Фармакологически вызванная хроническая гиперпролактинемия
Помимо наиболее широко используемой модели дегенерации катехоламинерги- ческих нейронов была разработана модель избирательной дегенерации только ДА- ергических нейронов при сохранении НА-ергических аксонов в аркуатном ядре у крыс.
Это делает возможным оценить вклад в развитие гиперопролактинемии каждой из катехоламинергических компонент - ДА-ергической и НА-ергической. Дегенерация только ДА-ергических нейронов тубероинфундибулярной системы моделируется путем внутрижелудочкового введения 6-ГДА после предварительного системного введения десметилимипрамина (ДМИ) - протектора НА-ергических нейронов. ДМИ связывается с мембранным переносчиком НА на НА-ергических нейронах и таким образом ингибирует обратный захват НА, предотвращая захват 6-ГДА и дегенерацию нейронов (Biegon, Rainbow, 1983; Fulceri et al., 2006; Sheward et al., 1985). В отличие от предыдущей модели с введением только 6-ГДА, на этой модели не происходит компенсаторного восстановления уровня ПРЛ в крови (табл. 4) (Дильмухаметова и др., 2009). В результате концентрация ПРЛ в крови сохраняется в 2 раза повышенной как на 14-й, так и на 45-й день после введения 6-ГДА с ДМИ (табл. 4) (Дильмухаметова и др., 2009). В гипофизе наблюдаются отчасти схожие с предыдущей моделью изменения: содержание ПРЛ снижено на 14-й день на 65%, а на 45-й день на 30%, вес гипофиза до 45-го дня отстает на 20% от веса гипофиза контрольных особей, а содержание мРНК Д2-рецепторов уменьшается на 30%, причем только на 45-й день (см. табл. 4) (Дильмухаметова и др., 2010). Кроме того, через 48 часов после введения 6-ГДА и ДМИ, так же как и на модели введения только 6-ГДА снижается, митотическая активность лактотрофов, а через 96 и 144 часа повышается (Lewinski et al., 1984).На данной модели изменения в уровне ПРЛ в крови коррелируют также с изменениями уровня ДА в гипоталамусе. В срединном возвышении и стебле гипофиза
Таблица 4
Изменение уровня пролактина (ПРЛ), мРНК Д^-рецепторов, дофамина (ДА), норадреналина (НА) и L-диоксифенилаланина (L-ДОФА) после введения ДМИ и 6-ГДА на 14-й и 45-й дни по сравнению с контролем и на 45-й день после введения ДМИ и 6-ГДА по сравнению с 14-м днем после введения ДМИ и 6-ГДА
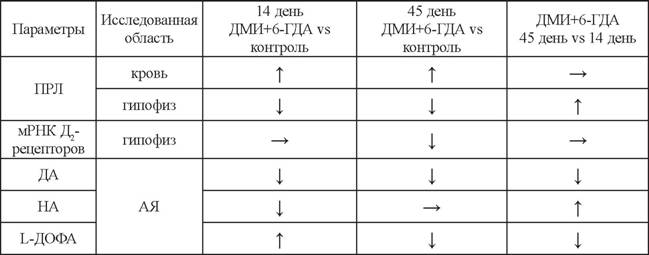
на 3-й день после введения 6-ГДА с ДМИ наблюдается снижение ДА на 50% без изменений содержания НА (Sheward et al., 1985).
В аркуатном ядре содержание ДА на 14-й день снижается на 75%, а на 45-й день - на 90%. При этом содержание НА незначительно (на 20%) снижается только на 14-й день и полностью восстанавливается к 45-му дню (см. табл. 4) (Дильмухаметова и др., 2009).Для оценки секреторной активности нейронов аркуатного ядра также применяли метод проточной инкубации слайсов ткани на 14-й и 45-й дни после введения нейротоксина и в контроле (Дильмухаметова и др., 2012, 2014). При введения ДМИ с 6-ГДА на 14-й день НА выделялся спонтанно и стимулированно, но уровень выделявшегося НА был ниже, чем в контроле (см. табл. 5). При этом НА-ергические нейроны отвечают на калиевую-стимуляцию (см. табл. 5). Кроме того, суммарный уровень НА при введении ДМИ с 6-ГДА почти не отличается от контрольного (см. табл. 5). Эти результаты отчасти согласуются с данными других авторов, продемонстрировавших, что концентрация НА в срединном возвышении не изменялась через 24 часа после внутривенного введения ДМИ и 6-ГДА (Sheward et al., 1985). Таким образом, наблюдается незначительное снижение НА-ергической иннервации с последующим частичным восстановлением.
В норме стимулированное выделение ДА нейронами аркуатного ядра в 3 раза выше, чем спонтанное. На 14-й день при введении ДМИ и 6-ГДА, так же как и при введении только 6-ГДА, было показано, что уровень спонтанного выделения ДА
Таблица 5
Изменение уровня дофамина (ДА), норадреналина (НА) и L-диоксифенилаланина (L-ДОФА) после введения ДМИ и 6-ГДА на 14-й и 45-й дни по сравнению с контролем и на 45-й день после введения ДМИ и 6-ГДА по сравнению с 14-м днем после введения ДМИ и 6-ГДА
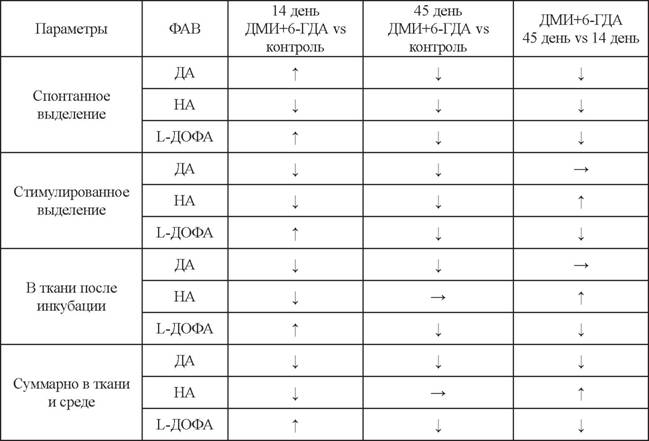
усиливался, а стимулированное выделение ДА отсутствовало, и в ткани уровень ДА был значительно снижен по сравнению с контролем (см. табл. 5) (Дильмухаметова и др., 2012). Также наблюдалось значительное снижение спонтанного и стимулированного выделения ДА по сравнению с контролем, но при этом ДА-ергическая система восстановила способность выделять ДА в ответ на калиевую стимуляцию, по сравнению с тем, что наблюдалось на 14-й день после введения токсина (см.
табл. 5). Таким образом, на 45-й день наблюдается восстановление функциональной активности ДА-продуцирующих нейронов. Однако уровень ДА в ткани после инкубации на 45-й день после введения только 6-ГДА был выше, а после введения ДМИ с 6-ГДА - значительно ниже, чем в контроле (Дильмухаметова и др., 2014). Это свидетельствует о том, что НА оказывает ингибирующее влияние на синтез ДА.На 45-й день после введения ДМИ с 6-ГДА уровень спонтанного и стимулированного выделения L-ДОФА значительно ниже, чем в контроле (см. табл. 5) (Дильмухаметова и др., 2014). Кроме того, суммарное содержание L-ДОФА в ткани и среде после введения ДМИ с 6-ГДА меньше, чем после введения только 6-ГДА (Дильмухаметова и др., 2014). Это свидетельствует о том, что уровень синтеза L-ДОФА ниже при сохранении НА-ергической иннервации аркуатного ядра, чем при ее отсутствии.
Биохимические изменения уровня ДА и НА в аркуатном ядре при введении ДМИ и 6-ГДА коррелируют с морфологическими изменениями в гипоталамусе после ведения нейротоксина. На модели введения ДМИ и 6-ГДА, так же как и на модели введения только 6-ГДА, наблюдалось снижение числа тирозингидроксила- за-иммунопозитивных нейронов в дорсомедиальной области аркуатного ядра, где локализованы в основном биферментные ДА-ергические нейроны, что объясняет значительное снижение синтеза ДА на обеих моделях как на 14-й день, так и на 45-й день (табл. 6) (Дильмухаметова и др., 2012, 2014). Однако в дорсомедиальной области оптическая плотность нейронов, коррелирующая с внутриклеточным содержанием тирозингидроксилазы, и площадь самих нейронов увеличивается при сохранении НА-ергического контроля (см. табл. 6). Это подтверждает предположение о протекторном влиянии НА по отношению к ДА-ергическим биферментным нейронам, которые локализованы в этой части аркуатного ядра.
При сохранении НА-ергической иннервации после введения ДМИ с 6-ГДА число тирозингидроксилаза-иммунопозитивных нейронов и содержание тирозингидрок- силазы в нейронах в вентролатеральной области не изменилось (Дильмухаметова и др., 2012, 2014) (см.
табл. 6). Приведенные данные свидетельствуют о том, что НА ингибирует синтез тирозингидроксилазы в моноферментных нейронах вентролатеральной области аркуатного ядра. Это хорошо согласуется с данными Дайкоку с соавторами (Daikoku et al., 1986), показавшими, что хирургическая деафферен- тация медиобазального гипоталамуса приводит к увеличению числа тирозингид- роксилаза-иммунопозитивных нейронов в вентролатеральной области аркуатного ядра. Прямое доказательство НА-ергического ингибиторного контроля экспрессии тирозингидроксилазы в неДА-ергических нейронах было недавно получено нами при изучении вазопрессинергических нейронов супраоптического ядра, эксперси- ирующих тирозингидроксилазу (Abramova et al., 2011).Изменение числа тирозингидроксилаза-иммунопозитивных (ТГ) нейронов и норадренергических (НА) терминалей аксоно, оптической плотности тирозингидроксилаза- иммунопозитивных нейронов в аркуатном ядре (АЯ) и в отдельных его областях - дорсомедиальной (ДМ) и вентролатеральной, после введения ДМИ и 6-ГДА на 14-й и 45-й дни по сравнению с контролем и на 45-й день после введения ДМИ и 6-ГДА по сравнению с 14-м днем после введения ДМИ и 6-ГДА
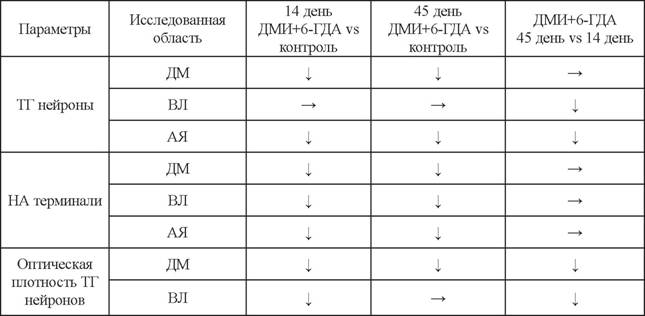
Из приведенных наших и литературных данных следует, что НА оказывает ингибирующее влияние на экспрессию тирозингидроксилазы в моноферментных нейронах вентролатеральной области аркуатного ядра и протекторное влияние на биферментные нейроны дорсомедиальной области аркуатного ядра.
Таким образом, при функциональной недостаточности ДА-ергических нейронов сохранение НА-ергической иннервации аркуатного ядра приводит к снижению экспрессии тирозингидроксилазы в неДА-ергических моноферментных нейронах аркуатного ядра и ингибированию синтеза ДА, что приводит к развитию хронической гиперпролактинемии.
2.2.
Еще по теме Фармакологически вызванная хроническая гиперпролактинемия:
- Фармакологически вызванная компенсируемая гиперпролактинемия
- Дефицит катехоламинов - компенсируемая и хроническая гиперпролактинемии
- Экспериментальные модели гиперпролактинемии
- Генетические модели гиперпролактинемии и пролактином
- Галоперидол - моделирование гиперпролактинемии без пролактиномы
- 4.2. Разработка способа закрытия трепанационного отверстия лицевой стенки гайморовой пазухи при хирургическом лечении хронического одонтогенного гайморита, вызванного выведением пломбировочного материала в верхнечелюстной синус (положительное решение о выдаче патента РФ по заявке №2008105099 (005542) от 20.02.2008).
- Моделирование гиперпролактинемии
- Фармакологические радиозащитные рецептуры
- Фармакологические препараты и НФР
- Фармакологическое действие глюкокортикоидов.
- Фармакологические аспекты хиральности
- 1.1. Фармакологическая характеристика и данные о влиянии барбитуратов на структуры ЦНС
- Роль статинов в фармакологической коррекции инсулинорезистентности при инфаркте миокарда
- Патогенетические подходы к фармакологической коррекции повреждений тонкой кишки и печени
- Молекулярные аспекты патогенеза и современные подходы к фармакологической коррекции болезни Альцгеймера
- Механизмы участия глутаматных рецепторов в патогенезе судорожных состояний и перспективы фармакологических воздействий
- 8.2. Вызванные потенциалы мозга