Фармакологически вызванная компенсируемая гиперпролактинемия
Гиперпролактинемию, обусловленную нарушением гипоталамического звена регуляции секреции ПРЛ, а именно дегенерацией ДА-ергических нейронов аркуат- ного ядра, чаще всего моделируют на крысах фармакологически с помощью 6-гид- роксидофамина (6-ГДА) - специфического нейротоксина катехоламинергических нейронов.
6-ГДА похож по структуре на ДА и НА, что обусловливает его специфический захват из внеклеточной среды в ДА-ергические и НА-ергические нейроны с помощью мембранного переносчика (Biegon, Rainbow, 1983; Blandini et al., 2008; Jonsson, 1983). В процессе действия 6-ГДА сначала подавляет синтез катехоламинов, а затем вызывает дегенерацию нейронов (Bartholini et al., 1970). Внутриклеточно токсин способствует разобщению окислительного фосфорилирования, что приводит к гибели нейронов. Традиционно 6-ГДА вводят стереотаксически в латеральные желудочки мозга (Ershov et al., 2005; Fenske, Wuttke, 1976; Зиязетдинова и др., 2008). 6-ГДА в процессе переноса с ликвором из боковых желудочков сначала в 3-й, а затем в 4-й желудочки диффундирует в близлежащие отделы мозга - последовательно в стриатум, медиобазальный гипоталамус и в черную субстанцию (Bosler, Calas, 1982). Кроме того, встречаются модели системного введения этого нейротоксина (Sheward et al., 1985). При этом 6-ГДА, который не проходит через гематоэнцефалический барьер (Blandini et al., 2008), оказывает токсическое действие на периферии и в циркумвентрикулярных областях мозга, в которых отсутствует гематоэнцефалический барьер, и одной из таких областей является срединное возвышение и вентарльная область аркуатного ядра (Sheward et al., 1985).При стереотаксическом введении под наркозом в латеральный желудочек мозга животного по координатам вводят канюлю для инъекции 6-ГДА в растворе или раствора для контроля (Ershov et al., 2005; Зиязетдинова и др., 2008). Особенностью этой модели является то, что первая волна дегенерации ДА-ергических нейронов сменяется компенсаторным увеличением ДА в аркуатном ядре гипоталамуса и восстановлением уровня ПРЛ в крови (Fenske, Wuttke, 1976; Зиязетдинова и др., 2008) (табл.
1). По данным М. Фенске и В. Вуттке (Fenske, Wuttke, 1976), концентрация ПРЛ в крови увеличивается уже через час после введения 6-ГДА, достигает максимума на 2-ой день, сохраняется повышенной до 15-го дня и восстанавливается после 37-го дня. В наших экспериментах на 14-й и 45-й дни после стереотаксических инъекций также наблюдается вначале повышение, а затем восстановление уровня ПРЛ в крови у крыс, что свидетельствует о компенсации гиперпролактинемии (см. табл. 1) (Зиязетдинова и др., 2008). В гипофизе также наблюдаются изменения после введения 6-ГДА: на 14-й и 45-й дни вес гипофиза снижается на 20% по сравнению с контролем, а содержание ПРЛ снижается на 30-35%. Содержание мРНК Д2-рецепторов в этом же эксперименте уменьшается на 30% на 14-й день и восстанавливается к 45-му дню (см. табл. 1) (Дильмухаметова и др., 2010). При этом в опыте митотическая активность уменьшается через 48 часов и увеличивается через 96 и 144 часа по сравнению с контролем, причем в самом контроле наблюдается постепенное снижение митотической активности в течение 48-144 часов для всех клеток переднего гипофиза (Lewinski et al., 1984).Выявленные изменения в уровне ПРЛ в крови у крыс на 14-й и 45-й дни после введения 6-ГДА хорошо коррелируют с изменениями уровня ДА в гипоталамусе. Так, по данным В.Дж. Шеварда с соавторами (Sheward et al., 1985), концентрация ДА и НА в срединном возвышении и стебле гипофиза на 3-й день после введения токсина снижается на 50%. По нашим данным, в аркуатном ядре после введения 6-ГДА на 14-й день содержание ДА снижается на 55%, а содержание НА - на 80%.
Изменение уровня пролактина (ПРЛ), мРНК Д^-рецепторов, дофамина (ДА), норадреналина (НА) и L-диоксифенилаланина (L-ДОФА) после введения 6-ГДА на 14-й и 45-й дни по сравнению с контролем и на 45-й день после введения 6-ГДА по сравнению с 14-м днем после введения 6-ГДА (Дильмухаметова и др., 2009, 2010)

На 45-й день уровень ДА в аркуатном ядре восстанавливается до контрольного уровня, а уровень НА по-прежнему снижен на 70% (см.
табл. 1) (Дильмухаметова и др., 2009; Зиязетдинова и др., 2008). Восстановление уровня ДА может быть связано с его кооперативным синтезом моноферментными недофаминергическими нейронами (Ugrnmov, 2008; Ugrumov et al., 2002, 2004, 2014) и/или с усилением его синтеза в сохранившихся ДА-ергических нейронах.Для оценки секреторной активности (синтез и выделение ДА) нейронов аркуат- ного ядра применяли метод проточной инкубации слайсов ткани на 14-й и 45-й дни после введения нейротоксина и в контроле (Дильмухаметова и др., 2012, 2014). В норме стимулированное выделение ДА нейронами аркуатного ядра в 3 раза выше, чем спонтанное. На 14-й день после введения 6-ГДА было показано, что уровень спонтанного выделения ДА усиливался, а стимулированное выделение ДА отсутствовало, причем в ткани в опыте уровень ДА был значительно ниже, чем в контроле (табл. 2) (Дильмухаметова и др., 2012). На 45-й день наблюдалось значительное снижение спонтанного и стимулированного выделения ДА по сравнению с контролем, но при этом ДА-ергическая система восстановила способность выделять ДА в ответ на калиевую стимуляцию (см. табл. 2) (Дильмухаметова и др., 2014). Таким образом, на 45-й день наблюдается восстановление функциональной активности ДА-продуцирующих нейронов. Однако уровень ДА в ткани после инкубации на 45-й день после введения только 6-ГДА был выше, чем в контроле (см. табл. 2). При этом на 14-й день после введения 6-ГДА наблюдалось спонтанное выделение НА без стимулированного усиления его выделения, и уровень НА в ткани был значительно снижен (см. табл. 2) (Дильмухаметова и др., 2012). Интересно отметить, что на 45-й день после введения 6-ГДА НА-ергические нейроны уже отвечают на калиевую стимуляцию (см. табл. 2) (Дильмухаметова и др., 2014).
Помимо ДА была изучена секреция L-ДОФА как конечного продукта синтеза в моноферментных нейронах, содержащих тирозингидроксилазу, и промежуточного продукта синтеза ДА в ДА-ергических нейронах. В норме в аркуатном ядре
Изменение уровня дофамина (ДА), норадреналина (НА) и L-диоксифенилаланина (L-ДОФА) после введения 6-ГДА на 14-й и 45-й дни по сравнению с контролем и на 45-й день после введения 6-ГДА по сравнению с 14-м днем после введения 6-ГДА
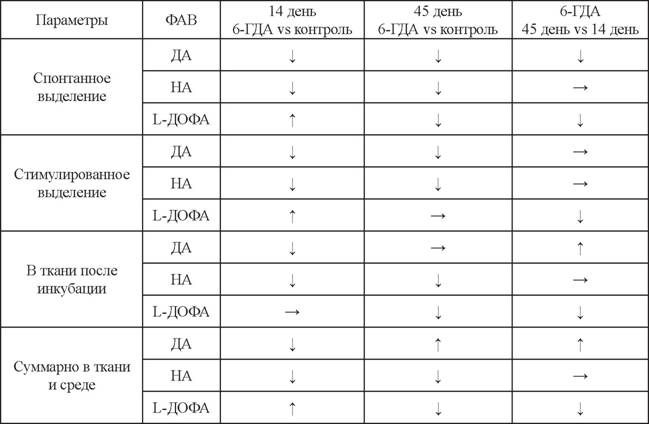
L-ДОФА выделяется только спонтанно (Дильмухаметова и др., 2012).
При этом на 45-й день после введения только 6-ГДА спонтанное выделение L-ДОФА ниже, чем в контроле, а после добавления калия в среду становиться равным контролю, демонстрируя небольшое симулированное усиление выделения (см. табл. 2) (Дильмухаметова и др., 2014). Это также сложно объяснить, так как L-ДОФА, как и ДА, синтезируются в цитоплазме, но при этом ДА после синтеза накапливается в везикулах, в то время как общепринято, что L-ДОФА содержится в цитозоле, а не в секреторных гранулах.Биохимические изменения уровня ДА и НА в аркуатном ядре тесно связаны с морфологическими изменениями в гипоталамусе после введения нейротоксина. Так, уже через 7 дней после введения 6-ГДА в аркуатном ядре наблюдается значительное снижение числа ДА-ергических нейронов, содержащих оба фермента синтеза ДА - тирозингидроксилазу и декарбоксилазу ароматических аминокислот, и увеличение числа недофаминергических моноферментных нейронов, содержащих по одному из ферментов синтеза ДА (Ershov et al., 2005). При введения только 6-ГДА на 14-й день наблюдалось значительное уменьшение числа НА-ергических аксонов (Дильмухаметова и др., 2012), а на 45-й день после введения только 6-ГДА в аркуатном ядре оставалось всего 35% терминалей НА-ергических аксонов (Дильмухаметова и др., 2014) (табл. 3). При этом на 14-й день после введения 6-ГДА
Изменение числа тирозингидроксилаза-иммунопозитивных (ТГ) нейронов и норадренергических (НА) терминалей аксонов, а также оптической плотности тирозингидроксилаза-иммунопозитивных нейронов в аркуатном ядре (АЯ) и в отдельных его областях - дорсомедиальной (ДМ) и вентролатеральной (ВЛ), после введения 6-ГДА на 14-й и 45-й дни по сравнению с контролем, а также на 45-й день после введения 6-ГДА по сравнению с 14-м днем после введения 6-ГДА
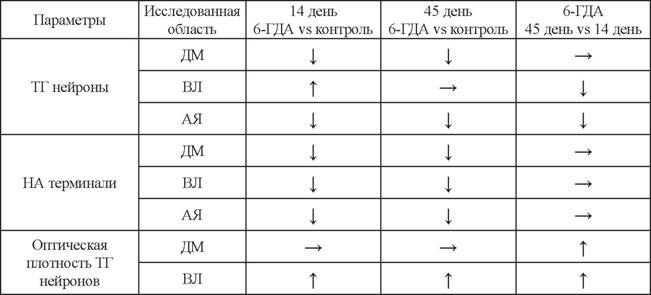
наблюдалось снижение числа тирозингидроксилаза-иммунопозитивных нейронов в дорсомедиальной области аркуатного ядра, где локализованы в основном биферментные ДА-ергические нейроны, что объясняет значительное снижение синтеза ДА как на 14-й, так и на 45-й день (Дильмухаметова и др., 2012, 2014) (см.
табл. 3). Однако в дорсомедиальной области оптическая плотность нейронов, коррелирующая с внутриклеточным содержанием тирозингидроксилазы, увеличивается при сохранении НА-ергического контроля (см. табл. 3). Это может свидетельствовать о протекторном влиянии НА на ДА-ергические биферментные нейроны, которые локализованы в этой области аркуатного ядра (Ershov et al., 2002а,Ь), что согласуется с данными о том, что НА способен снижать оксидативный стресс на культуре мезенцефальных ДА-ергических нейронов (Troadec et al., 2001). При этом на 45-й день после введения 6-ГДА при нарушении НА-ергической иннервации аркуатного ядра в вентролатеральной области аркуатного ядра, где содержатся преимущественно неДА-ергические нейроны, экспрессирующие только тирозингидроксилазу (Ershov et al., 2002а,Ь; Okamura et al., 1988) и участвующие в кооперативном синтезе ДА (Ugrumov et al., 2004), отмечено увеличение числа тирозингидроксила- за-иммунопозитивных нейронов и содержания самой тирозингидроксилазы в этих нейронах по сравнению с нормой (Дильмухаметова и др., 2012, 2014) (см. табл. 3).Таким образом, при фармакологическом выключении ДА-ергических нейронов и НА-ергических аксонов аркуатного ядра у крыс развивается компенсируемая гиперпролактинемия. При этом первоначальный дефицит ДА компенсируется за счет синтеза ДА неДА-ергическими нейронами аркуатного ядра (Fenske, Wuttke, 1976; Дильмухаметова и др., 2009, 2010; Зиязетдинова и др., 2008).
Еще по теме Фармакологически вызванная компенсируемая гиперпролактинемия:
- Фармакологически вызванная хроническая гиперпролактинемия
- Дефицит катехоламинов - компенсируемая и хроническая гиперпролактинемии
- Экспериментальные модели гиперпролактинемии
- Генетические модели гиперпролактинемии и пролактином
- Галоперидол - моделирование гиперпролактинемии без пролактиномы
- Моделирование гиперпролактинемии
- Фармакологические радиозащитные рецептуры
- Фармакологические препараты и НФР
- Фармакологическое действие глюкокортикоидов.
- Фармакологические аспекты хиральности
- 1.1. Фармакологическая характеристика и данные о влиянии барбитуратов на структуры ЦНС