОНМХЛЮМХЪ ОПХВХМ ЩРХУ МЕСДЮВ МЮЛ МЮДН ГЮЦКЪМСРЭ Б ОПНЬКНЕ МЮЬЕИ ХЛЛСММНИ ЯХЯРЕЛШ.
кНЙСЯ ЦЕМНБ Ragl/2. щРН ЦЕМШ ТЕПЛЕМРНБ, ЪБКЪЧЫХУЯЪ ЙКЧВЕБШЛХ СВЮЯРМХЙЮЛХ V(D)J-ПЕЙНЛАХМЮЖХХ. рПЮДХЖХНММН ЯВХРЮЕРЯЪ, ВРН ЦСЛНПЮКЭМЮЪ ХЛЛСММЮЪ ЯХЯРЕЛЮ, Б НЯМНБЕ ЙНРНПНИ КЕФХР V(D)J-ПЕЙНЛАХМЮЖХЪ, ≈ ЩБНКЧЖХНММНЕ ОПХНАПЕРЕМХЕ ОНГБНМНВМШУ (ЯЛ. С йСОЕПЮ щ., 1980). лНФЕР АШРЭ, РЮЙ НМН Х ЕЯРЭ МЮ ЯЮЛНЛ ДЕКЕ,,МН БНР ЯЮЛ КНЙСЯ ЦЕМНБ Ragl/2 ХЛЕЕР ЦНПЮГДН АНКЕЕ ДПЕБМЕЕ ОПНХЯУНФДЕМХЕ, ВЕЛ ОНГБНМНВМШЕ, Х Б ЩБНКЧЖХНММШИ ОЕПХНД ╚ДН ОНГБНМНВМШУ╩ НМ ХЦПЮК ПНКЭ ПЕЙНЛАХМЮГШ ПЕРПН- БХПСЯНБ (Fugmann S. D. et ЮЁ., 2006).
нАМЮПСФЕМН ГМЮВХРЕКЭМНЕ ЯРПСЙРСПМНЕ ЯУНДЯРБН ЛЕФДС RAG-АЕКЙЮЛХ Х АЕКЙЮЛХ, ОПХМЮДКЕФЮЫХЛХ Й ЯСОЕПЯЕЛЕИЯРБС ХМРЕЦПЮГ ПЕРПНБХПСЯНБ. дКЪ МХУ СЯРЮМНБКЕМШ НАЫХЕ ЛЕУЮМХГЛШ ПЮЯЯЕВЕМХЪ ЖЕОЕИ дмй (НДМНЖЕОНВЕВМШИ ПЮГПШБ ВЕПЕГ ЦХДПНКХГ Х ОЕПЕМНЯ МХРХ ВЕПЕГ РПЮМЯЩЯРЕПХТХЙЮЖХЧ); ОНРПЕАМНЯРЭ Б ДХБЮКЕМРМШУ ХНМЮУ ЛЕРЮККНБ (Mg2+ ХКХ Mn2+); ЯОНЯНАМНЯРЭ ХЯОНКЭГНБЮРЭ ЮКЭРЕПМЮРХБМШЕ МСЙКЕНТХКШ, РЮЙХЕ ЙЮЙ ЮКЙНЦНКЭ; НАПЮГНБЮМХЕ ЬОХКЕЙ дмй Х ЯОНЯНАМНЯРЭ Й НАПЮРМНИ ПЕЮЙЖХХ (disintegration) (Polard P., Chandler M., 1995; Landree л.
A. et al., 1999).яСОЕПЯЕЛЕИЯРБН ХЛЛСМНЦКНАСКХМНБ (Ig-SF). щРН НЦПНЛМНЕ ЯЕЛЕИЯРБН АЕКЙНБ ЮДЦЕГХХ, МЮГБЮМХЕ ЙНРНПНЦН АНКЕЕ ХГБЕЯРМН ОН МЮГБЮМХЧ НДМНЦН ХГ ТЮЙРНПНБ ХЛЛСММНИ ЯХЯРЕЛШ ОНГБНМНВМШУ ≈ ХЛЛСМНЦКНАСКХМНБШУ ЮМРХРЕК, ЩБНКЧЖХНММН ОНЪБХБЬХУЯЪ Б ЩРНЛ ЯЕЛЕИЯРБЕ ОНЯКЕДМХЛХ. нЯМНБМШЛХ ЙПХРЕПХЪЛХ БЙКЧВЕМХЪ АЕКЙНБ Б ЯСОЕПЯЕЛЕИЯРБН ЪБКЪЧРЯЪ НОПЕДЕКЕММЮЪ ОПНЯРПЮМЯРБЕММЮЪ НПЦЮМХГЮЖХЪ ЛНКЕЙСК Х ЯРЮРХЯРХВЕЯЙХ ДНЯРНБЕПМЮЪ ЦНЛНКНЦХЪ Я ХГБЕЯРМШЛХ ХЛЛСМНЦКНАСКХМЮЛХ. йЮФДШИ ДНЛЕМ, БУНДЪЫХИ Б ЯНЯРЮБ ХЛЛСМНЦКНАСКХМЮ, ОПЕДЯРЮБКЪЕР ЯНАНИ ДБСУЯКНИМНЕ ЛНКЕЙСКЪПМНЕ НАПЮГНБЮМХЕ, ОНЯРПНЕММНЕ ОН ОПХМЖХОС МЕЯЙНКЭЙХУ ЮМРХОЮПЮККЕКЭ- МШУ п-ЯРПСЙРСП, ЯРЮАХКХГХПНБЮММШУ ЯБЪГЪЛХ ≈S≈S≈. рЮЙ НАПЮГСЕРЯЪ ЛНКЕЙСКЪПМЮЪ ЙНМТНПЛЮЖХНММЮЪ ЯРПСЙРСПЮ, ЯБНИЯРБЕММЮЪ РНКЭЙН ВКЕМЮЛ ЯСОЕПЯЕЛЕИЯРБЮ ХЛЛСМНЦКНАСКХМНБ. б ЮМЦКНЪГШВМНИ КХРЕПЮРСПЕ НМЮ ОНКСВХКЮ НАНГМЮВЕМХЕ Ig-fold (ХЛЛСМНЦКНАСКХМНБЮЪ ЯЙКЮДВЮРНЯРЭ) (ПХЯ. 36).
б ЮЯОЕЙРЕ ОНМХЛЮМХЪ ОПНХЯУНФДЕМХЪ ЯЮЛХУ АЕКЙНБ Ig-SF КЧАНОШРМН РН, ВРН Х НМХ МЕ ЪБКЪЧРЯЪ НПХЦХМЮКЭМШЛХ ЯРПСЙРСПЮЛХ, ОНЪБХБЬХЛХЯЪ ХЯЙКЧВХРЕКЭМН ДКЪ НЯСЫЕЯРБКЕМХЪ ПЮГМННАПЮГМШУ ЛЕУЮМХГЛНБ ЙКЕРНВМНИ ЮДЦЕГХХ. дЮ Х ОНЪБХРЭЯЪ ╚МХНРЙСДЮ╩ НМХ МЕ ЛНЦКХ, ОПХПНДЮ ЩЙНМНЛМЮ Б РНЛ, ВРН ЙЮЯЮЕРЯЪ АЮГНБШУ ЯРПСЙРСП ОНДДЕПФЮМХЪ ФХБНИ ЛЮРЕПХХ. хУ ОПНХЯ-
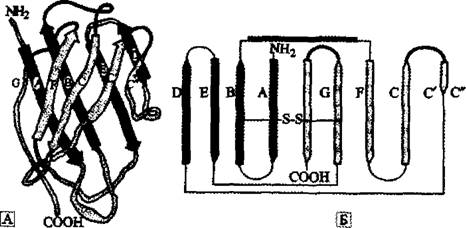
пХЯ. 36. яРПСЙРСПМЮЪ НПЦЮМХГЮЖХЪ Ѵ-ДНЛЕМЮ ОН ю. N. Barclay (7.999)
ю. дХЮЦПЮЛЛЮ ЙПХЯРЮККНЦПЮТХВЕЯЙНЦН ЮМЮКХГЮ Ѵ-ДНЛЕМЮ РЪФЕКНИ ЖЕОХ ЛХЕКНЛМНЦН АЕКЙЮ New ВЕКНБЕЙЮ, ДЕЛНМЯРПХПСЧЫЮЪ УЮПЮЙРЕП ДБСУЯКНИМНИ ЯЙКЮДВЮРНЯРХ РХОЮ Ig-fold ДЮММНЦН ДНЛЕМЮ. а. яУЕЛЮ ЯБЪГХ ЛЕФДС P-ЯРПСЙРСПМШЛХ ЯКНЪЛХ Х ╚-ЯОХПЮКЭМШЛХ ОНКНФЕМХЪЛ Ig-fold. вЕПМШЕ КХМХХ ≈ ЦХОЕПБЮПХЮАЕКЭМШЕ СВЮЯРЙХ (ПХЯСМНЙ ХГ ЙМХЦХ цЮКЮЙРХНМНБЮ б. ц., 2005)
УНФДЕМХЕ СУНДХР Й НВЕМЭ ДПЕБМХЛ АЕКЙЮЛ ЮДЦЕГХХ ≈ ЦХЯРНМЮЛ, НЯСЫЕЯРБКЪЧЫХЛ СОПЮБКЕМХЕ ПЮАНРНИ ЦЕМНБ Х ЯРЮАХКХГХПСЧЫХЛ БРНПХВМСЧ ЯРПСЙРСПС дмй Х УПНЛЮРХМЮ (ЯЛ. ПЮГД. 1.1). цХЯРНМШ СОПЮБКЪЧР ЯЙНПНЯРЭЧ V(D)J-ПЕЙНЛАХМЮЖХХ С ХМДХБХДССЛЮ (Espinoza я. et ЮЁ., 2005). Ho Ig-SF Б ╚ЦХЯРНМНБНЛ ЛХПЕ╩ ХЦПЮЕР АНКЕЕ ДПЕБМЧЧ ПНКЭ, ВЕЛ ЯЮЛХ ЦХЯРНМШ ≈ ПНКЭ ЬЮОЕПНМНБ, Р.
Е. ЯРПСЙРСП, ОНДДЕПФХБЮЧЫХУ ЙНМТНПЛЮЖХЧ ЦХЯРНМНБ Х НОПЕДЕКЪЧЫХУ УЮПЮЙРЕП ЦХЯРНМ/ЦХЯРНМНБНЦН БГЮХЛНДЕИЯРБХЪ (ПХЯ. 37).яКЕДНБЮРЕКЭМН, ЙНМТНПЛЮЖХНММЮЪ ЯРПСЙРСПЮ РХОЮ Ig-fold АЕПЕР ЯБНЕ МЮВЮКН Б ЩОНУС ОПНРНЙКЕРНВМШУ НАПЮГНБЮМХИ, ЙНЦДЮ ЙЮФДШИ ДНЛЕМ ЦХЯРНМНБНЦН АЕКЙЮ НАПЮГНБШБЮКЯЪ Б ПЕГСКЭРЮРЕ ПЕРПНРПЮМЯОНГХЖХНММНИ ЮЙРХБМНЯРХ ОЕПБШУ ПЕРПНЩКЕЛЕМРНБ. яТНПЛХПНБЮБЬХЕЯЪ МЮ НЯМНБЕ ЯРПСЙРСП РХОЮ Ig-fold ЦХДПНТНАМШЕ АЕКЙХ НЙЮГЮКХЯЭ БНЯРПЕАНБЮММШЛХ ЩБНКЧЖХЕИ ЙЮЙ ЯРПНХРЕКЭМШИ ЛЮРЕПХЮК ЛМНЦНЙКЕРНВМНЕ≥. оНЯПЕДЯРБНЛ ДСОКХЙЮЖХИ Х ОЕПЕЯРЮМНБНЙ ЦЕМНБ, ЙНДХПСЧЫХУ Ig-fold, ЯТНПЛХПНБЮКНЯЭ ЯСОЕПЯЕЛЕИЯРБН ХЛЛСМНЦКНАСКХМНБ.
яЕЦНДМЪ Й ЩРНЛС ЯЕЛЕИЯРБС НРМНЯЪРЯЪ ЛНКЕЙСКШ р-ЙКЯРНВМНЦН ЮМРХЦЕМ- ПЮЯОНГМЮЧЫЕЦН ЙНЛОКЕЙЯЮ, ЛНКЕЙСКШ I Х II ЙКЮЯЯНБ лмя, ЙНПЕЖЕОРНПШ р-ЙКЕРНЙ CD4 Х CD8, НДМНДНЛЕММШЕ АЕКЙХ ≈ Thy-Ι, β2'Μ> Po; ПЮГКХВМШЕ ЮДЦЕГХМШ Х ПЕЖЕОРНПМШЕ ЛНКЕЙСКШ, ЯОНЯНАЯРБСЧЫХЕ ЙНМРЮЙРМНЛС БГЮХЛНДЕИЯРБХЧ ХЛЛСМНЙНЛОЕРЕМРМШУ ЙКЕРНЙ ХКХ ЮДЯНПАЖХХ ПЮГКХВМШУ ЙКЮЯЯНБ ХЛЛСМНЦКНАСКХМНБ МЮ ЙКЕРНВМНИ ОНБЕПУМНЯРХ. мЮ КХЛТНХДМШУ ЙКЕРЙЮУ НДМЮ РПЕРЭ ОНБЕПУМНЯРМШУ ЛНКЕЙСК ≈ ЩРН ВКЕМШ ЯСОЕПЯЕЛЕИЯРБЮ ХЛЛСМНЦКНАСКХМНБ (цЮКЮЙРХНМНБ б. ц., 2005).
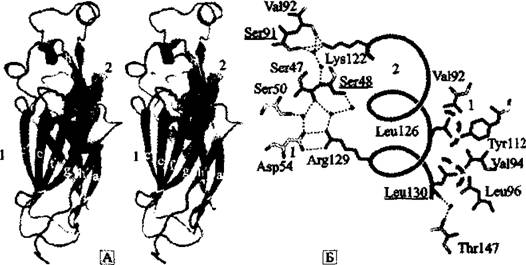
пХЯ. 37. Ig-SF ЬЮОЕПНМНБ ЦХЯРНМНБШУ АЕКЙНБ
ю. нАЫЮЪ ЯРПСЙРСПЮ ЬЮОЕПНМЮ AsflN, ЯБЪГШБЮЧЫЕЦН ЦХЯРНМНБШЕ АЕКЙХ мгЮг:
I ≈ AsflN, ЯБЪГШБЮЧЫХИЯЪ ЯН ЯОХПЮКЭЧ мгНЯг; 2 ≈ β-ЖЕОХ (НАНГМЮВЕМШ ОНЯПЕДЯРБНЛ МНЛЕМЙКЮРСПШ, ОПХМЪРНИ ДКЪ ОЕПЕЙКЧВЮЧЫЕЦНЯЪ РХОЮ Ig-ОНДНА- МНИ ЯЙКЮДВЮРНЯРХ; switched-type Ig-Itke folds). а. яРПСЙРСПМШЕ ДЕРЮКХ БГЮХЛНДЕИЯРБХЪ ЬЮОЕПНМЮ AsflN Я ЦХЯРНМНБШЛХ АЕКЙЮЛХ: I ≈ AsflN; 2 ≈ ЦХЯРНМНБШЕ АЕКЙХ мгЮг. БЮМДЕПБЮЮКЭЯНБШ БГЮХЛНДЕИЯРБХЪ ОНЙЮГЮМШ ОНКСЙПСФХЪЛХ. хНММНЕ ЯОЮПХБЮМХЕ Х БГЮХЛНДЕИЯРБХЕ ГЮ ЯВЕР НАПЮГНБЮМХЪ БНДНПНДМШУ ЯБЪГЕИ ОНЙЮГЮМШ ОСМЙРХПМШЛХ КХМХЪЛХ. оН ю. J. Antczak et ЮЁ. (2006)
мЮХАНКЕЕ ДПЕБМХЛХ Б ЩРНЛ ЯСОЕПЯЕЛЕИЯРБЕ ЪБКЪЧРЯЪ АЕКЙХ Po Х Thy-I1 РЮЙ ЙЮЙ НМХ ОПЕДЯРЮБКЪЧР ЯНАНИ НДМНДНЛЕММШЕ АЕКЙХ, МЕ ЯБЪГЮММШЕ МЮ ЙКЕРНВМНИ ОНБЕПУМНЯРХ Я ЙЮЙХЛХ-КХАН ДПСЦХЛХ.
оПХ ЩРНЛ ЦЕМ, ЙНМРПНКХПСЧЫХИ ЯХМРЕГ P0, ХЛЕЕР Б ЯПЕДМЕИ ВЮЯРХ ХМРПНМ. дЮММНЕ НАЯРНЪРЕКЭЯРБН, ОН ЛМЕМХЧ б. ц. цЮКЮЙРХНМНБЮ (2005), ОНГБНКЪЕР ОПЕДОНКНФХРЭ ЕЦН ОПНХЯУНФДЕМХЕ НР ЦЕМЮ ДКЪ ОНКСДНЛЕММНЦН ОЕОРХДЮ, ЯНЯРЮБКЪЧЫЕЦН НДХМ ХГ β-ЯРПСЙРСПМШУ ЯКНЕБ Б ЯНБПЕЛЕММШУ ДНЛЕМЮУ IgSF. б ПЕГСКЭРЮРЕ РЮМДЕЛМНИ ДСОКХЙЮЖХХ ЦЕМЮ ДКЪ МЮХАНКЕЕ ПЮММЕЦН ОНКСДНЛЕММНЦН ОПЕДЬЕЯРБЕММХЙЮ ЯТНПЛХПНБЮКЯЪ п0-ОНДНАМШИ АЕКНЙ. оПНЬЕДЬЮЪ ДЕКЕХХЪ ХМРПНММНИ ВЮЯРХ ЦЕМЮ P0 НАЕЯОЕВХКЮ БНГМХЙМНБЕМХЕ ЦЕМЮ-ОПЕДЬЕЯРБЕММХЙЮ ДКЪ ЮМРХЦЕМЮ Thy-1, щРН ЯНАШРХЕ ОПНХГНЬКН МЕ ЛЕМЕЕ 2 ЛКПД КЕЦ МЮГЮД. оНКХОЕОРХДШ, ЯЕПНКНЦХВЕЯЙХ ПНДЯРБЕММШЕ АЕКЙС Thy-I ЛКЕЙНОХРЮЧЫХУ, НАМЮПСФЕМШ С НДМНЙКЕРНВМШУ ЩСЙЮПХНР Х ОПНЙЮПХНР, ДНФДЕБШУ ВЕПБЕИ, ЦНКНБНМНЦХУ Х АПЧУНМНЦХУ ЛНККЧЯЙНБ, ПЕОРХКХИ, ПША, ОРХЖ. н ЦЕММНИ НПЦЮМХГЮЖХХ Х ПЮЯОПНЯРПЮМЕММНЯРХ Ig-SF ЛНФМН ОПНВХРЮРЭ Б ЙМХЦЕ б. ц. цЮКЮЙРХНМНБЮ (2005), ЛШ ФЕ ОНЙЮ НЯРЮМНБХЛЯЪ МЮ ЮМРХЦЕМЕ Thy-L еЦН ТСМЙЖХНМЮКЭМНЕ ОПЕДМЮГМЮВЕМХЕ РНВМН МЕ НОПЕДЕКЕМН. йПЮИМЕ ПЮММЕЕ ЕЦН ОНЪБКЕМХЕ Б ТХКНЦЕМЕГЕ ЯБЪГЮМН Я ОПНЖЕЯЯНЛ БНГМХЙМНБЕМХЪ ЛМНЦНЙКЕРНВМШУ НПЦЮМХГЛНБ, ЙНЦДЮ НМ БШЯРСОЮК Б ЙЮВЕЯРБЕ НДМНЦН ХГ ОНБЕПУМНЯРМШУ ЛНКЕЙСКЪПМШУ ТЮЙРНПНБ ЛЕФЙКЕРНВМНИ ЮДЦЕГХХ, ХЯОНКЭГСЪ ЛЕУЮМХГЛ ЦНЛНТХКЭМНЦН ЛЕФЙКЕРНВМНЦН БГЮХЛНДЕИЯРБХЪ, Р. Е. ЙНМРЮЙРМНЦН БГЮХЛНДЕИЯРБХЪ ╚ЯБНЕЦН╩ ЯН ╚ЯБНХЛ╩ (Barclay A. N., 1999).пЕКХЙРНБЮЪ ХЛЛСММЮЪ ЯХЯРЕЛЮ ВЕКНБЕЙЮ. б ПЮГД. 2.1 (╚оПНЯРЕИЬХЕ Х ХУ ОЮПЮГХРШ╩) ОПХБЕДЕМШ ЩЙЯОЕПХЛЕМРЮКЭМШЕ ДЮММШЕ S. Blazquez et ЮЁ. (2006), ОНЙЮГШБЮЧЫХЕ ЯСЫЕЯРБНБЮМХЕ НРБЕРМШУ ПЕЮЙЖХИ С ЮЛЕА МЮ ТЮЙРНП МЕЙПНГЮ НОСУНКЕИ (TNF), ХМРЕПКЕИЙХМ-1 П (IL-Ιβ), ХМРЕПКЕИЙХМ-8 (1L-8) Х ЖХЙКННЙЯХЦЕМЮГС 2, ЮМЮКНЦХВМШЕ РЮЙНБШЛ С МЕИРПНТХКНБ Х ЛЮЙПНТЮЦНБ ЛМНЦНЙКЕРНВМШУ НПЦЮМХГЛНБ. яКЕДНБЮРЕКЭМН, МЕ РНКЭЙН ЩРХ УЕЛНЙРШ Х ЖХРНЙХМШ ЯРЮКХ ╚ЯПЕДЯРБНЛ НАЫЕМХЪ╩ ЛЕФДС ЩСЙЮПХНРЮЛХ-ОПНЯРЕИХЬЛХ ГЮДНКЦН ДН ОНЪБКЕМХЪ МЕИРПНТХКНБ Х ЛЮЙПНТЮЦНБ ЙЮЙ ЙКЕРНЙ ХЛЛСММНИ ЯХЯРЕЛШ ОНГБНМНВМШУ НПЦЮМХГЛНБ, МН Х ПЕЖЕОРНПШ, ОНЯПЕДЯРБНЛ ЙНРНПШУ НЯСЫЕЯРБКЪКНЯЭ БГЮХЛНДЕИЯРБХЕ ОПНЯРЕИЬХУ. щРХ ФЕ ПЕЖЕОРНПШ ГЮ ЛХККХЮПДШ КЕР ЯНБЛЕЯРМНИ ЩБНКЧЖХХ МЮСВХКХЯЭ ХЯОНКЭГНБЮРЭ БМСРПХЙКЕРНВМШЕ ОПНЙЮПХНРХВЕЯЙХЕ ОЮПЮГХРШ ОПНЯРЕИЬХУ. б УНДЕ ЩБНКЧЖХХ ЛМНЦНЙКЕРНВМШУ ТНПЛ ФХГМХ НМХ АШКХ СМЮЯКЕДНБЮМШ ╚БЯЕЛ ОЮЙЕРНЛ╩.
уЕЛНЙХМШ. рЕПЛХМ ╚УЕЛНЙХМ╩ АШК ОПХЛЕМЕМ Й ЩРХЛ ЛНКЕЙСКЮЛ, ОНЯЙНКЭЙС ЯВХРЮКХ, ВРН ХУ НЯМНБМЮЪ АХНКНЦХВЕЯЙЮЪ ЮЙРХБМНЯРЭ ЪБКЪЕРЯЪ УЕЛНРЮЙЯХВЕЯЙНИ, Р, Е., МЮОПЮБКЪЧЫЕИ ДБХФЕМХЕ ЙКЕРНЙ ОН ЦПЮДХЕМРЮЛ ЙНМЖЕМРПЮЖХХ УЕЛНЙХМНБ БН БПЕЛЪ БНЯОЮКХРЕКЭМШУ ПЕЮЙЖХИ. уНРЪ УЕЛНЙХМШ ОПЕДЯРЮБКЪЧР РНКЭЙН НДХМ ЙКЮЯЯ ЛМНЦХУ РХОНБ ХГБЕЯРМШУ УЕЛНРЮЙР ХМНЕ (chemotactins), БЙКЧВЮЧЫХУ ЬХПНЙХИ ЛНКЕЙСКЪПМШИ ЯОЕЙРП БЕЫЕЯРБ НР КХОХДНБ ДН МСЙКЕНРХДНБ, НМХ БШЦНДМН НРКХВЮЧРЯЪ НР НЯРЮКЭМШУ АКЮЦНДЮПЪ ЯБНЕИ ЛНКЕЙСКЪПМНИ ЯРЮАХКЭМНЯРХ Х ЯОЕЖХТХВМНЯРХ ЙКЕРНЙ-ЛХЬЕМЕИ. кЧАНОШРМН Х РН, ВРН УЕЛНЙХМШ ОПЕДЯРЮБКЪЧР ЯНАНИ ЦНЛНКНЦХВМНЕ ЯСОЕПЯЕЛЕИЯРБН ЯПЮБМХРЕКЭМН МЕАНКЭЬХУ АЕКЙНБ НР 8 ДН 17 ЙдЮ, ОНЪБХБЬЕЦНЯЪ Б ПЕГСКЭРЮРЕ ДСОКХЙЮЖХХ Х ЛНДХТХЙЮЖХХ НДМНЦН ЮМЖЕЯРПЮКЭ- МНЦН ЦЕМЮ (Chensue S. W., 2001).
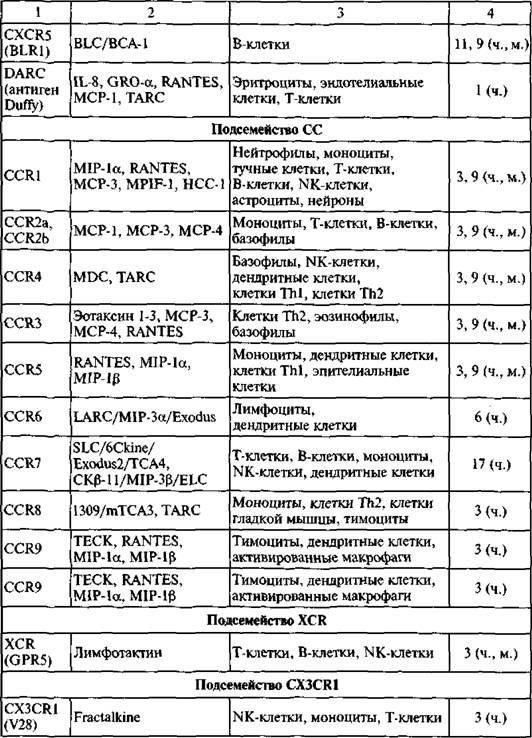
яПЮБМХРЕКЭМШИ ЮМЮКХГ ЮЛХМНЙХЯКНРМШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ ЛКЕЙНОХРЮЧЫХУ Х ОПХЛХРХБМШУ ОНГБНМНВМШУ ОНЙЮГШБЮЕР, ВРН ЦПСООШ CXC- Х яя-УЕЛНЙХМНБ ПЮГНЬКХЯЭ НР ОПЕДЙНБНЦН ЦЕМЮ ЕЫЕ ДН ОНЪБКЕМХЪ ЯЮЛХУ ОНГБНМНВМШУ (Najakshin ю. л. et ЮЁ., 1999) (ПХЯ. 38).
пЕЖЕОРНПШ УЕЛНЙХМНБ. бОЕПБШЕ НМХ АШКХ НОХЯЮМШ МЮ ╚НОЕПЕФЮЧЫЕЛ НАЗЕЙРЕ╩ РЮЙХУ ХЯЯКЕДНБЮМХИ, МЮ КЕИЙНЖХРЮУ. бОНЯКЕДЯРБХХ ПЕЖЕОРНПШ УЕЛНЙХМНБ НАМЮПСФЕМШ МЮ ЩМДНДЕПЛЮКЭМШУ, ЛЕГЕМУХЛМШУ, ЩЙРНДЕПЛЮКЭМШУ Х МЕИПНЩЙРНДЕПЛЮКЭМШУ ЙКЕРЙЮУ ЛМНЦНЙКЕРНВМШУ НПЦЮМХГЛНБ. нДМЮЙН ЕЯКХ ЯНОНЯРЮБХРЭ ДЮММШЕ, ОПХБЕДЕММШЕ МЮ ПХЯ. 38, Я ДЮММШЛХ, НАНАЫЕММШЛХ Б РЮАК. 9, РН ЯКЕДСЕР ЯДЕКЮРЭ БШБНД, ВРН НЯМНБМШЕ ОНДЯЕЛЕИЯРБЮ КХЛТНЙХМНБ, БГЮХЛНДЕИЯРБСЧЫХУ Я ЙКЕРЙЮЛХ ЯНБПЕЛЕММШУ ОНГБНМНВМШУ,
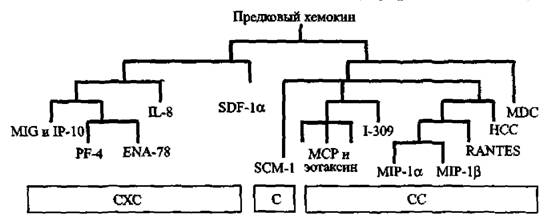
пХЯ. 38. щБНКЧЖХЪ ЯЕЛЕИЯРБ УЕЛНЙХМНБ
яСОЕПЯЕЛЕИЯРБН УЕЛНЙХМНБ ДЕКХРЯЪ МЮ ОНДЙКЮЯЯШ (ОНДЯЕЛЕИЯРБЮ) МЮ НЯМНБЕ ПЮЯОНКНФЕМХЪ ЖХЯРЕХМНБШУ НЯРЮРЙНБ, КНЙЮКХГНБЮММШУ Б N-РЕПЛХМЮКЭМНИ НАКЮЯРХ ЩРХУ ЛНКЕЙСК. оНДЙКЮЯЯШ НАНГМЮВЕМШ ЙЮЙ я, яя, CXC Х яуууя, ЦДЕ C НГМЮВЮЕР ЙНКХВЕЯРБН ЖХЯРЕХМНБШУ НЯРЮРЙНБ Б N-РЕПЛХМЮКЭМНИ НАКЮЯРХ, Ю X ОПЕДЯРЮБКЪЕР ЙНКХВЕЯРБН КЕФЮЫХУ ЛЕФДС МХЛХ ЮЛХМНЙХЯКНР. оНДЯЕЛЕИЯРБН CXC ПЮГДЕКЪЕРЯЪ ХМНЦДЮ ЕЬЕ МЮ РХОШ ELR Х non-ELR МЮ НЯМНБЮМХХ ОПХЯСРЯРБХЪ ХКХ НРЯСРЯРБХЪ ОНЯКЕДНБЮРЕКЭМНЯРХ ХГ РПЕУ ЮЛХМНЙХЯКНР (цКС-кЕИ-юПЦ), ЙНРНПЮЪ ОПЕДЬЕЯРБСЕР ОЕПБНЛС ЖХЯРЕХМНБНЛС ЮЛХМНЙХЯКНРМНЛС НЯРЮРЙС ОЕПБХВМНИ ЯРПСЙРСПШ ЩРХУ УЕЛНЙХМНБ.
оПХЯСРЯРБХЕ ОНЯКЕДНБЮРЕКЭМНЯРХ ЯННАЫЮЕР ЮМЦХНЦЕММСЧ ТСМЙЖХЧ ЩРНЛС РХОС УЕЛНЙХМНБ, РНЦДЮ ЙЮЙ ELR-НРПХЖЮРЕКЭМШЕ УЕЛНЙХМШ НАМЮПСФХБЮЧР ЮМЦХНЯРЮРХВЕЯЙХЕ ЯБНИЯРБЮ. бЯЕ НЯМНБМШЕ ОНДЯЕЛЕИЯРБЮ УЕЛНЙХМНБ ЯТНПЛХПНБЮКХЯЭ ЕЫЕ ДН ОНЪБКЕМХЪ ОНГБНМНВМШУ ФХБНРМШУ. оН ю. л, Najakskin ei al. (1999)пЕЖЕОРНПШ УЕЛНЙХМНБ*
рЮАКХЖЮ 9
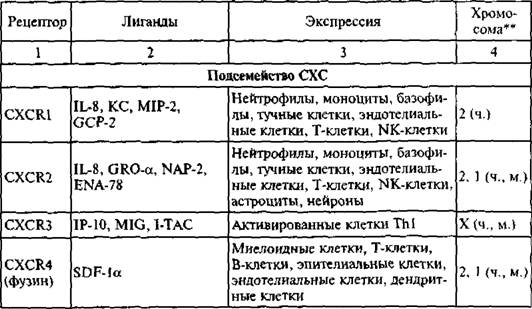
В
оПНДНКФЕМХЕ РЮАК. 9
[1] оН S, W. Chensue (2001).
[1] В, ≈ ВЕКНБЕВЕЯЙХЕ; Л. ≈ ЛШЬХМШЕ.
Ц
Х ЯННРБЕРЯРБЕММН ХУ ПЕЖЕОРНПШ ЯСЫЕЯРБНБЮКХ ЕЫЕ ДН ЯТНПЛХПНБЮМХЪ ЩРНЦН РЮЙЯНМЮ, Х ОПХ НОПЕДЕКЕММНИ МЮЯРНИВХБНЯРХ ХУ БЯЕУ ЛНФМН МЮИРХ Х С ОПНЯРЕИЬХУ. пЮАНРЮ S. Blazquez et Ю). (2006) ≈ ЩРН ЯХЦМЮК Н ╚ЯНГПЕБЮЧЫЕЛ╩ МЮОПЮБКЕМХХ РЮЙХУ ХЯЯКЕДНБЮМХИ.
бЯЕ УЕЛНЙХМШ НЙЮГШБЮЧР ДЕИЯРБХЕ МЮ ТЮЦНЖХРХПСЧЫХЕ ЙКЕРЙХ ОН НДМНЛС ЛЕУЮМХГЛС ≈ ВЕПЕГ ПЕЖЕОРНПШ, ЯБЪГЮММШЕ Я ЦСЮМНГХММСЙКЕНРХДНЛ- АЕКЙНЛ (GPCR). оНДНАМН БЯЕЛ ПЕЖЕОРНПЮЛ, ЯБЪГЮММШЛ Я GPCR, ПЕЖЕОРНПШ УЕЛНЙХМНБ ХЛЕЧР ЯЕЛЭ РПЮМЯЛЕЛАПЮММШУ ЦХДПНТНАМШУ ДНЛЕМНБ Я РПЕЛЪ БМСРПХЙКЕРНВМШЛХ Х РПЕЛЪ БМЕЙКЕРНВМШЛХ ЦХДПНТХКЭМШЛХ ОЕРКЪЛХ. оНРЕМЖХЮКЭМН ЦКХЙНГХКХПНБЮММЮЪ БМЕЙКЕРНВМЮЪ ЮЛХМНРЕПЛХМЮКЭМЮЪ НАКЮЯРЭ СВЮЯРБСЕР Б ЯБЪГШБЮМХХ УЕЛНЙХМЮ, РНЦДЮ ЙЮЙ БМСРПХЙКЕРНВМЮЪ ЙЮПАНЙЯХРЕПЛХМЮКЭМЮЪ НАКЮЯРЭ СВЮЯРБСЕР Б ЯБЪГШБЮМХХ Я АЕКЙНЛ G Х ОНДБЕПФЕМЮ ПЕЦСКЪРНПМНЛС ТНЯТНПХКХПНБЮМХЧ. юЙРХБЮЖХЪ ПЕЖЕОРНПНБ УЕЛНЙХМНБ МЮВХМЮЕРЯЪ ЯН ЯБЪГШБЮМХЪ Я БМЕЙКЕРНВМШЛ КХЦЮМДНЛ, ВРН ХМХЖХХПСЕР БГЮХЛНДЕИЯРБХЕ Я БМСРПХЙКЕРНВМШЛХ ОНЙНЪЫХЛХЯЪ цдт-МЕЯСЫХЛХ РПХЛЕПМШЛХ АЕКЙЮЛХ G. щРН ОПХБНДХР Й НАЛЕМС цдт МЮ црт, БШГШБЮЪ ПЮЯОЮД АЕКЙЮ G МЮ ЯСАЗЕДХМХЖШ α-G Х β/γ-G. оНЯКЕДМЪЪ ЯСАЗЕДХМХЖЮ, Б ЯБНЧ НВЕПЕДЭ, ЮЙРХБХПСЕР РЮЙХЕ ТЕПЛЕМРШ ЙЮЙ ТНЯТНКХОЮГЮ C Х ТНЯТНХМНГХРХД-3-ЙХМЮ- ГЮ, ЙНРНПШЕ ЙНМБЕПРХПСЧР ТНЯТНРХДХКХМНГХРНК-4,5-ДХТНЯТЮР (IP2) Б ТНЯ- ТНЬЖХКХМНГХРНК-1,4,5-РПХТНЯТЮР (IPj) Х ДХЮЖХКЦКХЖЕПХМ (DAG), IP3 ЯРХЛСКХПСЕР ОПХРНЙ ХНМНБ ЙЮКЭЖХЪ, Х DAG ЮЙРХБХПСЕР ХГНТНПЛШ ОПНРЕХМ- ЙХМЮГШ C (пй.я). рЮЙХЛ НАПЮГНЛ, ОНДЦНРЮБКХБЮЕРЯЪ БМСРПХЙКЕРНВМЮЪ ЯПЕДЮ ДКЪ ТНЯТНПХКХПНБЮМХЪ Я СВЮЯРХЕЛ ПЪДЮ ЙХМЮГ (МЮОПХЛЕП, ЮЙРХБХПНБЮММЮЪ ЛХРНЦЕМНЛ ОПНРЕХМЙХМЮГЮ, ОПНРЕХМРПХОЯХМЙХМЮГЮ) Х МЕАНКЭЬХУ црт-ЮГ (МЮОПХЛЕП, Ras Х Rho), ЙНРНПШЕ Б ЙНМЕВМНЛ ЯВЕРЕ НЙЮГШБЮЧР БКХЪМХЕ МЮ ЙКЕРНВМШЕ ТСМЙЖХХ, РЮЙХЕ ЙЮЙ ЮДЦЕГХЪ, УЕЛНРЮЙЯХЯ, ДЕЦПЮМСКЪЖХЪ Х БГПШБ ПЕЯОХПЮРНПМНИ ЮЙРХБМНЯРХ (Luttrell L. л. et ЮЁ., 1997).
аЕКЙХ РПЮМЯДСЙЖХХ ЯХЦМЮКНБ. хМРЕПЕЯМШЛ ЮЯОЕЙРНЛ АХНКНЦХХ ЛЮЙПНТЮЦНБ АШКН НАМЮПСФЕМХЕ ЯБЪГХ ХУ ПЕЖЕОРНПНБ Я АЕКЙЮЛХ G, ОПЕДЯРЮБКЪЧЫХЛХ АНКЭЬНИ ЙКЮЯЯ АЕКЙНБ РПЮМЯДСЙЖХХ ЙКЕРНВМШУ ЯХЦМЮКНБ (Locati M., Murphy п. M., 1999). мН Х С ЩРХУ АЕКЙНБ ЩБНКЧЖХНММЮЪ ХЯРНПХЪ МЮВХМЮЕРЯЪ Б ЮПУЕЕ. юМЮКХГ ОНЯКЕДНБЮРЕКЭМНЯРЕИ α-ЯСАЗЕДХМХЖ ЦЕРЕПНРПХЛЕПМН- ЦН G-АЕКЙЮ Protozoa, ОПНБЕДЕММШИ D. я. New Х J. р. Wong (1998), ОНЙЮГЮК , ВРН НМХ ХЛЕЧР МЕ РНКЭЙН НАЬХПМСЧ ЦНЛНКНЦХЧ ЯН ЯБНХЛХ ДБНИМХЙЮЛХ С ЛКЕЙНОХРЮЧЫХУ, МН Х ЮАЯНКЧРМСЧ ЦНЛНКНЦХЧ ОНЯКЕДНБЮРЕКЭМНЯРЕИ МЮХАНКЕЕ ГМЮВХЛШУ ТСМЙЖХНМЮКЭМШУ ПЕЦХНМНБ ЩРХУ АЕКЙНБ.
GPCR НРМНЯЪРЯЪ Й ЯСОЕПЯЕЛЕИЯРБС ПНДЯРБЕММШУ ПЕЖЕОРНПНБ, СВЮЯРБСЧЫХУ Б РПЮМЯДСЙЖХХ ЬХПНЙНЦН ЯОЕЙРПЮ БМЕЙКЕРНВМШУ ЯХЦМЮКЭМШУ ЛНКЕЙСК, РЮЙХУ ЙЮЙ ЦНПЛНМШ, МЕИПНРПЮМЯЛХРРЕПШ, ЖХРНЙХМШ (ХМРЕПКЕИЙХМШ, ТЮЙРНПШ МЕЙПНГЮ НОСУНКХ Х ХМРЕПТЕПНМШ ≈ ЯОНЯНАМШ ПЕЦСКХПНБЮРЭ ЩЙЯОПЕЯЯХЧ
УЕЛНЙХМНБ Х МЮНАНПНР), ЮПНЛЮРХГЮРНПШ, ЯБЕР Х ДП. пЕЖЕОРНПШ УЕЛНЙХМНБ ХЛЕЧР Д КХМС НР 320 ДН 3S0 ЮЛХМНЙХЯКНР. рЮЙФЕ ЙЮЙ Х ЯЮЛХ УЕЛНЙХМШ, НМХ НАМЮПСФХБЮЧР ГМЮВХРЕКЭМСЧ ЦНЛНКНЦХЧ ОНЯКЕДНБЮРЕКЭМНЯРЕИ. бЯЕЦН ХУ АНКЕЕ РШЯЪВХ, Х ХУ ПЮЯЯЛНРПЕМХЕ МЕ БУНДХР Б ГЮДЮВС ЩРНИ ПЮАНРШ (АНКЕЕ ОНДПНАМН Н ПЕЖЕОРНПЮУ УЕЛНЙХМНБ ЯЛ. С Chensue S. W., 2001).
йЮЙ ╚МЕ ОН СВЕАМХЙС╩ Б НРМНЬЕМХХ МЕЙНРНПШУ АЮЙРЕПХИ Х БХПСЯНБ БЕДСР ЯЕАЪ ЛНАХКХГНБЮММШЕ УЕЛНЙХМЮЛХ ТЮЦНЖХРХПСЧЫХЕ ЙКЕРЙХ ОНГБНМНВМШУ, ЛШ СФЕ ЙПЮРЙН ПЮЯЯЛНРПЕКХ БШЬЕ (ЯЛ. ПЮГЮ. 2.1). мН БШАПНЯШ ТЮЦНЖХРХПСЧЫХЛХ ЙКЕРЙЮЛХ УЕЛНЙХМНБ ДЮЧР Х ДПСЦХЕ ЩТТЕЙРШ, НАШВМН МЕ БЙКЧВЮЕЛШЕ Б НДМНГМЮВМШЕ ЯУЕЛШ ТСМЙЖХНМХПНБЮМХЪ ХЛЛСММНИ ЯХЯРЕЛШ, ОПЕДМЮГМЮВЕММШЕ ДКЪ ЯРСДЕМРНБ. мЮОПХЛЕП, ЩРХНКНЦХВЕЯЙХИ ЮЦЕМР АНКЕГМХ кЮИЛЮ, Borrelia burgdorferi, ЯРХЛСКХПСЕР ЯХМРЕГ MIP-lcc, IL-8, GRO-oc, MCP-I Х RANTES ВЕКНБЕВЕЯЙХЛХ ЛНМНЖХРЮЛХ. мН ХУ ОНЪБКЕМХЕ Б ЙПНБХ Б ОНБШЬЕММШУ ЙНКХВЕЯРБЮУ МЕ РНКЭЙН МЕ ОПХБНДХР Й ЩТТЕЙРХБМНИ ЩКХЛХМЮЖХХ ЛЮЙПНТЮЦЮЛХ АНППЕКХИ ХГ НПЦЮМХГЛЮ ВЕКНБЕЙЮ, МН Х БШГШБЮЕР ОНБПЕФДЕМХЕ ЕЦН РЙЮМЕИ Х УПНМХВЕЯЙХИ ЮПРПХР (Sprenger м. et ЮЁ., 1997). м. Yoneyama Я ЯНРП. (1998) ОПНДЕЛНМЯРПХПНБЮКХ, ВРН БМЕГЮОМН Х АШЯРПН ПЮГБХБЮЧЫХИЯЪ ЦЕОЮРХР, ХМДСЖХПНБЮММШИ Propionibacterium acnes Х ЩМДНРНЙЯХМНЛ, ГЮБХЯЕК НР TARC ≈ КХЦЮМДЮ CCR4, НРБЕРЯРБЕММНЦН ГЮ ЛНАХКХГЮЖХЧ ОЕВЕМНВМШУ CD4+ р-ЙКЕРНЙ CCR4+ УНГЪХМЮ. лНАХКХГЮЖХЪ ХЛЛСММШУ ЙКЕРНЙ ЮЯЯНЖХХПНБЮКЮЯЭ Я ОЕВЕМНВМНИ МЕДНЯРЮРНВМНЯРЭЧ, ОПХБНДЪЫЕИ Й КЕРЮКЭМШЛ ХЯУНДЮЛ. гДЕЯЭ БЮФМН НРЛЕРХРЭ Х ДПСЦСЧ ЯРНПНМС ОНБПЕФДЕМХЪ ОЮПЮГХРХВЕЯЙХЛ НПЦЮМХГЛНЛ РЙЮМЕИ УНГЪХМЮ Х ЯКЕДСЧЫЕЦН ГЮ ЩРХЛ КЕРЮКЭМНЦН ХЯУНДЮ АНКЕГМХ ≈ НМХ НЙЮГШБЮЧРЯЪ ЯНБЕПЬЕММН АЕЯЯЛШЯКЕММШЛХ ДКЪ ОНДДЕПФЮМХЪ Б ОПХПНДЕ ЯЮЛНЦН ОЮПЮГХРЮ.
мЕР МЕНАУНДХЛНЯРХ ОПХБНДХРЭ РЮЙХЕ МЮАКЧДЕМХЪ ДЮКЭЬЕ. хУ АНКЕЕ ВЕЛ ДНЯРЮРНВМН, ВРН АШ СЯНЛМХРЭЯЪ Б ╚ПЮГСЛМНЛ СЯРПНИЯРБЕ╩ (Я РНВЙХ ГПЕМХЪ ВЕКНБЕЙЮ, ЙНМЕВМН) НПЦЮМХГЮЖХХ МЮЬЕИ ХЛЛСММНИ ЯХЯРЕЛШ МЮ СПНБМЕ БГЮХЛНДЕИЯРБХЪ ТЮЦНЖХРХПСЧЫХУ ЙКЕРНЙ ЛЕФДС ЯНАНИ Х Я ОЮПЮГХРХВЕЯЙХЛХ ЛХЙПННПЦЮМХГЛЮЛХ. дЮ Х ЯЮЛХ ОЮПЮГХРШ МЕ ЛНЦСР ╚ОНУБЮЯРЮРЭ╩ ЖЕКЕЯННАПЮГМШЛ ОНБЕДЕМХЕЛ Б НРМНЬЕМХХ ЛЮЙПНТЮЦНБ.
бОПНВЕЛ, ЩРС ХЛЛСММСЧ ЯХЯРЕЛС ЯОЕЖХЮКЭМН МХЙРН Х МЕ ЯНГДЮБЮК ОН ЙЮЙНЛС-РН ГЮПЮМЕЕ ОПНДСЛЮММНЛС ОКЮМС, ОПЕДОНКЮЦЮЪ, ВРН ЕЕ НЯМНБМНЕ ОПЕДМЮГМЮВЕМХЕ ЯНЯРНХР Б ЩКХЛХМЮЖХХ ОЮРНЦЕММШУ ЛХЙПННПЦЮМХГЛНБ. нМЮ ЯКСВЮИМН ЯКНФХКЮЯЭ ХГ ЙКЕРНЙ, ОЕПБНМЮВЮКЭМН ОПЕДМЮГМЮВЕММШУ БШОНКМЪРЭ БЯЕЦН КХЬЭ ПНКЭ ╚ЛСЯНПЫХЙНБ╩ С ОЕПБШУ ЛМНЦНЙКЕРНВМШУ НПЦЮМХГЛНБ (ЙЮЙ, МЮОПХЛЕП, ЩРН ДЕКЮЧР АКСФДЮЧЫХЕ ЮЛЕАНЖХРШ С ЦСАНЙ), МН Б УНДЕ ЩБНКЧЖХХ УНПДНБШУ ГЮЙПЕОХКЮЯЭ ГЮ МХЛХ ЕЯРЕЯРБЕММШЛ НРАНПНЛ. б ХМРЕПЕЯЮУ ДЮКЭМЕИЬЕЦН ХГКНФЕМХЪ ЛЮРЕПХЮКЮ ДЮММНИ ЙМХЦХ МЮГНБЕЛ ЕЕ ПЕКХЙРНБНИ ХЛЛСММНИ ЯХЯРЕЛНИ.
еЫЕ ОН РЕЛЕ юПУЕИЯЙХЕ ОПЕДЙХ ХЛЛСММНИ ЯХЯРЕЛШ ВЕКНБЕЙЮ:
- мЮПСЬЕМХЪ ЯХЯРЕЛ, ТСМЙЖХНМЮКЭМН ЯНОПЪФЕММШУ Я ХЛЛСММНИ ЯХЯРЕЛНИ
- оЯХУНКНЦХВЕЯЙХЕ РПЕАНБЮМХЪ ОПНТЕЯЯХИ РХОЮ ╚ВЕКНБЕЙ-ВЕКНБЕЙ╩, ╚ВЕКНБЕЙ-ГМЮЙ╩, ╚ВЕКНБЕЙ √ УСДНФЕЯРБЕММШИ НАПЮГ╩, ╚ВЕКНБЕЙ-ОПХПНДЮ╩
- хЛЛСММЮЪ ЯХЯРЕЛЮ
- релю 3: нПЦЮМШ ХЛЛСММНИ ЯХЯРЕЛШ.
- 14.1. хЛЛСММЮЪ ЯХЯРЕЛЮ Х НОСУНКЕБШИ ПНЯР.
- цКЮБЮ 6. оЮРНКНЦХВЕЯЙЮЪ ТХГХНКНЦХЪ ХЛЛСММНИ ЯХЯРЕЛШ
- I. нПЦЮМШ ХЛЛСММНИ ЯХЯРЕЛШ.
- хЛЛСММЮЪ ЯХЯРЕЛЮ
- нАЫХЕ ГЮЙНМНЛЕПМНЯРХ МЮПСЬЕМХИ ХЛЛСММНИ ЯХЯРЕЛШ
- 1.3. лНКЕЙСКШ ХЛЛСММНИ ЯХЯРЕЛШ
- релю 4: нМРНЦЕМЕГ ХЛЛСММНИ ЯХЯРЕЛШ
- 1.2. йКЕРЙХ ХЛЛСММНИ ЯХЯРЕЛШ
- цСЛНПЮКЭМЮЪ Х ЙКЕРНВМЮЪ ХЛЛСММШЕ ЯХЯРЕЛШ.
- хЛЛСММЮЪ ЯХЯРЕЛЮ С ОЮЖХЕМРНБ Я ЖХППНГНЛ ОЕВЕМХ
- щЙЯОЕПХЛЕМРЮКЭМНЕ БНЯОПНХГБЕДЕМХЕ ЦХОНТСМЙЖХХ ХЛЛСММНИ ЯХЯРЕЛШ.
- нЯМНБМШЕ ОПЕДЯРЮБКЕМХЪ Н ЯРПНЕМХХ Х ТСМЙЖХНМХПНБЮМХХ ХЛЛСММНИ ЯХЯРЕЛШ
- 3.3. хЛЛСММЮЪ ЯХЯРЕЛЮ
- 3.1. нЯМНБМШЕ ЩРЮОШ ПЮГБХРХЪ ХЛЛСММНИ ЯХЯРЕЛШ ОКНДЮ
- 4.2.4. нЯМНБМШЕ ЮКЦНПХРЛШ МЮПСЬЕМХИ Б ХЛЛСММНИ ЯХЯРЕЛЕ ОПХ бхд.
- III. пЮГБХРХЕ ХЛЛСММНИ ЯХЯРЕЛШ