Роль аутоантител в патогенезе РС
В норме В-клетки не способны преодолевать гематоэнцефалический барьер, однако при развивающимся воспалении его проницаемость может нарушаться, открывая доступ в ЦНС В-клеткам, иммуноглобулинам и белкам комплемента.В- клетки могут служить антиген-презентирующими для аутореактивных Т-клеток, привлекать их в ЦНС и производить костимуляцию (Sospedra, Martin, 2005).
Так, у больных наблюдается накопление в ЦНС клонально родственных В-клеток (Colombo et al., 2000) и повышенный синтез в ткани мозга и спинномозговой жидкости антител, специфичных к ОБМ, ПЛП и МОГ (Reindl et al., 1999), а также к ДНК и другим компонентам нервных тканей. Показано, что уровень аутоантител к ОБМ коррелирует с воспалительным процессом в ЦНС, согласно данным МРТ (Kuhle et al., 2007а). Анализ аутореактивных клонов В-клеток показал повышенную встречаемость семейств D2, D3, JH4 и зародышевых линий VH1-69, VH4-34 и VH4-39 (Baranzini et al., 1999).Превалирующим иммуногенным компонентом миелина для В-клеток человека является ОБМ, он же является наиболее сильным индуктором ЭАЭ у животных. При исследовании роли В-клеточного звена в патогенезе РС было показано, что у некоторых больных количество антител-продуцирующих В-лимфоцитов, специфичных к ОБМ, составляло в тканях мозга более 50% в общем пуле В-клеток (Gerritse et al., 1994). С другой стороны, IgG-опосредованный фагоцитоз миелинового дебриса макрофагами и активация комплемента внутри ЦНС являются косвенными доказательствами участия антител в патогенезе РС. Группой исследователей (Egg et al., 2001) было показано, что у 72% больных РС в сыворотке детектируются анти-МОГ антитела и в 59% случаев обнаруживаются аутоантитела к ОБМ. При этом среди иммуноглобулинов к названным антигенам доминировали изотипы IgM, а также IgG1 и IgG3. При скрининге пациентов с риском заболевания РС на присутствие аутоантител к МОГ и ОБМ было установлено, что 83% больных с положительным ответом на анти-МОГ и 95% на оба антитела имели первое обострение в течение 52 месяцев, в то время как 77% испытуемых, сыворотка крови которых не содержала ни одно из этих антител, не были затронуты болезнью.
Данное наблюдение, возможно, позволит диагностировать РС в его доклинической форме (Berger et al., 2003).Известно, что у мышей, дефицитных по В-системе иммунитета, возможно индуцировать ЭАЭ энцефалитогенным пептидом МОГ35-55, однако они резистентны к цельному белку (Lyons et al., 1999). Это наблюдение очередной раз подчеркивает факт о необходимости В-клеточной системы для корректного презентирования антигена и инициации значимого аутоиммунного заболевания. Линии В-клеток узнают различные эпитопы МВР, в первую очередь область от 84-го до 102-го аминокислотного остатка. К.Г. Уоренн и И. Кац (Warren, Catz, 1995) в своих работах неоднократно демонстрировали, что внутривенное введение больным РС синтетических пептидов, повторяющих иммунодоминантные эпитопы ОБМ (ОБМ75-95), приводит к значительному снижению титра анти-ОБМ антител как в сыворотке, так и в спинномозговой жидкости (Warren, Catz, 1995). Эти же исследователи в своих дальнейших публикациях (Warren, Catz, 1997) показали, что наиболее иммунодоминантным эпитопом ОБМ, с наибольшей эффективностью блокирующим аутоантитела к ОБМ, является последовательность аминокислот, расположенная между 85-м и 96-м пролинами, а именно: 85PVVHFFKNrVTP96.
Интересно, что, как уже упоминалось, подобный пептид можно использовать в качестве эффективного индуктора ЭАЭ у модельных животных. Однако только использование полноразмерной молекулы ОБМ и пептида МОГ 91-106 приводит к возникновению так называемого «epitope spreading», т.е. переключению иммунной системы на другие эпитопы и молекулы нейроантигенов (Bischof et al., 2004). Перечисленные сведения о деградации ОБМ свидетельствуют о важности этого процесса для презентации этого нейроантигена компонентам иммунной системы. В ранних работах лаборатории биокатализа ИБХ РАН было показано, что протеолиз этого белка может осуществляться аутоантителами (Ponomarenko et al., 2002), что нашло подтверждение в работах других авторов (Polosukhina et al., 2004).
Развитие этой работы состояло в исследовании специфичности антитело-зависимого связывания и процессинга ОБМ и его фрагментов, а также возможности применения этого процесса для диагностики РС.Активность аутоантител была нами охарактеризована по реакциям протеолиза с использованием неспецифических (биотинилированный бычий сывороточный альбумин (БСА), МОГ, тиоредоксин I E. coli) и специфического (ОБМ) субстратов.Ре- зультаты, представленные на рис. 1, свидетельствуют о том, что препараты антител, выделенных из сывороток больных РС и модельных мышей линии SJL с индуцированным ЭАЭ, обладали каталитической активностью только в отношении ОБМ.
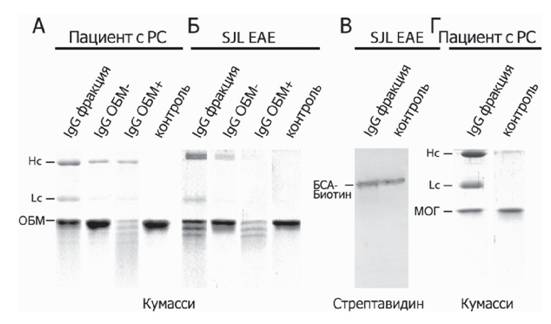
Рис. 1. Электрофореграмма (А, Б, Г) и иммуноблот (В) образцов гидролиза ОБМ (А), БСА (В) и МОГ (Г) препаратами антител, полученными из сывороток больных рассеянным склерозом (А, Г) и модельных мышей линии SJLc индуцированным ЭАЭ (экспериметальный аутоиммунный энцефаломиелит, EAE) (Б, В)
Показана протеолитическая активность суммарного пула иммуноглобулинов класса G (IgG фракция), а также сорбирующихся (IgG ОБМ+) и не сорбирующихся (IgG ОБМ-) на иммобилизованный ОБМ антител. Легкие и тяжелые цепи антител обозначены Lc и Hc соответственно. Адаптировано из (Ponomarenko et al., 2006)
Контрольные антитела из пулированной сыворотки здоровых доноров проявляли фоновый уровень протеолитической активности ко всем описанным субстратам.
Для определения сайт-специфичности гидролиза ОБМ аутоантителами было применено сочетание методов обращенно-фазовой хроматографии, трицинового белкового электрофореза и масспектрометрии SELDI. На первом этапе продукты гидролиза были разделены с использованием обращенно-фазовой хроматографии на колонке C4 (рис. 2А). Далее образцы фракций анализировали с использованием
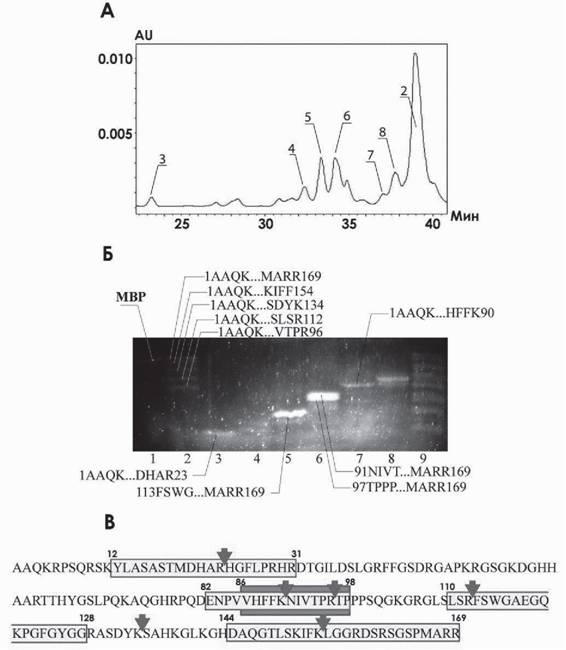
Рис. 2. Обращенно-фазовая хроматография продуктов гидролиза ОБМ аутоантителами (А) и анализ образцов посредством трицинового электрофореза (Б): 1 - полноразмерный ОБМ, 2-8 - фракции, полученные при разделении продуктов гидролиза на обращенной фазе (соответствующие им пики на (А) обозначены теми же номерами), 9 - гидролизат до разделения; стрелками показаны идентифицированные фрагменты.
В - основные сайты гидролиза ОБМ специфическимиаутоантителами (стрелки) Светло-серым отмечены иммунодоминатные районы белка, темно-серым обозначен энцефалитогенный пептид. Адаптировано из (Ponomarenko et al., 2006)трицинового электрофореза, с последующей детекцией низкомолекулярных пептидов при помощи флуоресцентного красителя Sypro Orange (рис. 2Б). Молекулярная масса преимущественных в количественном отношении и четко детектируемых на гидролизате до разделения (см. рис. 2Б, дорожка 9) продуктов была уточнена SELDI масспектрометрией. В результате объединения экспериментальных результатов и картирования молекулярных масс продуктов протеолиза относительно аминокислотной последовательности ОБМ удалось идентифицировать каждый фрагмент в структуре ОБМ (см. рис. 2Б, обозначены стрелками). В итоге было показано присутствие шести основных сайтов гидролиза ОБМ специфическими аутоантителами (рис. 2В).
При анализе характера расщепления ОБМ антителами (см. рис. 2) стало очевидным, что пять из шести сайтов гидролиза расположены в иммунодоминантных районах ОБМ, распознаваемых молекулами главного комплекса гистосовместимости II класса (HLA-DR2), вовлеченными в презентацию антигенов при РС (Hafler et al., 2005), а два локализованы на энцефалитогенном пептиде - индукторе ЭАЭ у модельных животных.
Для определения различий в скорости гидролиза ОБМ по отдельным сайтам было необходимо ввести специфическую метку с известной локализацией в молекулу ОБМ. Ранее in vitro было показано, что протеинкиназа С фосфорилирует бычий ОБМ преимущественно по остаткам Ser7, Ser54, и в малой степени Ser109, Ser114 и Ser150 (Kishimoto et al., 1985). Данный способ модификации белка был признан подходящим для анализа. Для детекции продуктов гидролиза 32Р-меченного in vitro ОБМ был применен трициновый электрофорез в ПААГ с последующей авторадиографией (рис. 3). Полученные результаты на качественном уровне демонстрируют, что гидролиз протекает независимо и с примерно равной скоростью по всем основным сайтам (Ponomarenko et al., 2006).
6.1.
Еще по теме Роль аутоантител в патогенезе РС:
- I.4. ИММУНОМОДУЛИРУЮЩАЯ РОЛЬ И ДИАГНОСТИЧЕСКАЯ ЦЕННОСТЬ ИССЛЕДОВАНИЯ РЕГУЛЯТОРНЫХ АУТОАНТИТЕЛ
- Роль микро-РНК в патогенезе болезни Паркинсона
- Роль в-амилоида в патогенезе БА
- Роль сфингомиелиназы в патогенезе болезни Альцгеймера
- Открытие генетической формы тромбофилии и АФС, и их роль в патогенезе ПЭ
- 1.2;2. Роль активации РААС в патогенезе АГ при ХОБЛ
- Роль эндокринных взаимоотношений в патогенезе эндокринопатий.
- Роль аутоиммунных процессов в патогенезе эндокринных нарушений.
- Роль нарушений липидного обмена в патогенезе атеросклероза
- Роль синдрома эндогенной интоксикации в патогенезе гестоза
- Роль ангиогенных факторов роста в патогенезе плацентарной недостаточности
- 1.2. Роль микроциркуляторных расстройств и дисфункции сосудистого эндотелия в патогенезе кардиометаболических заболеваний
- Роль предшественника амилоидного пептида и его метаболитов в патогенезе болезни Альцгеймера