Исследование субстратной специфичности анти-ОБМаутоантител на модельных пептидах
Дальнейшее исследование специфичности аутоантител мы проводили с помощью сконструированной «эпитопной библиотеки» перекрывающихся пептидов в составе слитных белков с тиоредоксином, соответствующей полной последовательности ОБМ (Belogurov et al., 2008; Бачева и др., 2011).
Этот подход был выбран на основании концепции предпочтительного «представления» субстратных пептидов в растворе. Доступность соответствующего эпитопа в качестве потенциального субстрата аутоантител требует придания этому пептидному фрагменту определенной структуры в растворе, что может быть обеспечено лишь в составе биополимера. Для изучения протеолиза отдельных эпитопов ОБМ специфичными антителами были созданы двенадцать генноинженерных конструкций, содержащих последовательности, кодирующие различные фрагменты ОБМ (рис. 4), на основе экспрессионного вектора pET-32b-CH(+). Между тиоредоксином и последовательностью пептидов ОБМ был помещен линкер (SGGGG)3S. Подобный серин-глици- новый линкер используется в большом количестве работ для придания гибкости и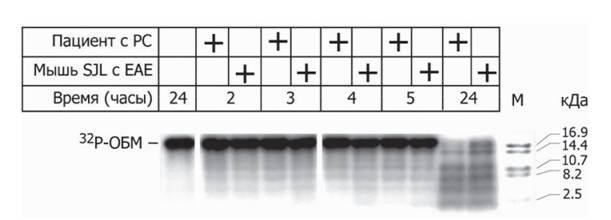
Рис. 3. Электрофореграмма продуктов гидролиза фосфорилированного 32P ОБМ специфичны- миаутоантителами, полученными от больных РС и модельных мышей линии SJLc индуцированным ЭАЭ, визуализированная с помощью авторадиографии
Представлены точки, соответствующие 2, 3, 4, 5 и 24 часам прохождения реакции. М - низкомолекулярный маркер. Адаптировано из (Ponomarenko et al., 2006)
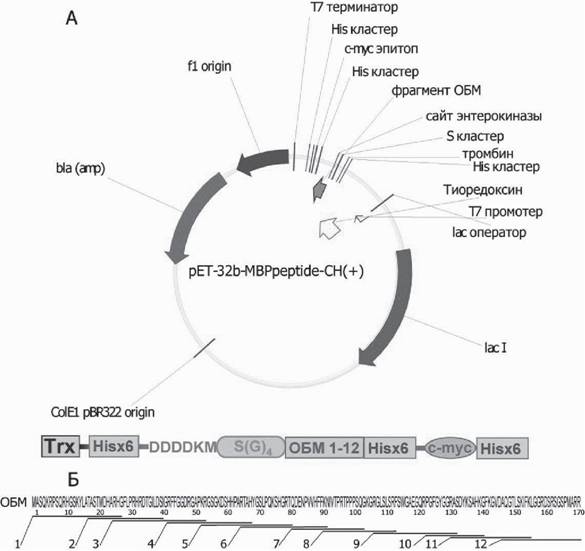
Рис. 4. А - схема плазмидного вектора, сконструированного для экспрессии фьюжн-белка тио- редоксина E. coli c фрагментами ОБМ. Б - аминокислотная последовательность ОБМ человека. Показано распределение пептидов 1-12 на последовательности ОБМ. Адаптировано из (Ponomarenko et al., 2006)
подвижности конструируемым рекомбинантным белкам.
Наличие подобного пептидного линкера содействует корректной презентации фрагментов ОБМ в составе фьюжн-белка с тиоредоксином.Для характеристики субстратной специфичности антител фьюжн-белки тиоре- доксина с фрагментами ОБМ инкубировали с иммуноглобулинами (суммарная IgG фракция), выделенными из сыворотки крови 26 больных РС, 22 пациентов с другими нейродегенеративными заболеваниями (other neurological diseases, OND), 11 здоровых доноров (healthy donors, HD) и мышей аутоиммунной линии SJL с ЭАЭ, индуцированным иммунизацией ОБМ (Belogurov et al., 2008). Как видно из приведенных электрофореграмм (рис. 5), в случае пациентов с РС и остеохондрозом (OND) протеолизу подвергались только пептиды, содержащие энцефалитогенную последовательность ОБМ. С другой стороны, у мышей SJL c ЭАЭ, предрасположенных к развитию аутоиммунных заболеваний, наблюдался множественный гидролиз перекрывающихся эпитопов ОБМ.
Каталитические свойства проявили антитела, изолированные из сывороток крови 77% пациентов с прогрессирующим и 85% с ремитирующим типом течения РС. Среди больных нейродегенеративными заболеваниями и здоровых доноров доля положительного каталитического ответа была 9% и 0% соответственно (рис 6). Исключение составили два случая остеохондроза, при которых было отмечено наличие каталитической деградации пептида ОБМ под действием аутоантител. Причем эти больные характеризовались значительной длительностью патологического процесса и серьезными нейрональными поражениями. Расщепление рекомбинантных белков бычьим трипсином, катепсином D и металлопротеазой матрикса 3 (MMP-3) (см. рис. 5, нижняя часть) проводили как контроль на специфичность гидролиза. Для всех исследованных антител методом масспектрометрии SELDI сайты гидролиза были локализованы исключительно в рекомбинантных фрагментах ОБМ (рис. 7, табл. 4), белок-носитель и линкерная часть оставались интактными. Причем, в случае абзимов, выделенных из сыворотки крови мышей линии SJL, сайты гидролиза рекомбинантных фрагментов повторяют таковые в нативном белке (см.
рис. 7). При детальном анализе продуктов расщепления энцефалитогенного пептида антителами было обнаружено, что сайтами гидролиза являются как Lys91 так и Arg97, причем лишь у двух пациентов степень расщепления по двум приведенным сайтам оказалась сравнимой (см. табл. 4). В большинстве случаев при РС наиболее предпочтительным для гидролиза являлся аргининовый сайт. Приведенные результаты свидетельствуют о том, что каталитические антитела, специфичные к участку ОБМ81-103, составляют подавляющее большинство в общем пуле ОБМ-гидролизу- ющих иммуноглобулинов при РС. Полученные данные находятся в соответствии с сообщениями относительно В-клеток, полученных от больных РС, узнающих преимущественно данный эпитоп, а также хорошо согласуются с фактом значительной иммунодоминантности этого фрагмента при представлении молекулами главного комплекса гистосовместимости II класса патогенных T-клеток (Ota et al., 1990; Hafler, 2004). В случае мышей линии SJL, предрасположенных к развитию аутоиммунных заболеваний, репертуар протеолитических IgG, специфичных к ОБМ, намного шире и включает в себя антитела практически на все исследован-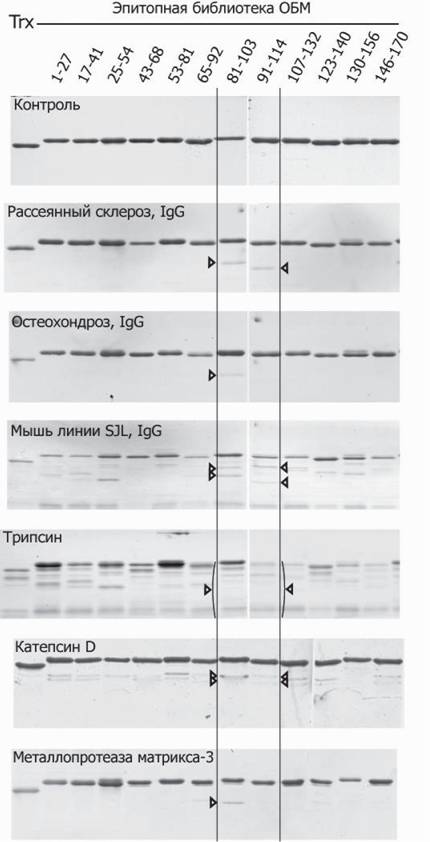
Рис. 5. Гидролиз эпитопной библиотеки аутоантителами и протеазами
Сверху вниз: контроль; гидролиз модельных пептидов протеолитическими антителами, полученными от пациента с РС, с остеохондрозом, аутоиммунных мышей линии SJL с ЭАЭ, индуцированным ОБМ; или же бычьим трипсином, катепсином D и металлопротеазой матрикса 3. Адаптировано из (Belogurov et al., 2008)
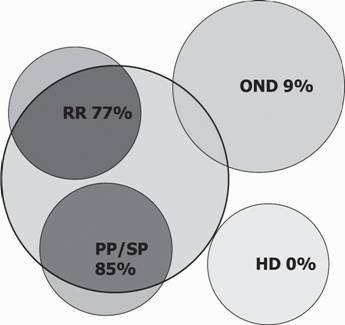
Рис. 6. Схематическое представление доли пациентов с различными формами РС (RR - ремитирующая, PP/SP - прогрессирующая), больных другими нейродегенеративными заболеваниями (OND), а также здоровых доноров, антитела которых проявляли каталитическую активность. Адаптировано из (Belogurov et al., 2008)

Рис.
7. Схема слитных белков, содержащих различные последовательности ОБМ человека Сайты гидролиза рекомбинантных фрагментов основного белка миелина специфичными иммуноглобулинами, изолированными из пациентов с рассеянным склерозом (обозначены серым) и мышей линии SJL с ЭАЭ (отмечены белым). Адаптировано из (Belogurov et al., 2008)ные пептиды (см. рис. 5). Сайты гидролиза рекомбинантных фрагментов данными антителами (см. рис. 7) в целом аналогичны действию на них неспецифической протеазы, однако по сравнению с ограниченным трипсинолизом не распространяются на тиоредоксин и линкерную часть. Объяснением возникновения такого разнообразия ОБМ-специфичных протеолитических антител у мышей линии SJL при иммунизации основным белком миелина может служить уже упоминавшаяся ранее особенность данной модельной линии - крайнее высокая частота индукции клонов В-клеток, продуцирующих каталитические антитела. Сравнение сайтов расщепления рекомбинантных субстратов из сконструированной нами библиоте-
Сайты гидролиза модельных пептидов аутоантителами, выделенными из сывороток крови пациентов с PC, и протеолитическими ферментами
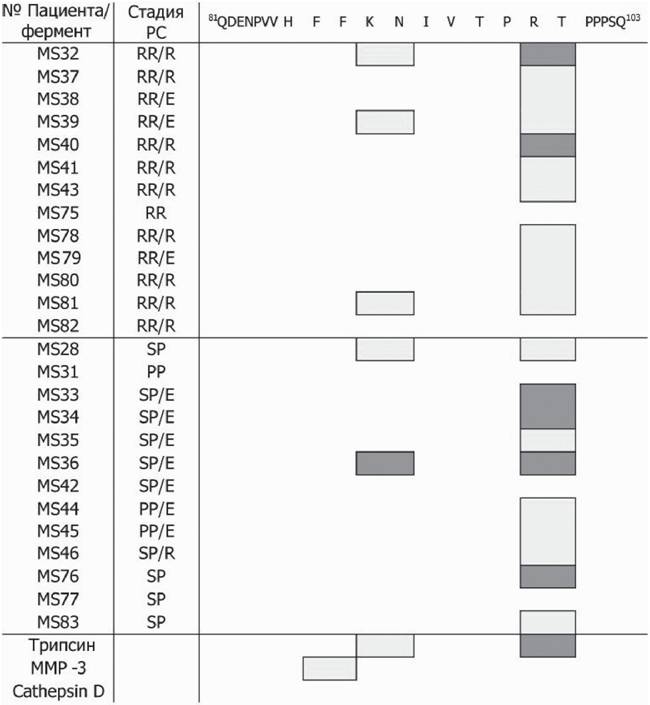
RR - ремиттирующий, PP - первично прогрессирующий, SP - вторично прогрессирующий тип РС; E - обострение, R - ремиссия, основные сайты отмечены светло-серым цветом, темно-серым показаны минорные.
ки под действием аутоантител и набора протеиназ, в том числе специфичных к ОБМ (см. рис. 5, 7, табл. 4), приводит к выводу о достаточно значительных отличиях в специфичности этих биокатализаторов (процессинг полноразмерного белка аналогичными биокатализаторами представлен в табл. 5). Этот факт позволил нам
Таблица 5
Сайты протеолиза основного белка миелина различными протеазами и каталитическими
антителами
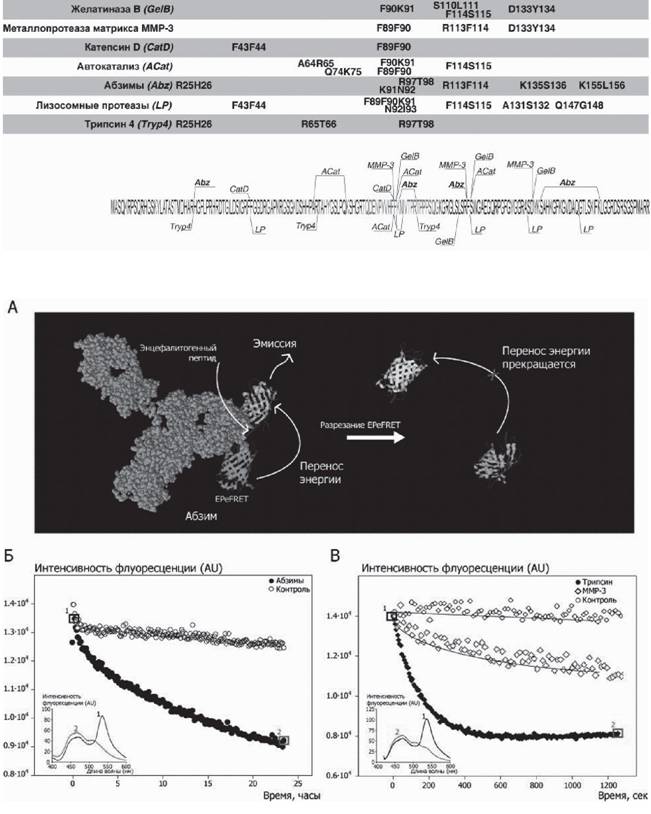
Рис. 8. А - принципиальная схема FRET-подхода к исследованию кинетики абзиматической реакции. Б, В - расщепление белка EPeFRET абзимами и модельными протеазами соответственно.
Во врезке показано изменение спектра флуоресценции во времени. Адаптировано из (Belogurov et al., 2008)
поставить вопрос о «специфическом модельном субстрате» для возможного скринингового анализа сывороток больных РС и гибридом, продуцирующих моноклональные каталитические антитела к ОБМ.
На основании данных по «дифференциальной специфичности» по сравнению с расщеплением протеиназами для реакции с аутоантителами нами был выбран фрагмент 81-103, содержащий энцефалитогенную последовательность ОБМ, для интеграции во FRET-пару Для этого на основе вектораpQe30 была создана генно-инженерная конструкция, экспрессирующая два флуоресцентных белка PS- CFP2 и TurboYF, соединенные вышеуказанным пептидом ОБМ, так называемый «EPeFRET»-белок (рис. 8А). При разрезании линкера прекращается резонансный перенос энергии и происходит падение флуоресценции (рис. 8Б,В).
6.2.
Еще по теме Исследование субстратной специфичности анти-ОБМаутоантител на модельных пептидах:
- Исследование стабильности фуллерена Сбо в биологических субстратах с использованием модельных систем in vitro
- Исследование фармакокинетики и фармакодинамики амидной формы нейропротекторного пептида HLDF-6 с использованием равномерно меченных изотопами водорода соединений
- Эксперимент по исследованию стабильности фуллерена Сбо в биологических субстратах с использованием модельных систем in vitro
- Исследование уровня концентрации антимикробных пептидов ротовой полости в зависимости от интенсивности кариозного процесса
- Исследование уровня концентрации антимикробных пептидов ротовой полости при заболеваниях пародонта
- Образование флуорофоров - продуктов взаимодействия свободного транс-ретиналя с аминогруппами белка и липидов в фоторецепторных клетках (исследование in vitro на модельных системах)
- ДИСПАНСЕРИЗАЦИЯ БОЛЬНЫХ ХРОНИЧЕСКИМИ ВИРУСНЫМИ ГЕПАТИТАМИ И НОСИТЕЛЕЙ НВsАg и анти – ВГС
- Использование специфичных тестов
- Глава 3. Исследование уровня окислительной модификации белков и молекул средней массы на модельной биологической системе желточных липопротеидов, продуктах пчеловодства каквеществах природного происхождения, обладающих антиоксидантным действием, и сыворотке крови экспериментальных животных (крысы)
- Что такое «заболевания психосоматической специфичности»?
- Факторы, определяющие чувствительность и специфичность
- Компромисс между чувствительностью и специфичностью
- Определение специфичности, чувствительности, положительного и отрицательного прогностического показателя (ПП)
- Специфичность изменений системы крови больных острой лучевой болезнью
- Структурная специфичность образной психонервной деятельности.
- Верификация, оценка чувствительности и специфичности модели классификации