цкюбю 10 ренперхвеяйхе юяоейрш щохделхнкнцхвеяйни нжемйх оюмделхх цпхоою A(H1N1)pdm09 2009-2010 ЦЦ.
цПХОО НЙЮГЮКЯЪ ЦНПЮГДН АНКЕЕ ЯКНФМНИ Х ЛМНЦНЦПЮММНИ ОПН╜АКЕЛНИ, ВЕЛ БЯЕ ХКХ, ОН ЙПЮИМЕИ ЛЕПЕ, АНКЭЬХМЯРБН ХГ ХГБЕЯРМШУ НЯРПШУ Х МЮХАНКЕЕ ПЮЯОПНЯРПЮМЕММШУ ГЮАНКЕБЮМХИ ВЕКНБЕЙЮ ХМ╜ТЕЙЖХНММНИ ОПХПНДШ.
цПХООНГМЮЪ ХМТЕЙЖХЪ НРКХВЮЕРЯЪ НР ОПЮ╜ЙРХВЕЯЙХ КЧАШУ ДПСЦХУ БХПСЯМШУ ХМТЕЙЖХИ ПЪДНЛ НЯНАЕММНЯРЕИ, ЙНРНПШЕ ДЕКЮЧР ЕЕ ОНХЯРХМЕ СМХЙЮКЭМНИ. оНЩРНЛС МЮ ЕЕ ОПХЛЕПЕ МЕЙНРНПШЕ ГЮЙНМНЛЕПМНЯРХ ЩОХДЕЛХНКНЦХХ ЦПХООЮ ЛШ ОНОШРЮКХЯЭ ХМРЕПОПЕРХПНБЮРЭ Я ОНГХЖХХ РЕУ РЕНПЕРХВЕЯЙХУ ОНКНФЕМХИ, ЙНРН╜ПШЕ ОПЕДЯРЮБКЕМШ Б ОПЕДШДСЫХУ ЦКЮБЮУ.б 2009-2010 ЦЦ. Б ЛХПЕ ЯКНФХКЮЯЭ НЯНАЕММЮЪ ЩОХДЕЛХНКН╜ЦХВЕЯЙЮЪ ЯХРСЮЖХЪ ОН ЦПХООС, НАСЯКНБКЕММЮЪ РЕЛ, ВРН МЮЯЕКЕМХЧ ОКЮМЕРШ НДМНБПЕЛЕММН СЦПНФЮКХ ЖХПЙСКХПСЧЫХЕ ЬРЮЛЛШ РЮЙ МЮГШБЮЕЛНЦН ╚ЯЕГНММНЦН╩ ЦПХООЮ A(H1N1), A(H3N2) Х б, БОЕПБШЕ БШЪБКЕММНЦН Б 2009 Ц. БХПСЯЮ ю/йЮКХТНПМХЪ/07/2009(мь17 ХКХ ЙЮЙ ЕЦН ЯЕИВЮЯ НАНГМЮВЮЧР - A(H1N1)pdm09, Х ОПХ ЩРНЛ НЯРЮБЮ╜КЮЯЭ ПЕЮКЭМНИ СЦПНГЮ ПЮЯОПНЯРПЮМЕМХЪ ОРХВЭЕЦН ЦПХООЮ Λ(H5N1) Х A(H7N7), НЯКНФМХБЬЕЦН ЩОХДЕЛХВЕЯЙСЧ НАЯРЮМНБЙС МЮ МЮЬЕИ ОКЮМЕРЕ Б ЙНМЖЕ ОПНЬКНЦН БЕЙЮ. яКЕДСЕР ГЮЛЕРХРЭ, ВРН ДН 1977 Ц., ЦНДЮ ОНЯКЕДМЕИ Б XX Б. ОЮМДЕЛХХ ЦПХООЮ, МЮЯЕКЕМХЧ СЦПНФЮК РНКЭ╜ЙН ЙЮЙНИ-МХАСДЭ НДХМ ЯЕПНРХО БХПСЯЮ ЦПХООЮ ю. нДМЮЙН ОНЯКЕ МЕЦН ЯХРСЮЖХЪ ОН МЕХГБЕЯРМШЛ ОПХВХМЮЛ ЙЮПДХМЮКЭМН ХГЛЕМХКЮЯЭ, Х Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ ЯРЮКХ НДМНБПЕЛЕММН ЖХПЙСКХПНБЮРЭ ЙЮЙ ПЮГМШЕ ЯЕПНРХОШ БХПСЯЮ ЦПХООЮ ю (H1N1 Х H3N2), РЮЙ Х ЦПХОО б.
бМНБЭ ТНПЛХПСЧЫХЕЯЪ ДПЕИТНБШЕ БЮПХЮМРШ БХПСЯНБ ОЕПЕ╜ЯРЮКХ БШРЕЯМЪРЭ ПЮМЕЕ ЖХПЙСКХПСЧЫХЕ [1, 2]. оПХ ЩРНЛ ЯРЮК ЯС╜ЫЕЯРБЕММН ХГЛЕМЪРЭЯЪ Х ЛЕУЮМХГЛ ПЮГБХРХЪ ЩОХДЕЛХИ ЦПХООЮ: ХГ ОПЕХЛСЫЕЯРБЕММН ЩЙГНЦЕММНЦН, Р.Е. НАСЯКНБКЕММНЦН ГЮМНЯНЛ БМНБЭ ЯТНПЛХПНБЮММШУ ЬРЮЛЛНБ (ДПЕИТНБШУ ХКХ ЬХТРНБШУ) ЩРНИ ХМ╜ТЕЙЖХХ Б МЮЬС ЯРПЮМС, ОПНХГНЬЕК ЯДБХЦ Б ОНКЭГС ОПЕНАКЮДЮМХЪ 209
ЩМДНЦЕММНЦН УЮПЮЙРЕПЮ ПЮЯОПНЯРПЮМЕМХЪ ЦПХООЮ [1, 3]. й ЯНФЮКЕ╜МХЧ, ЮБРНПШ МЕ ОНЪЯМЪЧР, ВРН НМХ ХЛЕЧР Б БХДС ОНД ЩМДНЦЕММШЛ ЛЕУЮМХГЛНЛ.
йЮЙ ЛШ ОНКЮЦЮЕЛ, РЮЙНЕ ПЮГБХРХЕ ЩОХДЕЛХИ БНГЛНФ╜МН РНКЭЙН БЯКЕДЯРБХЕ ДКХРЕКЭМНИ ОЕПЯХЯРЕМЖХХ БХПСЯЮ ЦПХООЮ (ЙНРНПЮЪ ОПХГМЮЕРЯЪ МЕ БЯЕЛХ ХЯЯКЕДНБЮРЕКЪЛХ) Х ОНЯКЕДСЧЫЕИ ЕЕ РПЮМЯТНПЛЮЖХХ Б ЛЮМХТЕЯРМСЧ ТНПЛС ОНД БКХЪМХЕЛ ТЮЙРНПНБ, БКХЪЧЫХУ МЮ ХЛЛСММШИ ЯРЮРСЯ ВЕКНБЕЙЮ (ЯЛ. ЦК. 4). мЮ РЮЙСЧ БНГ╜ЛНФМНЯРЭ ПЮГБХРХЪ ЩОХДЕЛХИ ЦПХООЮ СЙЮГШБЮК ЕЫЕ й. щМДПЧЯ [4] - НДХМ ХГ ОЕПБННРЙПШБЮРЕКЕИ БХПСЯЮ ЦПХООЮ ю.оН ЛМЕМХЧ б.д. аЕКЪЙНБЮ Я ЯНЮБР. [5], Б ЛЕУЮМХГЛЕ ТНПЛХПН╜БЮМХЪ ОЕПЯХЯРЕМЖХХ БХПСЯЮ ЦПХООЮ ю ЛНФМН БШДЕКХРЭ ЯКЕДСЧЫХЕ ТЮГШ: 1) ОЕПЕУНД Й ОЕПЯХЯРЕМЖХХ; 2) ЯРЮАХКХГЮЖХЧ ОЕПЯХЯРЕМЖХХ; 3) ПЕЮЙРХБЮЖХЧ БНГАСДХРЕКЪ Х 4) ОПНЦПЕЯЯХПСЧЫЕЕ ЯНБЕПЬЕМЯРБН╜БЮМХЕ ЯОНЯНАМНЯРХ Й ОПНДСЙРХБМНИ ХМТЕЙЖХХ (БШУНД ХГ ОЕПЯХЯРЕМ╜ЖХХ). нЙНМВЮРЕКЭМШИ БШУНД ОПНХЯУНДХР ОПХ ЮЙРХБЮЖХХ ОЮЯЯЮФЮ БХПСЯЮ МЮ МЕХЛЛСММНЛ ЙНККЕЙРХБЕ. рЮЙНЕ ПЮГБХРХЕ ЯНАШРХИ ОПЕД╜ОНКЮЦЮЕР БНГЛНФМНЯРЭ МЕГЮБХЯХЛНЦН ТНПЛХПНБЮМХЪ НВЮЦНБ ЦПХООЮ ю Я МНБНИ ЮМРХЦЕММНИ ПЮГМНБХДМНЯРЭЧ [6]. оПХ ЩРНЛ ЯСЫЕЯРБЕММН ХГЛЕМХКХЯЭ Х ДПСЦХЕ НЯМНБМШЕ УЮПЮЙРЕПХЯРХЙХ ЩОХДЕЛХИ ЦПХООЮ: НМХ ЯРЮКХ МЮВХМЮРЭЯЪ Б ЦНПНДЮУ, МЕ РХОХВМШУ ДКЪ ПЮМЕЕ БНГМХЙЮБ╜ЬХУ ЩОХДЕЛХИ; ЯМХГХКЮЯЭ ВЮЯРНРЮ МЮОПЪФЕММШУ ЩОХДЕЛХИ ЦПХООЮ Я МЮПЮЯРЮМХЕЛ ЮЙРХБМНЯРХ ОПНВХУ ПЕЯОХПЮРНПМШУ БХПСЯНБ; ЩОХДЕЛХХ ПЮЯОПНЯРПЮМЪКХЯЭ Б ЛХПЕ Х МЮЬЕИ ЯРПЮМЕ ГМЮВХРЕКЭМН ЛЕДКЕММЕЕ Х ЯРЮКХ МНЯХРЭ ТПЮЦЛЕМРЮПМШИ УЮПЮЙРЕП; ОНЯРЕОЕММН СБЕКХВХБЮКНЯЭ ВХЯКН ЦНПНДНБ, МЕ БНБКЕЙЮЕЛШУ Б ЩОХДЕЛХЧ; ЯМХГХКЮЯЭ ЮЙРХБМНЯРЭ ЩОХДЕЛХВЕЯЙНЦН ОПНЖЕЯЯЮ (що) Б ЦНПНДЮУ ЛЕЦЮОНКХЯЮУ; ЯКНФХКЮЯЭ ПЕЦПЕЯЯХПСЧЫЮЪ РЕМДЕМЖХЪ ЯЛЕПРМНЯРХ Х КЕРЮКЭМНЯРХ; Б НЯМНБМНЛ ОНПЮФЮКЯЪ ДЕРЯЙХИ ЙНМРХМЦЕМР; ВЮЫЕ ЯРЮК ОНДЙКЧВЮРЭЯЪ Й ЩОХДЕЛХ╜ВЕЯЙХЛ ЯНАШРХЪЛ БХПСЯ ЦПХООЮ б [1-3, 7, 8].
мЮ ЩРНЛ НЯМНБЮМХХ ч.ц. хБЮММХЙНБ Я ЯНЮБР. [1] ОПХЬКХ Й ГЮ╜ЙКЧВЕМХЧ Н БНГЛНФМНЯРХ ОНЯРЕОЕММНЦН ОПЕЙПЮЫЕМХЪ ЩОХДЕЛХИ ЦПХООЮ, БШГБЮММШУ БХПСЯНЛ ю, Х БЯЕ АНКЭЬЕИ ЮЙРХБХГЮЖХХ МЮ ЩРНЛ ТНМЕ БХПСЯЮ б. йПНЛЕ РНЦН, ЙЮГЮКНЯЭ АШ ОНДРБЕПФДЮКЮЯЭ ЦХОНРЕГЮ НА НЦПЮМХВЕММНЯРХ БНГЛНФМШУ БЮПХЮМРНБ БХПСЯЮ ЦПХООЮ ю, ЯОН╜ЯНАМШУ БШГШБЮРЭ ОЮМДЕЛХЧ, ХАН ЯКНФХБЬЕЕЯЪ ДН 1977 Ц. ЯРПНЦНЕ ВЕПЕДНБЮМХЕ ЖХЙКНБ, ЯБЪГЮММШУ Я МНБШЛХ ХКХ ЯСЫЕЯРБЕММН НАМНБ-
КЕММШЛХ ЯЕПНРХОЮЛХ БХПСЯЮЛХ ЦПХООЮ ю, АШКН МЮПСЬЕМН ОНБРНП╜МНИ ХМРПНДСЙЖХЕИ Б ОНОСКЪЖХЧ ЯРЮПНЦН БЮПХЮМРЮ - ю(мь1).
рЕЛ МЕ ЛЕМЕЕ ОНЯКЕДМЕЕ ДЕЯЪРХКЕРХЕ ОПЕДОЮМДЕЛХВЕЯЙНЦН ОЕПХНДЮ ОПНУНДХКН МЮ ТНМЕ НФХБКЕММШУ ДХЯЙСЯЯХИ ЙЮЙ Н ОЕПЯОЕЙ╜РХБЮУ ОНЪБКЕМХЪ МНБНЦН ОЮМДЕЛХВЕЯЙНЦН БХПСЯЮ ЦПХООЮ ю, РЮЙ Х Н БНГЛНФМШУ ЛЕУЮМХГЛЮУ ТНПЛХПНБЮМХЪ ЕЦН ЬХТРНБНЦН БЮПХЮМРЮ [9, 10]. б ВЮЯРМНЯРХ, ОНЪБКЕМХЕ Б ЙНМЖЕ XX Б. ГЮАНКЕБЮМХИ ВЕКНБЕ╜ЙЮ, БШГБЮММШУ БХПСЯНЛ ОРХВЭЕЦН ЦПХООЮ ю(ф^), ЯОНЯНАЯРБНБЮКН ЮЙРХБХГЮЖХХ ЛМЕМХИ Н БЕПНЪРМНЯРХ ОПНМХЙМНБЕМХЪ ЩРНЦН БХПСЯЮ Б ВЕКНБЕВЕЯЙСЧ ОНОСКЪЖХЧ Б ПЕГСКЭРЮРЕ ЦЕММНИ ЛСРЮЖХХ ХКХ БНГ╜МХЙМНБЕМХЪ ПЕЮЯЯНПРЮМРНБ БХПСЯНБ ЦПХООЮ ВЕКНБЕЙЮ Х ОРХЖ [9,11]. оПХ ЩРНЛ МЕ ХЯЙКЧВЮКНЯЭ, ВРН ПЕЮЯЯНПРЮЖХЪ БНГЛНФМЮ Б НПЦЮМХГЛЕ ЯБХМЭХ [12].
б РН ФЕ БПЕЛЪ ДПСЦХЕ ЮБРНПШ ЯВХРЮКХ АНКЕЕ БЕПНЪРМШЛ БНГ╜БПЮЫЕМХЕ Б ОНОСКЪЖХЧ ПЮМЕЕ ЖХПЙСКХПНБЮБЬЕЦН ЬРЮЛЛЮ ЦПХООЮ ю(м2╧), ХКХ ФЕ ПЕГЙНЕ ОНБШЬЕМХЕ БХПСКЕМРМНЯРХ (Б ПЕГСКЭРЮРЕ ЛСРЮЖХХ) МШМЕ ЖХПЙСКХПСЧЫХУ ЬРЮЛЛНБ БХПСЯЮ ЦПХООЮ ВЕКНБЕЙЮ ю(ьл) ХКХ ю(ч╧) [9-11]. ю.а. аЕКНБ Х о.х. нЦЮПЙНБ [9] ОН ЩРНЛС ОНБНДС ОХЯЮКХ: ╚...бЕПНЪРМН, НЙНМВЮРЕКЭМСЧ РНВЙС Б ЯОН╜ПЕ ОНЯРЮБХР ЯЮЛЮ ОЮМДЕЛХЪ, ЕЯКХ НМЮ БННАЫЕ ЙНЦДЮ-МХАСДЭ АСДЕР.╩ оНЩРНЛС ОНЪБКЕМХЕ Б ЛЮПРЕ 2009 Ц. Б лЕЙЯХЙЕ МНБНЦН ЮМРХЦЕММНЦН БЮПХЮМРЮ БХПСЯЮ ЦПХООЮ A/йЮКХТНПМХЪ/07/2009(H1N1), БШГБЮБЬЕ╜ЦН Б XXI Б. ОЕПБСЧ ОЮМДЕЛХЧ, ЙЮЙ ЛШ ОНКЮЦЮКХ, ОНГБНКХР МЮЛ МЮ НЯМНБЕ ХЛЕЧЫХУЯЪ Б КХРЕПЮРСПЕ ДЮММШУ Н ЕЦН ПЮЯОПНЯРПЮМЕМХХ [2, 13-20] Б ЯНОНЯРЮБКЕМХХ Я ЛЮРЕПХЮКЮЛХ Н ПЮМЕЕ БНГМХЙЮБЬХУ ОЮМДЕЛХЪУ Х АНКЭЬХМЯРБЕ ЩОХДЕЛХИ XX Б. [1, 21-25], Ю РЮЙФЕ Я ПЕ╜ГСКЭРЮРЮЛХ ЯНАЯРБЕММШУ МЮАКЧДЕМХИ ГЮ ГЮАНКЕБЮЕЛНЯРЭЧ Х ЖХПЙС╜КЪЖХЕИ БХПСЯНБ ЦПХООЮ Я 1982 Ц. ОН МЮЯРНЪЫЕЕ БПЕЛЪ Б оПХЛНПЯЙНЛ ЙПЮЕ ДЮРЭ АНКЕЕ НАЗЕЙРХБМСЧ ЩОХДЕЛХНКНЦХВЕЯЙСЧ НЖЕМЙС ПЮГ╜КХВМШЛ ОПНЦМНГЮЛ, ЦХОНРЕГЮЛ Н ТЮЙРНПЮУ Х ЛЕУЮМХГЛЮУ ПЮГБХРХЪ що ЩРНИ ХМТЕЙЖХХ.
оПНБЕДЕММШИ ЮМЮКХГ, БН-ОЕПБШУ, ОНЙЮГЮК, ВРН ОНДРБЕПДХК╜ЯЪ ОПНЦМНГ бнг Н БНГЛНФМНЯРХ ОНЪБКЕМХЪ МНБНЦН ОЮМДЕЛХВЕЯЙН╜ЦН БХПСЯЮ ЦПХООЮ Б КЧАНЕ БПЕЛЪ Х Б КЧАНИ РНВЙЕ ОКЮМЕРШ [11]. бРНПНИ ПЮГ ГЮ ОЕПХНД ЦКНАЮКЭМНЦН ЛНМХРНПХМЦЮ ГЮ БХПСЯНЛ Х ХЛЕ╜ЧЫХУЯЪ ДЮММШУ Н ОЮМДЕЛХХ 1918-1920 ЦЦ. [10, 25] ОЮМДЕЛХЪ ЦПХО╜ОЮ МЮВЮКЮЯЭ МЮ юЛЕПХЙЮМЯЙНЛ ЙНМРХМЕМРЕ, РНЦДЮ ЙЮЙ ОПЕДОНВРЕМХЕ 211
Б ЩРНЛ ОКЮМЕ НРДЮБЮКНЯЭ чЦН-бНЯРНВМНИ юГХХ [11, 26].
бЛЕЯРЕ Я РЕЛ ОНЙЮ, БХДХЛН, МЕКЭГЪ ЯВХРЮРЭ НЙНМВЮРЕКЭМН СЯРЮМНБКЕММШЛ, Б ЙЮЙНЛ ЦЕНЦПЮТХВЕЯЙНЛ ПЕЦХНМЕ Х С ЙЮЙНЦН АХНКНЦХВЕЯЙНЦН БХДЮ ОПНХГНЬКЮ НЙНМВЮРЕКЭМЮЪ ЦЕМЕРХВЕЯЙЮЪ ПЕЙНЛАХМЮЖХЪ [18]. бН- БРНПШУ, ОНДРБЕПДХКЮЯЭ Х БНГНАКЮДЮКЮ ГННЮМРПНОНМНГМЮЪ РЕНПХЪ ОПНХЯУНФДЕМХЪ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ, ХАН МНБШИ БХПСЯ НЙЮ╜ГЮКЯЪ РПНИМШЛ ПЕЮЯЯНПРЮМРНЛ БХПСЯНБ ЦПХООЮ ЯБХМЭХ, ОРХЖ Х ВЕ╜КНБЕЙЮ [13].бЮФМН НРЛЕРХРЭ, ВРН РПНИМШЕ ПЕЮЯЯНПРЮМРШ БХПСЯНБ ЦПХО╜ОЮ Λ(H3N2) Х ю(мь1) Б ПЮГМШУ РНВЙЮУ ОКЮМЕРШ МЮВЮКХ БШДЕКЪРЭ ЕЫЕ Б 90-У ЦНДЮУ ОПНЬКНЦН БЕЙЮ, МН ОЮМДЕЛХВЕЯЙНЦН ПЮЯОПНЯРПЮМЕ╜МХЪ НМХ МЕ ОНКСВХКХ [13]. б ЩРНИ ЯБЪГХ СЛЕЯРМН ОПХБЕЯРХ ЛМЕМХЕ й. щМДПЧЯ, ЯВХРЮБЬЕЦН, ВРН МНБШЕ ЬРЮЛЛШ БХПСЯЮ ЦПХООЮ ОНЪБ╜КЪЧРЯЪ ДНБНКЭМН ВЮЯРН, МН НМХ МЕ НАЪГЮРЕКЭМН БЕДСР Й ОЮМДЕЛХХ [4]. мЕ ЛЕМЕЕ ХГБЕЯРМШИ ЯОЕЖХЮКХЯР ОН ЦПХООС щ. йХКАСПМ РЮЙФЕ ЯВХРЮЕР, ВРН ЮМРХЦЕММЮЪ МНБХГМЮ ЯЮЛЮ ОН ЯЕАЕ МЕДНЯРЮРНВМЮ ДКЪ ЬХПНЙНЦН ПЮЯОПНЯРПЮМЕМХЪ БХПСЯЮ ЦПХООЮ [27].
б ЩРНЛ ОКЮМЕ ОПЕФДЕ БЯЕЦН МЕНАУНДХЛН ОПХМХЛЮРЭ БН БМХЛЮ╜МХЕ БНГЛНФМНЯРЭ ЩТТЕЙРХБМНИ ПЕЮКХГЮЖХХ ЛЕУЮМХГЛЮ ОЕПЕДЮВХ Б ЙНМЙПЕРМШУ СЯКНБХЪУ ЛЕЯРЮ Х БПЕЛЕМХ ХКХ МЕБШЯНЙСЧ ХМТЕЙЖХ- НММНЯРЭ ЬРЮЛЛНБ [11,13]. бЛЕЯРЕ Я РЕЛ Б ПСЯКЕ БНГГПЕМХИ ЮЙЮДЕ╜ЛХЙНБ б.л. фДЮМНБЮ [28] Х т.т. яНОПСМНБЮ [29], ПХЯЙМЕЛ ОПЕДОН╜КНФХРЭ, ВРН ТНПЛХПНБЮМХЕ Б НРДЕКЭМНЛ ЙНККЕЙРХБЕ ХКХ ПЕЦХНМЕ Б ЖЕКНЛ ЯОЕЖХТХВЕЯЙНЦН АХНЖЕМНГЮ, НЦПЮМХВХБЮЕР ОПНМХЙМНБЕМХЕ МЮ ЕЦН РЕППХРНПХЧ МНБШУ ОНРПЕАХРЕКЕИ АХНЩМЕПЦНПЕЯСПЯНБ ХГ-ГЮ МЕНАУНДХЛНЯРХ ╚ЩЙНМНЛМНЦН╩ ПЮЯУНДНБЮМХЪ ЛХЙПННПЦЮМХГЛЮЛХ ОНРЕМЖХЮКЮ ОНОСКЪЖХХ УНГЪЕБ. х РНКЭЙН ЙНЦДЮ ЯНГДЮЧРЯЪ НОПЕДЕ╜КЕММШЕ СЯКНБХЪ, ЯОНЯНАЯРБСЧЫХЕ ТНПЛХПНБЮМХЧ С ГЮМНЯХЛШУ ЬРЮЛЛНБ ЩЙНКНЦХВЕЯЙНЦН ОПЕХЛСЫЕЯРБЮ, ЩРН ОПХБНДХР Й ПЮЯ╜ОПНЯРПЮМЕМХЧ ЩОХДЕЛХВЕЯЙХ ГМЮВХЛШУ БЮПХЮМРНБ БНГАСДХРЕКЪ. бЮФМН ОНДВЕПЙМСРЭ, ВРН БХПСЯ A(H1N1/йЮКХТНПМХЪ) МЕЙНРНПНЕ БПЕЛЪ ЖХПЙСКХПНБЮК ЯПЕДХ ЯБХМЕИ Б лЕЙЯХЙЕ Х МЮ ЧЦЕ яью, МЕ БШГШБЮЪ ГЮАНКЕБЮМХИ МХ С ЩРХУ ФХБНРМШУ, МХ ХМТЕЙЖХХ С КЧДЕИ. нДМЮЙН Б ЛЮПРЕ 2009 Ц. МЮ ЯБХМНТЕПЛЕ МЕДЮКЕЙН НР Ц. лЕУХЙН БНГ╜МХЙКЮ ЩОХГННРХЪ ЦПХООЮ ЯБХМЕИ Х ПЮМЕЕ ЮБХПСКЕМРМШИ БХПСЯ ОН МЕОНМЪРМШЛ ОПХВХМЮЛ ОПЕБПЮРХКЯЪ Б БХПСКЕМРМШИ [13].
оПХ ЩРНЛ ОПНБНДХЛШЕ ХЯЯКЕДНБЮМХЪ ОНЙЮ МЕ БШЪБХКХ ЙЮЙХУ-КХАН ЛСРЮЖХНМ- 212
МШУ ХГЛЕМЕМХИ, ЙНРНПШЕ ЛНЦКХ АШ НАСЯКНБХРЭ ЯОНЯНАМНЯРЭ ЯБХ╜МНЦНБХПСЯЮ ЦПХООЮ ю(мь1/йЮКХТНПМХЪ) ХМТХЖХПНБЮРЭ КЧДЕИ Х ОЕПЕДЮБЮРЭЯЪ ЙНМРЮЙРМШЛ КХЖЮЛ [30].
мЮВЮБЬХЯЭ Б лЕЙЯХЙЕ, ЩОХДЕЛХЪ ОПНМХЙКЮ МЮ РЕППХРНПХЧ яью Х йЮМЮДШ, Ю ГЮРЕЛ БХПСЯ ПЮЯОПНЯРПЮМХКЯЪ Б ДПСЦХЕ ЯРПЮМШ Х ВЕПЕГ 3 ЛЕЯ. СФЕ АШК БШЪБКЕМ Б 137 ЦНЯСДЮПЯРБЮУ. оН ЯПЮБМЕМХЧ Я ОПНЬКШЛХ ОЮМДЕЛХЪЛХ ЕЦН ПЮЯОПНЯРПЮМЕМХЕ МЕЯЙНКЭЙН СЯЙНПХ╜КНЯЭ, ВРН НАЗЪЯМЪЕРЯЪ ЯНБПЕЛЕММШЛХ РЕМДЕМЖХЪЛХ ЦКНАЮКХГЮЖХХ що [12, 31]. оПХ ЩРНЛ Б чФМНЛ ОНКСЬЮПХХ, Ю Б ОНЯКЕДСЧЫЕЛ Х Б яЕБЕПМНЛ, МНБШИ БХПСЯ ЦПХООЮ БШРЕЯМХК (ГЮ МЕЙНРНПШЛ ХЯЙКЧ╜ВЕМХЕЛ) ХГ ЖХПЙСКЪЖХХ ЯЕГНММШЕ ЬРЮЛЛШ БХПСЯНБ ЦПХООЮ [2, 16].
юМЮКХГ ГЮАНКЕБЮЕЛНЯРХ ОЮМДЕЛХВЕЯЙХЛ ЦПХООНЛ Б ЛХПЕ Б 2009 Ц. ЯБХДЕРЕКЭЯРБНБЮК Н МЮКХВХХ ДБСУ БНКМ ЕЦН ПЮЯОПНЯРПЮМЕ╜МХЪ: ОЕПБЮЪ - ДН ЯЕПЕДХМШ ЮБЦСЯРЮ Я ОХЙНЛ Б ХЧКЕ Х БРНПЮЪ - Я НЙРЪАПЪ Х ОНЯКЕДСЧЫЕИ ПЕЦХЯРПЮЖХЕИ ГЮАНКЕБЬХУ Б ЪМБЮПЕ 2010 Х ДЮКЕЕ БОКНРЭ ДН КЕРМХУ ЛЕЯЪЖЕБ [2, 17]. цПСООНИ ПХЯЙЮ ГЮАНКЕБЮ╜МХЪ ЯРЮКХ КХЖЮ ЛНКНДНЦН Х ЯПЕДМЕЦН БНГПЮЯРЮ. аНКЕЕ РЪФЕКШЕ Х КЕ╜РЮКЭМШЕ ЯКСВЮХ ГЮАНКЕБЮМХЪ РЮЙФЕ НРЛЕВЮКХЯЭ С БГПНЯКШУ ЛНКНФЕ 50 КЕР, ОПХ ЩРНЛ КЕРЮКЭМШЕ ЯКСВЮХ ЯПЕДХ КХЖ ОНФХКНЦН БНГПЮЯРЮ ПЕЦХЯРПХПНБЮКХЯЭ ЯПЮБМХРЕКЭМН ПЕДЙН, РНЦДЮ ЙЮЙ ОПХ ЯЕГНММНЛ ЦПХООЕ, МЮОПНРХБ, НЙНКН 90 % РЪФЕКШУ Х КЕРЮКЭМШУ ХЯУНДНБ НРЛЕ╜ВЮЕРЯЪ ЯПЕДХ КХЖ Б БНГПЮЯРЕ 65 КЕР Х ЯРЮПЬЕ [2, 13, 18].
бЮФМН ОНДВЕПЙМСРЭ, ВРН ОНДНАМНЕ ПЮГБХРХЕ ЯНАШРХИ АШКН УЮПЮЙРЕПМН Б ОЮМДЕЛХЧ ╚ХЯОЮМЙХ╩ 1918-1919 ЦЦ. Х Б ОЮМДЕЛХХ 1957-1958, 1967-1968 ЦЦ. Х Б 1977 Ц. [1, 32]. б ЩРНИ ЯБЪГХ ЯКЕДС╜ЕР ГЮЛЕРХРЭ, ВРН ЛЕМЭЬХИ ПХЯЙ ГЮАНКЕБЮМХЪ КЧДЕИ ЯРЮПЬЕ 60 КЕР Б ОЮМДЕЛХЪУ ОПНЬКНЦН БЕЙЮ НАЗЪЯМЪКХ РЕЛ, ВРН НМХ ЛНЦКХ БЯРПЕ╜ВЮРЭЯЪ Я ЩРХЛ ХКХ АКХГЙХЛ ОН ЮМРХЦЕММНИ ЯОЕЖХТХВМНЯРХ БХПСЯНЛ ЦПХООЮ, ЙНРНПШИ, БНГЛНФМН, ЖХПЙСКХПНБЮК 60 Х АНКЕЕ КЕР МЮГЮД [1, 13]. оПЕДОНКЮЦЮЧР, ВРН Х Б ЩРС ОЮМДЕЛХЧ ОНРНЛС ПЕФЕ БНБКЕЙЮ╜КХЯЭ ОНФХКШЕ КЧДХ, ВРН НМХ ЛНЦКХ ЙНМРЮЙРХПНБЮРЭ Я АКХГЙХЛ ОН ЮМРХЦЕММНИ ЯРПСЙРСПЕ БХПСЯНЛ (A/H1N1) ДН 1957 Ц. [13]. нДМЮЙН ЩРНР ЯЕПНРХО БХПСЯЮ ЦПХООЮ ОПЮЙРХВЕЯЙХ Б МЕХГЛЕМЕММНЛ БЮПХЮМРЕ БМНБЭ ОНЪБХКЯЪ Б 1977 Ц.
[22, 33, 34] Х, ЯКЕДНБЮРЕКЭМН, Б ОНЯКЕДСЧ╜ЫЕЛ Я МХЛ БЯРПЕВЮКХЯЭ Х АНКЕЕ ЛНКНДШЕ ЦПЮФДЮМЕ. рЕЛ МЕ ЛЕМЕЕ, ЙЮЙ Х Б ОПНЬКШУ ОЮМДЕЛХЪУ, ЛНКНДШЕ КЧДХ Х БНБКЕЙЮКХЯЭ Б ЩРС ОЮМДЕЛХЧ ВЮЫЕ ОНФХКШУ Х ОЕПЕМНЯХКХ ГЮАНКЕБЮМХЕ Б АНКЕЕ РЪФЕ- 213КНИ ТНПЛЕ. оНЩРНЛС ч.г. цЕМДНМ [13] МЕ ХЯЙКЧВЮЕР БНГЛНФМНЯРЭ РНЦН, ВРН ОНЯКЕ 1977 Ц. ЛНЦ БНГМХЙМСРЭ ЮМРХЦЕММШИ БЮПХЮМР БХПСЯЮ A(H1N1), ЯУНДМШИ ОН ЮМРХЦЕММНИ ЯОЕЖХТХВМНЯРХ Я ЦЕЛЮЦКЧРХ- МХММНЛ ю(H1N1/йЮКХТНПМХЪ) Х КХЖЮ, Я МХЛ ЙНМРЮЙРХПНБЮБЬХЕ, ОПХНАПЕКХ ХЛЛСМХРЕР Х Й МНБНЛС ОЮМДЕЛХВЕЯЙНЛС БХПСЯС.
оПХ ЩРНЛ ЩОХДЕЛХНКНЦХВЕЯЙЮЪ ЯХРСЮЖХЪ Б пНЯЯХХ ПЮГБХБЮ╜КЮЯЭ ЯКЕДСЧЫХЛ НАПЮГНЛ. б РЕВЕМХЕ КЕРМЕЦН ОЕПХНДЮ ЯПЕДХ МЮ╜ЯЕКЕМХЪ ПЕЦХЯРПХПНБЮКХЯЭ ЕДХМХВМШЕ ГЮБНГМШЕ ЯКСВЮХ ЦПХООЮ. оПХВЕЛ, ГЮМНЯ ХМТЕЙЖХХ НЯСЫЕЯРБКЪКЯЪ ЙЮЙ ВЕПЕГ ГЮОЮДМШЕ, РЮЙ Х ЧФМШЕ, Х БНЯРНВМШЕ ЦПЮМХЖШ МЮЬЕИ ЯРПЮМШ [15] Б НРКХВХЕ НР ОПЕДШДСЫХУ ОЮМДЕЛХИ Х АНКЭЬХМЯРБЮ ЩОХДЕЛХИ ДН 1977 Ц., ЙНЦДЮ БХПСЯ ЦПХООЮ ОНОЮДЮК Б пНЯЯХЧ Б НЯМНБМНЛ РНКЭЙН ВЕПЕГ ЙЮЙНИ- КХАН ЯРПНЦН НОПЕДЕКЕММШИ ЦНПНД (лНЯЙБЮ, кЕМХМЦПЮД, нДЕЯЯЮ, уЮАЮПНБЯЙ) [1]. мЮДН ОНКЮЦЮРЭ, ВРН ЩРЮ НЯНАЕММНЯРЭ НАСЯКНБКЕМЮ ХГЛЕМХБЬХЛХЯЪ СЯКНБХЪЛХ ФХГМХ ПНЯЯХЪМ. бННАЫЕ ДКЪ АНКЭ╜ЬХМЯРБЮ ОПНЬКШУ ЩОХДЕЛХИ Х ОЮМДЕЛХИ ЦПХООЮ АШКН УЮПЮЙРЕПМН ДНБНКЭМН РХОХВМНЕ ПЮЯОПНЯРПЮМЕМХЕ. йЮЙ ОПЮБХКН, НМХ МЮВХМЮ╜КХЯЭ Б чЦН-бНЯРНВМНИ юГХХ, Ю ГЮРЕЛ ВЕПЕГ дЮКЭМХИ бНЯРНЙ ХКХ еБПНОЕИЯЙСЧ ВЮЯРЭ яяяп, гЮОЮДМСЧ еБПНОС Х яЕБЕПМСЧ юЛЕПХЙС ПЮЯОПНЯРПЮМЪКХЯЭ ОН ОКЮМЕРЕ [1]. хМРЕПЕЯМН, ВРН МХ ЩОХДЕЛХХ, МХ ОЮМДЕЛХХ ЦПХООЮ ПЮМЕЕ МХЙНЦДЮ Я бКЮДХБНЯРНЙЮ МЕ МЮВХМЮКХЯЭ, МЕЯЛНРПЪ МЮ РН, ВРН НМ ЦПЮМХВХР Я йХРЮЕЛ Х ХЛЕЕР НФХБКЕММШЕ ЛНПЯЙХЕ РПЮМЯОНПРМШЕ ЯБЪГХ ЯН ЯРПЮМЮЛХ ПЕЦХНМЮ чЦН-бНЯРНВМНИ юГХХ. нРЙПШРХЕ ЦНПНДЮ Б МЮВЮКЕ 90-У, ЯОНЯНАЯРБНБЮКН ЮЙРХБХГЮ╜ЖХХ РПЮМЯОНПРМШУ ЯБЪГЕИ Х ОПЕФДЕ БЯЕЦН ЮБХЮЯННАЫЕМХИ Я ПЮГ╜КХВМШЛХ ЯРПЮМЮЛХ Х ЯННРБЕРЯРБЕММН ПХЯЙ ГЮБНГЮ ЩРНИ ХМТЕЙЖХХ ЯСЫЕЯРБЕММН БНГПНЯ.
оЕПБШЕ ЯКСВЮХ БШДЕКЕМХЪ МНБНЦН ОЮМДЕЛХВЕЯЙНЦН БХПСЯЮ БН бКЮДХБНЯРНЙЕ АШКХ ГЮПЕЦХЯРПХПНБЮМШ СФЕ Б ХЧКЕ, ЙНЦДЮ НР ГЮПЮГХБЬЕЦНЯЪ Б яью ЦПЮФДЮМХМЮ АШКХ ХМТХЖХПНБЮМШ ЕЦН ДНВЭ Х ФЕМЮ[1]. бЮФМН ОНДВЕПЙМСРЭ, ВРН ГЮМНЯ ОЮМДЕЛХВЕЯЙНЦН БХПСЯЮ ЦПХООЮ МЕ БШГБЮК ЩОХДЕЛХХ Б ЦНПНДЕ, ЙЮЙ, БОПНВЕЛ, ЩРН ОПНХЯУН╜ДХКН Х Б ДПСЦХУ ПЕЦХНМЮУ пНЯЯХХ [2, 17]. хМРЕПЕЯМН, ВРН Б МЮЬЕИ ЯРПЮМЕ Б КЕРМХЕ ЛЕЯЪЖШ МХ ОЮМДЕЛХХ, МХ ЩОХДЕЛХХ ЦПХООЮ, БХДХ-
ЛН, МХЙНЦДЮ МЕ МЮВХМЮКХЯЭ, УНРЪ, Б ОПХМЖХОЕ, КЕРМХИ ОЕПХНД ДКЪ ЯРПЮМ яЕБЕПМНЦН ОНКСЬЮПХЪ МЕ ЪБКЪЕРЯЪ ХМЦХАХПСЧЫХЛ ТЮЙРНПНЛ ПЮГБХРХЪ ЩОХДЕЛХИ. б КХРЕПЮРСПЕ ХЛЕЕРЯЪ СОНЛХМЮМХЕ Н РНЛ, ВРН Б 1918 Ц. БХПСЯ ╚ХЯОЮМЙХ╩ АШК ГЮБЕГЕМ ХГ аПХРЮМХХ ХМРЕПБЕМРЮЛХ Б лСПЛЮМЯЙ Б ХЧМЕ, НРЙСДЮ АШЯРПН ПЮЯОПНЯРПЮМХКЯЪ ОН ЕБПНОЕИЯЙНИ ВЮЯРХ ЯРПЮМШ [25]. нДМЮЙН ЯКЕДСЕР ОНЛМХРЭ, ЙЮЙНЕ ЩРН АШКН БПЕЛЪ Б пНЯЯХХ. ьКЮ цПЮФДЮМЯЙЮЪ БНИМЮ, Х БПЪД КХ Б ЩРНР ОЕПХНД ОПН╜БНДХКНЯЭ МЮДКЕФЮЫЕЕ ЯКЕФЕМХЕ ГЮ ЩРНИ ХМТЕЙЖХЕИ. б ОЮМДЕЛХЧ 1957 Ц. ЯЕПНРХО БХПСЯ ЦПХООЮ ю(м2╧) АШК ГЮМЕЯЕМ МЮ РЕППХРНПХЧ яяяп Б ХЧМЕ [22], МН ЕЦН ЖХПЙСКЪЖХЪ НЦПЮМХВХКЮЯЭ МЕАНКЭЬХЛХ БЯОШЬЙЮЛХ Б КЕРМХУ ДЕРЯЙХУ СВПЕФДЕМХЪУ (ДЕРЯЙХЕ КЮЦЕПЪ, КЕР╜МХЕ ДЮВХ). йЯРЮРХ, ЩРНР БЮПХЮМР БХПСЯЮ БОЕПБШЕ ОНЪБХКЯЪ Б йХРЮЕ КЕРНЛ 1956 Ц., НДМЮЙН РНФЕ МЕ ОНКСВХК ЬХПНЙНЦН ПЮГБХРХЪ Б ЯРПЮМЕ, УНРЪ БЕЯМНИ 1957 Ц. НМ ЯРПЕЛХРЕКЭМН ПЮЯОПНЯРПЮМХКЯЪ МЕ РНКЭЙН Б йХРЮЕ, МН Х Б РПНОХВЕЯЙХУ ЯРПЮМЮУ [25]. рЮЙЮЪ НЯНАЕММНЯРЭ, МЮДН ОНКЮЦЮРЭ, НРМЧДЭ МЕ ЯБЪГЮМЮ, ЙЮЙ ЯВХРЮЧР МЕЙНРНПШЕ ЮБРНПШ, Я БКХ╜ЪМХЕЛ ЙКХЛЮРХВЕЯЙХУ ТЮЙРНПНБ МЮ що ЦПХООЮ [35].б ЩРНЛ ОКЮМЕ ЯКЕДСЕР ГЮЛЕРХРЭ, ВРН Б еБПНОЕ ЛЮЙЯХЛСЛ ГЮАНКЕБЮЕЛНЯРХ ЦПХООНЛ A(H1N1/йЮКХТНПМХЪ) ГЮПЕЦХЯРПХПНБЮМ Б ХЧКЕ [2], ДЮ Х ЩЙЯОКНГХБ╜МШИ ЕЕ ОНДЗЕЛ Б пНЯЯХХ ОПХЬЕКЯЪ МЮ НЙРЪАПЭ - ОЕПХНД, ЙНЦДЮ БН ЛМНЦХУ ПЕЦХНМЮУ ЯРПЮМШ ЕЫЕ ЯНУПЮМЪЧРЯЪ БШЯНЙХЕ РЕЛОЕПЮРСПШ. й РНЛС ФЕ Б РПНОХЙЮУ БХПСЯ ЖХПЙСКХПСЕР ЙПСЦКНЦНДХВМН [23].
мЕЯЛНРПЪ МЮ СЯХКЕММНЕ ЛЕДХЖХМЯЙНЕ МЮАКЧДЕМХЕ, Б ЯЕМРЪАПЕ Б оПХЛНПЯЙНЛ ЙПЮЕ, ЙЮЙ Х Б ЖЕКНЛ ОН пНЯЯХХ, ГЮАНКЕБЮМХЪ ЦПХООНЛ МЕ ПЕЦХЯРПХПНБЮКХЯЭ Х БХПСЯ МЕ БШДЕКЪКХ БОКНРЭ ДН ЙНМЖЮ ЯЕМРЪ╜АПЪ - МЮВЮКЮ НЙРЪАПЪ. б ЯБНЕ БПЕЛЪ й. щМДПЧЯ БШЯЙЮГЮК ОПЕДОНКН╜ФЕМХЕ, ВРН БХПСЯШ ЦПХООЮ ЛНЦСР ОЕПЕДЮБЮРЭЯЪ НР НДМНЦН УНГЪХМЮ Й ДПСЦНЛС, МЮУНДЪЯЭ МЮ МХГЙНЛ СПНБМЕ ЮЙРХБМНЯРХ; ОПХ ЩРНЛ НМХ КХЬЭ НАЕЯОЕВХБЮЧР ЯЕАЕ КНЙЮКЭМСЧ АЮГС МЮ ЯКХГХЯРШУ, МЕ БШГШ╜БЮЪ АНКЕГМХ, МН Х МЕ ЯНГДЮБЮЪ НАЫЕЦН ХЛЛСМХРЕРЮ [4]. б ЩРНЛ ОКЮ╜МЕ БЮФМШЕ ЯБЕДЕМХЪ ОПХБНДЪР ЯОЕЖХЮКХЯРШ ХГ МЕЙНРНПШУ НАКЮЯРЕИ дбтн. б ВЮЯРМНЯРХ, Б ЯШБНПНРЙЮУ ЙПНБХ ФХРЕКЕИ юЛСПЯЙНИ НА╜КЮЯРХ Х уЮАЮПНБЯЙНЦН ЙПЮЪ ДН МЮВЮКЮ ЩОХДЕЛХХ 2009 Ц. Б ОЕПБНЛ ЯКСВЮЕ Б 6 %, Ю БН БРНПНЛ Б 4 % АШКХ БШЪБКЕМШ ЮМРХРЕКЮ Й МНБНЛС БХПСЯС [19, 20]. рЕЛ ЯЮЛШЛ ОНДРБЕПФДЮЕРЯЪ БНГЛНФМНЯРЭ ЯЙПШРНИ ЖХПЙСКЪЖХХ БХПСЯЮ ЦПХООЮ. лНФМН ДСЛЮРЭ, ВРН ОНЯКЕ ДНЯРХФЕМХЪ МЕЙНЕЦН ЙПХРХВЕЯЙНЦН СПНБМЪ ЯПЕДХ МЮЯЕКЕМХЪ МЮВХМЮЧР ДНЛХМХ- 215
ПНБЮРЭ ЛЮМХТЕЯРМШЕ ТНПЛШ ХМТЕЙЖХХ Х ЩРН МЮУНДХР НРПЮФЕМХЕ Б ПНЯРЕ ГЮАНКЕБЮЕЛНЯРХ.
цКЮБМШЕ ЯНАШРХЪ БН бКЮДХБНЯРНЙЕ, ЙЮЙ Х Б ЖЕКНЛ ОН ЯРПЮМЕ, ЯРЮКХ ПЮГБНПЮВХБЮРЭЯЪ Б ЙНМЖЕ ЯЕМРЪАПЪ Х НЙРЪАПЕ, ЙНЦДЮ ОЕПБНМЮ╜ВЮКЭМН ЦПХОО A(H1N1)pdm09 АШК ГЮПЕЦХЯРПХПНБЮМ С 3 ЦПЮФДЮМ, БНГБПЮРХБЬХУЯЪ ХГ йХРЮЪ Х АШБЬХУ Б НДМНИ РСПХЯРХВЕЯЙНИ ЦПСО╜ОЕ, Ю ГЮРЕЛ, ОНЯКЕ НРДЕКЭМШУ ЯОНПЮДХВЕЯЙХУ ЯКСВЮЕБ, БЯОШЬЙХ ХМТЕЙЖХХ ЯРЮКХ БНГМХЙЮРЭ Б ЙПСОМШУ СВЕАМШУ СВПЕФДЕМХЪУ ЦН╜ПНДЮ: бцлс, бцсщя, дбцрс, ЬЙНКЮУ Х ОПХ ЩРНЛ СФЕ ЛНФМН АШКН ОПНЯКЕДХРЭ ЩОХДЕЛХВЕЯЙХЕ ЖЕОНВЙХ ЛЕФДС НРДЕКЭМШЛХ ЯКСВЮЪЛХ ГЮАНКЕБЮМХИ. лЮЙЯХЛСЛ ГЮАНКЕБЮЕЛНЯРХ, ЙЮЙ Х Б ЖЕКНЛ ОН пНЯЯХХ, ОПХЬЕКЯЪ МЮ НЙРЪАПЭ-ДЕЙЮАПЭ. оНЯКЕДМХЕ 5 ЯКСВЮЕБ ГЮПЕЦХЯРПХ╜ПНБЮМШ Б МЮВЮКЕ ЪМБЮПЪ 2010 Ц. Х ОНЯКЕ ЩРНЦН МЮВЮКЯЪ ОНДЗЕЛ ГЮ╜АНКЕБЮЕЛНЯРХ ЦПХООНЛ, БШГБЮММШИ БХПСЯНЛ б. мЕНАУНДХЛН ОНД╜ВЕПЙМСРЭ, ВРН МНБШИ ЩОХДЕЛХВЕЯЙХИ ЯЕГНМ 2010-2011 ЦЦ. УЮПЮЙРЕ╜ПХГНБЮКЯЪ РЕЛ, ВРН БХПСЯ A(H1N1)pdm09 БН бКЮДХБНЯРНЙЕ МЮВЮКХ БШДЕКЪРЭ РНКЭЙН Я ДЕЙЮАПЪ 2010 Ц. оПХВЕЛ Б НРКХВХЕ НР 2009 Ц. НМ ЖХПЙСКХПНБЮК НДМНБПЕЛЕММН Я БХПСЯЮЛХ A(H3N2) Х б, Р.Е. ОЕПЕ╜ЯРЮК ХУ БШРЕЯМЪРЭ, Ю ГЮМЪК ЛЕЯРН Б ЩРХНКНЦХВЕЯЙНИ ЯРПСЙРСПЕ ЯЕ╜ГНММНЦН ЦПХООЮ (РЮАК. 10.1).
рЮАКХЖЮ 10.1
оНМЕДЕКЭМЮЪ ДХМЮЛХЙЮ БШДЕКЕМХЪ БХПСЯНБ ЦПХООЮ НР ФХРЕКЕИ бКЮДХБНЯРНЙЮ
| мЕДЕКЪ ЦНДЮ | бЯЕЦН НАЯКЕДНБЮМН | вХЯКН БШДЕКЕММШУ ЬРЮЛЛНБ | |||
| Λ(H1N1) | A(H3N2) | A(H1N1) pdm09 | B | ||
| 1 | 2 | 3 | 4 | 5 | 6 |
2009 Ц.
| 4 | 33 | 8 | |||
| 5 | 28 | 14 | |||
| 6 | 30 | 13 | |||
| 7 | 57 | 17 | 1 | ||
| 8 | 45 | 23 | |||
| 9 | 81 | 22 | |||
| 10 | 63 | 16 | |||
| 11 | 81 | 15 | 1 | ||
| 12 | 112 | 34 | 4 | ||
| 13 | 74 | 19 | |||
| 14 | 70 | 3 | |||
| 15 | 79 | 10 | 1 | 10 | |
| 16 | 42 | 3 | 1 | 3 | |
| 17 | 20 | 1 | |||
| 18 | 17 | 1 | |||
| 19 | 24 | 1 | |||
| 20 | 19 | 1 | |||
| 21 | 9 | 1 |
оПНДНКФЕМХЕ РЮАК. 10.1
| 2 | 3 | 4 | 5 | 6 | |
| 25 | 14 | 1 | |||
| 28 | 11 | 4 | |||
| 33 | 35 | 1 | |||
| 36 | 22 | 1 | |||
| 40 | 85 | 1 | |||
| 42 | 62 | 1 | |||
| 43 | 92 | 9 | |||
| 44 | 143 | 18 | |||
| 45 | 161 | 20 | |||
| 46 | 172 | 10 | |||
| 47 | 454 | 54 | |||
| 48 | 4 | 2 | |||
| 50 | 178 | 25 | |||
| 51 | 190 | 33 | |||
| 52 | 75 | 3 | |||
| 53 | 60 | 13 | |||
| хРНЦН | 2642 | 201 | 10 | 188 | 16 |
2010 Ц.
| 4 6 7 8 9 10 11 12 13 14 15 16 17 19 20 22 26 43 44 47 49 50 51 52 53 | 71 30 33 35 36 52 22 33 46 41 23 15 9 11 10 6 5 12 8 8 11 12 10 11 14 17 | 1 3 1 4 1 | 2 4 1 2 2 | 2 1 1 5 1 7 2 9 14 6 8 1 7 1 1 2 2 2 1 1 1 | |
| хРНЦН | 581 | 10 | 5 | 76 |
2011 Ц.
| 4 | 22 | 5 | 3 | ||
| 5 | 23 | 4 | 6 | ||
| 6 | 24 | 8 | 2 | 4 | |
| 7 | 43 | 3 | 1 | 3 | |
| 8 | 71 | 3 | 2 | 7 | |
| 9 | 167 | 3 | 10 | ||
| 10 | 162 | 1 | 10 | 10 |
оПНДНКФЕМХЕ РЮАК. 10.1
| 1 | 2 | 3 | 4 | 5 | 6 |
| 11 | 54 | 2 | 4 | 5 | |
| 12 | 38 | 1 | |||
| 13 | 39 | 6 | |||
| 14 | 48 | 2 | 3 | 4 | |
| 15 | 55 | 1 | 3 | 6 | |
| 16 | 43 | 1 | 1 | 5 | |
| 17 | 44 | 1 | 2 | ||
| 18 | 26 | 1 | |||
| 19 | 14 | 3 | |||
| 21 | 11 | 1 | |||
| 22 | 5 | 1 | |||
| 23 | 8 | 1 | |||
| хРНЦН | 897 | 35 | 28 | 76 |
бЮФМН РЮЙФЕ НРЛЕРХРЭ, ВРН ЖХПЙСКХПНБЮБЬХИ Я 1977 Ц. БХПСЯ A(HlNl) ОЕПЕЯРЮК БШДЕКЪРЭЯЪ. тЮЙР ХЯВЕГМНБЕМХЪ ЩРНЦН БХПСЯЮ ХГ ОНОСКЪЖХХ ВЕКНБЕЙЮ ЙНМЯРЮРХПНБЮКХ Х КЮАНПЮРНПХХ ДПСЦХУ ЯРПЮМ [36]. яНОНЯРЮБКЕМХЕ ОПХБЕДЕММШУ ДЮММШУ ЯН ЯБЕДЕМХЪЛХ Н ПЮЯ╜ОПНЯРПЮМЕМХХ ПЮМЕЕ ОПНУНДХБЬХУ ОЮМДЕЛХИ БШЪБХКН ХМРЕПЕЯМСЧ ГЮЙНМНЛЕПМНЯРЭ: ОЮМДЕЛХЪ 1957-1958 ЦЦ. Б пНЯЯХХ РНФЕ МЮВХМЮ╜КЮЯЭ Б НЙРЪАПЕ, Ю БШГБЮБЬХИ ЕЕ ЯЕПНРХО БХПСЯЮ A(H2N2) Б ОНЯКЕ╜ДСЧЫЕЛ ОЕПЕЬЕК Б ПЮГПЪД ЯЕГНММШУ ЩОХДЕЛХИ Х БШДЕКЪКЯЪ СФЕ Б РХОХВМШИ ДКЪ МЕЕ ОЕПХНД [21]. оНЩРНЛС Б 2009 Ц. ЛНФМН АШКН ОПЕДОНКЮЦЮРЭ БНГЛНФМНЯРЭ ПЮГБХРХЪ ЩОХДЕЛХХ ЦПХООЮ, БШГБЮММНИ МНБШЛ ОЮМДЕЛХВЕЯЙХЛ БХПСЯНЛ, Б МЕРПЮДХЖХНММШИ ДКЪ ОНЯКЕД╜МХУ ДЕЯЪРХКЕРХИ ПЮЯОПНЯРПЮМЕМХЪ ЩРНИ ХМТЕЙЖХХ Б МЮЬЕИ ЯРПЮМЕ БПЕЛЕММНИ ОЕПХНД.
б ЩРНЛ ОКЮМЕ ОПЕДЯРЮБКЪЕР ХМРЕПЕЯ ЩОХДЕЛХНКНЦХВЕЯЙЮЪ ЯХРСЮЖХЪ, ЙНРНПЮЪ ЯКНФХКЮЯЭ БН бЯЕПНЯЯХИЯЙНЛ ДЕРЯЙНЛ ЖЕМРПЕ (бдж) ╚нЙЕЮМ╩, ПЮЯОНКНФЕММНЛ Б ОПХЦНПНДЕ бКЮДХБНЯРНЙЮ. гЮМНЯ ХМТЕЙЖХХ ХГ йХРЮЪ НРДШУЮЧЫЕИ ДЕБНВЙНИ Б ХЧКЕ 2009 Ц. РЮЙ ФЕ МЕ ОНБКЕЙ ГЮ ЯНАНИ ЩОХДЕЛХВЕЯЙНИ БЯОШЬЙХ, УНРЪ Б бдж, ЕЯРЕЯР╜БЕММН, БШЯНЙЮ ЯЙСВЕММНЯРЭ ДЕРЕИ Б ЙНККЕЙРХБЕ, ВРН ДНКФМН АШКН АШ ЯОНЯНАЯРБНБЮРЭ ЮЙРХБХГЮЖХХ ЛЕУЮМХГЛЮ ОЕПЕДЮВХ ОЮМДЕЛХВЕ╜ЯЙНЦН БХПСЯЮ. нДМЮЙН ЩРНЦН МЕ ОПНХГНЬКН. йЮЙ Х БН бКЮДХБНЯРНЙЕ, ЩЙЯОКНГХБМШИ ОНДЗЕЛ ГЮАНКЕБЮЕЛНЯРХ ГДЕЯЭ ОПХЬЕКЯЪ МЮ НЙРЪАПЭ, ЙНЦДЮ ОПЮЙРХВЕЯЙХ ЯПЮГС ОНЯКЕ ГЮЕГДЮ МЮВЮКХ АНКЕРЭ ДЕРХ ХГ ПЮГ╜МШУ НРПЪДНБ. х Б РЕВЕМХЕ ЙНПНРЙНЦН БПЕЛЕМХ ОЕПЕАНКЕКН 82 ВЕК., ЙЮЙ ДЕРЕИ, РЮЙ Х БГПНЯКШУ (10 % НР ВХЯКЕММНЯРХ ЙНККЕЙРХБЮ).
бЯОШЬЙЮ ГЮЙНМВХКЮЯЭ Б ЙНМЖЕ НЙРЪАПЪ, Х Б МНБШУ ЯЛЕМЮУ ГЮАНКЕ╜БЮМХИ МЕ НРЛЕВЕМН.
яКЕДСЕР ГЮЛЕРХРЭ, ВРН НДМНИ ХГ НЯНАЕММНЯРЕИ ЯНБПЕЛЕММНИ ЩОХДЕЛХНКНЦХХ ЯЕГНММНЦН ЦПХООЮ, ЙЮЙ АШКН ЯЙЮГЮМН, ЪБКЪЕРЯЪ РНР ТЮЙР, ВРН ПЮГМШЕ ЬРЮЛЛШ БХПСЯЮ ЖХПЙСКХПСЧР Б ЩОХДЕЛХ╜ВЕЯЙХИ ОЕПХНД НДМНБПЕЛЕММН, МЕ БШРЕЯМЪЪ ДПСЦ ДПСЦЮ, ЙЮЙ ЩРН АШКН ПЮМЕЕ ДН 1977 Ц. нДМЮЙН, ЙЮЙ ОНЙЮГШБЮЧР МЮЬХ МЮАКЧДЕМХЪ ГЮ 1982-2009 ЦЦ., Б ЙЮФДШИ ЩОХДЕЛХВЕЯЙХИ ОЕПХНД, ЙЮЙ ОПЮБХКН, Б оПХЛНПЯЙНЛ ЙПЮЕ АНКЕЕ ГМЮВХЛСЧ ПНКЭ ХЦПЮК ЙЮЙНИ-КХАН НДХМ ХГ ЯЕГНММШУ ЬРЮЛЛНБ БХПСЯЮ ЦПХООЮ ю (Б НЯМНБМНЛ H3N2) ХКХ ЦПХОО б (ПХЯ. 10.1.). оНЯКЕДМХИ, БХДХЛН, ЯОНЯНАЕМ ЙНМЙСПХПНБЮРЭ Я КЧАШЛ ХГ ОЕПЕВХЯКЕММШУ БЮПХЮМРНБ.
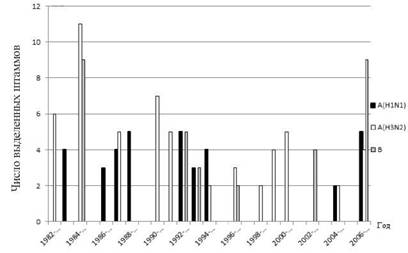
пХЯ. 10.1.бШДЕКЕМХЕ БХПСЯНБ ЦПХООЮ НР ФХРЕКЕИ бКЮДХБНЯРНЙЮ Б ЩОХДЕЛХНКНЦХВЕЯЙХЕ ЯЕГНМШ 1982-2007 ЦЦ.
лЮЯЯНБНЕ БШДЕКЕМХЕ МНБНЦН ОЮМДЕЛХВЕЯЙНЦН БХПСЯЮ ЦПХООЮ ю Б оПХЛНПЯЙНЛ ЙПЮЕ МЮВЮКНЯЭ Я НЙРЪАПЪ 2009 Ц. Х ОПНДНКФЮКНЯЭ ДН ЙНМЖЮ ДЕЙЮАПЪ Х МЮВЮКЮ 2010 Ц. оПХ ЩРНЛ НМ БШРЕЯМХК ХГ ОНОС╜КЪЖХХ БН БПЕЛЪ ЯБНЕИ ЖХПЙСКЪЖХХ БЯЕ ДПСЦХЕ БХПСЯШ ЦПХООЮ, ВРН ЯБХДЕРЕКЭЯРБСЕР Н АНКЕЕ БШЯНЙНИ ЕЦН ЙНМЙСПЕМРНЯОНЯНАМНЯРХ Б ЩРНР ОЕПХНД (ЯЛ. РЮАК. 10.1). бЮФМН ОНДВЕПЙМСРЭ, ВРН МНБШИ БХПСЯ
ЖХПЙСКХПНБЮК ОПЕХЛСЫЕЯРБЕММН Б РЕУ ФЕ БНГПЮЯРМШУ ЦПСООЮУ, ВРН Х БХПСЯ A (H1N1), ЙНРНПШИ Б НЯМНБМНЛ Х БШГБЮК ЯЕГНММШИ ОНДЗ╜ЕЛ ГЮАНКЕБЮЕЛНЯРХ ЦПХООНЛ Б МЮВЮКЕ 2009 Ц. (РЮАК. 10.2), ВЕЛ ОНД╜РБЕПФДЮЕРЯЪ НРЯСРЯРБХЕ ОЕПЕЙПЕЯРМНЦН ХЛЛСМХРЕРЮ ЛЕФДС ЩРХЛХ БХПСЯЮЛХ [19].
рЮАКХЖЮ 10.2
бШДЕКЕМХЕ БХПСЯНБ ЦПХООЮ НР ФХРЕКЕИ бКЮДХБНЯРНЙЮ ПЮГКХВМШУ БНГПЮЯРМШУ ЦПСОО Б 2009 Ц.
| бНГПЮЯР, КЕР | бЯЕЦН НАЯКЕ╜ДНБЮМН, ВЕК. | йНКХВЕЯРБН БШДЕКЕММШУ ЬРЮЛЛНБ | ||||||||
| БЯЕЦН | A(H1N1) | A(H3N2) | B | A(H1N1) pdm09 | ||||||
| ЮАЯ. | % | ЮАЯ. | % | ЮАЯ. | % | ЮАЯ. | % | |||
| 0-2 | 195 | 41 | 23 | 11,4 | 2 | 20 | 0 | 16 | 8,5 | |
| 3-6 | 289 | 55 | 26 | 12,9 | 4 | 40 | 0 | 25 | 13,2 | |
| 7-14 | 384 | 93 | 50 | 24,9 | 0 | 0 | 43 | 23,0 | ||
| 15-20 | 265 | 46 | 21 | 10,4 | 0 | 1 | 6,3 | 22 | 11,7 | |
| 21-30 | 262 | 60 | 25 | 12,4 | 0 | 4 | 25 | 29 | 15,4 | |
| 31-40 | 274 | 64 | 26 | 12,9 | 2 | 20 | 3 | 18,7 | 31 | 16,5 |
| 41-50 | 241 | 45 | 19 | 9,5 | 1 | 10 | 5 | 31,3 | 20 | 10,6 |
| 51-60 | 84 | 14 | 8 | 4,0 | 1 | 10 | 2 | 12,4 | 2 | 1,1 |
| 61 Х | ||||||||||
| ЯРЮПЬЕ | 22 | 4 | 3 | 1,6 | 0 | 1 | 6,3 | 0 | ||
| бЯЕЦН | 1516 | 415 | 201 | 100 | 10 | 100 | 16 | 100 | 188 | 100 |
хМРЕПЕЯМШЕ Б ЩРНЛ ОКЮМЕ ЯБЕДЕМХЪ ОНКСВЕМШ МЮЛХ ОПХ НЖЕМЙЕ ОНОСКЪЖХНММНЦН ХЛЛСМХРЕРЮ С МЮЯЕКЕМХЪ Й ЖХПЙСКХПС╜ЧЫХЛ БХПСЯЮЛ ЦПХООЮ, ОПНБЕДЕММНИ Б ЯННРБЕРЯРБХХ Я ОПХКНФЕ╜МХЕЛ Й оПХЙЮГС тЕДЕПЮКЭМНИ ЯКСФАШ ОН МЮДГНПС Б ЯТЕПЕ ГЮЫХ╜РШ ОПЮБ ОНРПЕАХРЕКЕИ Х АКЮЦНОНКСВХЪ ВЕКНБЕЙЮ НР 31.03.2005 Ц. ╧ 373. б ПЮАНРЕ ХЯОНКЭГНБЮКХ ЙНЛЛЕПВЕЯЙХЕ ЯСУХЕ ЦПХООНГМШЕ ДХЮЦМНЯРХЙСЛШ РХОНБ A(H1N1), A(HSw1N1), A(H3N2) Х B, БШОС╜ЯЙЮЕЛШЕ ннн ╚оодо╩ мхх ЦПХООЮ пюлм. оНЯЙНКЭЙС, ЙЮЙ АШКН ОНЙЮГЮМН БШЬЕ, Б ЙЮФДШИ ЩОХДЕЛХВЕЯЙХИ ОЕПХНД, ЙЮЙ ОПЮБХКН, Б оПХЛНПЯЙНЛ ЙПЮЕ АНКЕЕ ГМЮВХЛСЧ ПНКЭ ХЦПЮК ЙЮЙНИ-КХАН НДХМ ХГ ЯЕГНММШУ ЬРЮЛЛНБ БХПСЯЮ ЦПХООЮ, РН КНЦХВМН АШКН ОПЕДОНКЮЦЮРЭ, ВРН ХЛЕММН Й ЩРНЛС ЬРЮЛЛС С МЮЯЕКЕМХЪ АСДЕР ТНПЛХПНБЮРЭЯЪ ЯЮ╜ЛШИ БШЯНЙХИ СПНБЕМЭ ОНОСКЪЖХНММНЦН ХЛЛСМХРЕРЮ, РЕЛ АНКЕЕ ВРН НМ БУНДХР Б БЮЙЖХМС ОПНРХБ ЦПХООЮ ДКЪ ОПЕДЯЕГНММНИ ХЛЛСМХГЮ╜ЖХЪ МЮЯЕКЕМХЪ.
нДМЮЙН МЕЯЛНРПЪ МЮ РН, ВРН Б ЯЕГНМ 2007-2008 ЦЦ. БЕДСЫХЛ ЩОХДЕЛХВЕЯЙХЛ БЮПХЮМРНЛ АШК ОНДРХО БХПСЯЮ ЦПХООЮ A (H3N2), Б ЙНМРПНКЭМНИ ЦПСООЕ ДНМНПНБ ХЛЕММН Й ЩРНЛС ЬРЮЛЛС Б НЙРЪАПЕ 2008 Ц. АШК БШЪБКЕМ ЯЮЛШИ БШЯНЙХИ ОПНЖЕМР ЯЕПНМЕЦЮРХБМШУ КХЖ (РЮАК. 10.3).
рЮАКХЖЮ 10.3
сПНБЕМЭ ОНОСКЪЖХНММНЦН ХЛЛСМХРЕРЮ Й ПЮГКХВМШЛ БХПСЯЮЛ ЦПХООЮ С ФХРЕКЕИ бКЮДХБНЯРНЙЮ Б 2008-2010 ЦЦ.
| дЮРЮ НАЯКЕДНБЮМХЪ | юМРХЦЕММЮЪ ТНПЛСКЮ ДХЮЦМН ЯРХВЕЯЙНЦН ЬРЮЛЛЮ БХПСЯЮ ЦПХООЮ | йНКХВЕЯРБН НАЯКЕДНБЮММШУ ДНМНПНБ, % | яПЕДМЕЦЕНЛЕР╜ПХВЕЯЙХЕ РХРПШ ЮМРХРЕК | |
| БЯЕЦН | ЯЕПНМЕЦЮ╜РХБМШУ | |||
| нЙРЪАПЭ 2008 Ц. | A(H1N1) | 46 | 17,1 | |
| A(H3N2)* | 70 | 17,1 | ||
| B | 100 | 14 | 39,0 | |
| юОПЕКЭ 2009 Ц. | A(H1N1)* | 42 | 16,0 | |
| A(H3N2) | 24 | 34,0 | ||
| B | 100 | 2 | 60,0 | |
| нЙРЪАПЭ 2009 Ц. | A(H1N1)* | 64 | 10,6 | |
| A(H3N2) | 52 | 19,7 | ||
| B | 100 | 12 | 49,0 | |
| юОПЕКЭ 2010 Ц. | A(H1lN1)pdm09* | 54 | 16,0 | |
| A(H1N1) | 42 | 22,6 | ||
| A(H3N2) | 36 | 24,2 | ||
| B | 100 | 6 | 91,0 | |
| нЙРЪАПЭ 2011 Ц. | A(H1N1)pdmo9* | 58 | 9,2 | |
| A(H1N1) | 15 | 49,0 | ||
| A(H3N2) | 50 | 14,9 | ||
| B | 100 | 12 | 45,0 | |
| юОПЕКЭ 2011 Ц. | A(H1N1)pdm09* | 78 | 5,7 | |
| A(H1N1) | 4 | 91,0 | ||
| A(H3N2) | 50 | 14,9 | ||
| B | 100 | 4 | 84,0 | |
* бЕДСЫХИ БЮПХЮМР БХПСЯЮ.
б 2009 Ц. (ЪМБЮПЭ-ЮОПЕКЭ) МЮХАНКЭЬЕЕ ПЮЯОПНЯРПЮМЕМХЕ ХЛЕК ОНДРХО A(H1N1), НДМЮЙН Х Б ЮОПЕКЕ, Х Б НЙРЪАПЕ Й МЕЛС РЮЙФЕ БШЪБКЕМШ ЛЮЙЯХЛЮКЭМНЕ ЙНКХВЕЯРБН ЯЕПНМЕЦЮРХБМШУ КХЖ Х ЯЮЛШЕ МХГЙХЕ ЯПЕДМЕЦЕНЛЕРПХВЕЯЙХЕ РХРПШ ЮМРХРЕК. б ЮОПЕКЕ 2010 Ц. Й
МНБНЛС ОЮМДЕЛХВЕЯЙНЛС БХПСЯС ЦПХООЮ РЮЙФЕ ГЮПЕЦХЯРПХПНБЮМН МЮХАНКЭЬЕЕ ЙНКХВЕЯРБН ЯЕПНМЕЦЮРХБМШУ ДНМНПНБ Х МЮХЛЕМЭЬХЕ ЯПЕДМЕЦЕНЛЕРПХВЕЯЙХЕ БЕКХВХМШ РХРПНБ ЮМРХРЕК. рНКЭЙН Й БХПСЯС ЦПХООЮ б БН БЯЕ ЮМЮКХГХПСЕЛШЕ ЦНДШ НРЛЕВЕМШ МЕХГЛЕММН МХГЙНЕ ЙНКХВЕЯРБН ЯЕПНМЕЦЮРХБМШУ КХЖ Х БШЯНЙХЕ ЯПЕДМХЕ РХРПШ ЮМРХ╜РЕК, ВРН ЯБХДЕРЕКЭЯРБСЕР Н БШЯНЙНЛ Й МЕЛС СПНБМЧ ОНОСКЪЖХНМ╜МНЦН ХЛЛСМХРЕРЮ. оПХБЕДЕММШЕ ДЮММШЕ ОНГБНКЪЧР СЯНЛМХРЭЯЪ Б РНЛ, ВРН ХЛЕММН БЮЙЖХМЮЖХЪ ОПХБЕКЮ Й ЯМХФЕМХЧ ГЮАНКЕБЮЕЛНЯРХ ЦПХООНЛ. хМЮВЕ ЙЮЙ НАЗЪЯМХРЭ РНР ТЮЙР, ВРН Б ЯЕГНМ 2012-2013 ЦЦ. ОПХ ОПЮЙРХВЕЯЙХ НДХМЮЙНБНЛ НУБЮРЕ МЮЯЕКЕМХЪ ОПХБХБЙЮЛХ Х ОПХ╜ЛЕМЕМХХ НДМХУ Х РЕУ ФЕ БЮЙЖХММШУ ЬРЮЛЛНБ, ЯПЕДХ МЮЯЕКЕМХЪ, ФХБСЫЕЦН Б еБПНОЕИЯЙНИ ВЮЯРХ пт, ОПЕХЛСЫЕЯРБЕММН ЖХПЙСКХПН╜БЮК ОНДРХО A(H1N1)pdm 09, Ю - яХАХПХ Х дЮКЭМЕЦН бНЯРНЙЮ - ОНД╜РХО A(H3N2) (РЮАК. 10.4).
йЮЙ ХГБЕЯРМН, ТНПЛХПНБЮМХЕ С МЮЯЕКЕМХЪ ХЛЛСММНИ ОПН╜ЯКНИЙХ НЙЮГШБЮЕР ПЕЬЮЧЫЕЕ БКХЪМХЕ МЮ ЮМРХЦЕММШЕ ХГЛЕМЕМХЪ БХПСЯЮ Х ГЮЙНМНЛЕПМНЯРХ що. нДМЮЙН МЕ БЯЕ НЯНАЕММНЯРХ ЩОХДЕ╜ЛХНКНЦХХ ЦПХООЮ, БХДХЛН, ЛНФМН НАЗЪЯМХРЭ РНКЭЙН ХМТЕЙЖХНММН╜ХЛЛСМНКНЦХВЕЯЙХЛХ БГЮХЛННРМНЬЕМХЪЛХ. б ВЮЯРМНЯРХ, ОНВЕЛС ХЛЛСМНКНЦХВЕЯЙХИ ОПЕЯЯ ОНЯКЕ 1977 Ц. ОЕПЕЯРЮК БШРЕЯМЪРЭ ХГ ОН╜ОСКЪЖХХ ЖХПЙСКХПСЧЫХЕ БЮПХЮМРШ БХПСЯНБ A(H1N1) Х A(H3N2). й РНЛС ФЕ Б 2010 Ц. ХЯВЕГ БХПСЯ A(H1N1), ОПХ РНЛ ВРН A(H3N2) НЯРЮКЯЪ, УНРЪ НМХ ЖХПЙСКХПНБЮКХ БЛЕЯРЕ Я 1977, Х, ЯКЕДНБЮРЕКЭ╜МН, СПНБЕМЭ ОНОСКЪЖХНММНЦН ХЛЛСМХРЕРЮ Й МХЛ С МЮЯЕКЕМХЪ ЯС╜ЫЕЯРБЕММН МЕ ДНКФЕМ ПЮГКХВЮРЭЯЪ? хКХ ОНВЕЛС, МЮОПХЛЕП, ОПХ МЮКХВХХ ЕЫЕ АНКЭЬНЦН ЙНКХВЕЯРБЮ МЕХЛЛСММШУ КЧДЕИ Й МНБНЛС ОЮМДЕЛХВЕЯЙНЛС БХПСЯС Б ЩОХДЕЛХВЕЯЙХИ ЯЕГНМ 2009-2010 ЦЦ. МЮ ЯЛЕМС ЕЛС Б ЙНМЖЕ ЩОХДЕЛХВЕЯЙНЦН ЯЕГНМЮ (ЯЛ. РЮАК. 9.4) ОПХЬЕК БХПСЯ ЦПХООЮ б? оПХВЕЛ ОНЯКЕДМХИ, МЕЯЛНРПЪ МЮ ТНПЛХПНБЮМХЕ Й МЕЛС С МЮЯЕКЕМХЪ БШЯНЙНЦН СПНБМЪ ОНОСКЪЖХНММНЦН ХЛЛСМХРЕРЮ Х ГМЮВХРЕКЭМН ЛЕМЭЬСЧ ХГЛЕМВХБНЯРЭ ОН ЯПЮБМЕМХЧ Я БХПСЯЮЛХ ЦПХООЮ ю ОПНДНКФЮЕР ЮЙРХБМН ЖХПЙСКХПНБЮРЭ? яКЕДСЕР ГЮЛЕРХРЭ, ВРН МЕЙНРНПШЕ ЯНЛМЕМХЪ ОН ОНБНДС БКХЪМХЪ ХЛЛСМНКНЦХВЕЯЙНЦН ОПЕЯЯЮ МЮ що ЦПХООЮ БШЯЙЮГШБЮКХЯЭ Х ПЮМЕЕ ╚... МЕОНМЪРМН, МЮ╜ОПХЛЕП, ОНВЕЛС ХЛЛСМНКНЦХВЕЯЙХИ ОПЕЯЯ МЕ ДЕИЯРБСЕР МЮ ЮМРХЦЕ╜МШ БХПСЯНБ ЙНПХ ХКХ ОНКХНЛХЕКХРЮ, ЯРНКЭ ФЕ САХЙБХРЮПМШЕ ЙЮЙ Х БХПСЯШ ЦПХООЮ╩ [5].
б ЩРНИ ЯБЪГХ НРЛЕРХЛ, ВРН Б ПЮГПЮАНРЮММНИ МЮЛХ ЙНМЖЕО╜ЖХХ ХМРЕЦПЮЖХНММН-ЙНМЙСПЕМРМНЦН ПЮГБХРХЪ що, СВХРШБЮЧРЯЪ МЕ РНКЭЙН ХМТЕЙЖХНММН-ХЛЛСМНКНЦХВЕЯЙХЕ БГЮХЛННРМНЬЕМХЪ ЛЕФДС ОНОСКЪЖХЕИ ВЕКНБЕЙЮ Х ЛХЙПННПЦЮМХГЛНБ, МН Х БНГЛНФМНЕ БКХЪМХЕ МЮ що ЩЙНКНЦХВЕЯЙНЦН ТЮЙРНПЮ, ОНД ЙНРНПШЛ ОНМХЛЮЧРЯЪ ТНПЛХ╜ПСЧЫХЕЯЪ Б АХНЖЕМНГЕ ХМРЕЦПЮЖХНММН-ЙНМЙСПЕМРМШЕ ЯБЪГХ ЛЕФДС ПЮГКХВМШЛХ ЛХЙПННПЦЮМХГЛЮЛХ (ЯЛ. ЦК. 8). оНЩРНЛС НАЗЪЯМЕМХЕ НЯНАЕММНЯРЕИ ЩОХДЕЛХНКНЦХХ ЦПХООЮ, ЙЮЙ ЛШ ОНКЮЦЮЕЛ, ЯКЕДСЕР ХЯЙЮРЭ Х Б ДЮММНЛ МЮОПЮБКЕМХХ, СВХРШБЮЪ БНГЛНФМНЯРЭ РЕУ ХКХ ХМШУ БГЮХЛНДЕИЯРБХИ ЛЕФДС ПЮГКХВМШЛХ ЯЕПНРХОЮЛХ Х ОНДРХОЮ╜ЛХ БХПСЯНБ. н РНЛ, ВРН ЙНМЙСПЕМРМШЕ БГЮХЛННРМНЬЕМХЪ ЛЕФДС БХ╜ПСЯЮЛХ ЦПХООЮ ХЛЕЧР ЛЕЯРН, ЯБХДЕРЕКЭЯРБСЧР, МЮОПХЛЕП, ПЕГСКЭ╜РЮРШ БШДЕКЕМХЪ БХПСЯНБ Б ПЮГКХВМШУ ТЕДЕПЮКЭМШУ НЙПСЦЮУ пт Б ОЕПХНД 2012-2013 ЦЦ. (РЮАК. 10.4).
рЮАКХЖЮ 10.4
жХПЙСКЪЖХЪ БХПСЯНБ ЦПХООЮ Б ПЮГКХВМШУ ТЕДЕПЮКЭМШУ НЙПСЦЮУ пт Б ОЕПХНД ЯЕГНММНИ ЩОХДЕЛХХ 2012-2013 ЦЦ.
| нЙПСЦ | бЯЕЦН БШДЕКЕМН ЬРЮЛЛНБ | бХПСЯ, ЮАЯ. ВХЯКН (%) | |||
| A(H1N1) pdm | A(H3N2) | A МЕЯСАРХ- ОХПНБЮМ╜МШИ | B | ||
| жЕМРПЮКЭМШИ | 281 | 142 (50,5) | 28 (9,9) | 111 (39,5) | |
| оПХБНКФЯЙХИ | 57 | 28 (49,1) | 14 (24,6) | 15 (26,3) | |
| яЕБЕПН-гЮОЮДМШИ | 458 | 203 (44,3) | 76 (16,6) | 179 (36,1) | |
| чФМШИ | 65 | 19 (29,2) | 16 (24,6) | 5 (7,7) | 25 (38,5) |
| сПЮКЭЯЙХИ | 53 | 36 (67,9) | 1 (1,9) | 16 (30,2) | |
| яХАХПЯЙХИ | 274 | 8 (2,9) | 121 (44,2) | 4 (1,4) | 141 (51,4) |
| дЮКЭМЕБНЯРНВМШИ | 150 | 24 (16) | 117 (78,5) | 9 (6,0) | |
| оПХЛНПЯЙХИ ЙПЮИ | 104 | 6 (5,7) | 92 (88,5) | 6 (5,7) | |
| хРНЦН | 1338 | 460 (34,4) | 373 (27,9) | 9 (0,7) | 496 (37,0) |
рЮЙ, Б АНКЭЬХМЯРБЕ ГЮОЮДМШУ ПЕЦХНМНБ ДНЛХМХПНБЮК ОЮМДЕ╜ЛХВЕЯЙХИ БЮПХЮМР БХПСЯЮ ЦПХООЮ ю, РНЦДЮ ЙЮЙ ГЮ сПЮКНЛ, ОПЕХЛС╜ЫЕЯРБЕММН БШДЕКЪКХ БХПСЯ ЦПХООЮ A(H3N2). рНКЭЙН БХПСЯ ЦПХООЮ б, ЙЮЙ БХДМН, ЛНФЕР СЯОЕЬМН ЙНМЙСПХПНБЮРЭ Я РЕЛ Х ДПСЦХЛ БХ╜ПСЯЮЛХ ЦПХООЮ ю. мН ОПХ ЩРНЛ ЯКЕДСЕР СВЕЯРЭ, ВРН НМ МЮВХМЮЕР ЮЙРХБМН БШДЕКЪРЭЯЪ, ЙЮЙ ОПЮБХКН, ОН ГЮБЕПЬЕМХХ ЯЕГНММНИ ЩОХ-
ДЕЛХХ, БШГБЮММНИ БХПСЯЮЛХ ЦПХООЮ ю. яКЕДНБЮРЕКЭМН, Х МЮ ЮЙРХБ╜МНЯРЭ ЕЦН ЖХПЙСКЪЖХХ, БХДХЛН, ЩРХ БХПСЯШ БКХЪЧР.
й ЯНФЮКЕМХЧ, ЩЙЯОЕПХЛЕМРЮКЭМШЕ ХЯЯКЕДНБЮМХЪ, Б ЙНРНПШУ АШ ХГСВЮКНЯЭ БГЮХЛНДЕИЯРБХЕ БХПСЯНБ ЦПХООЮ ЛЕФДС ЯНАНИ Х ДПС╜ЦХЛХ БХПСЯЮЛХ, ЕДХМХВМШ. рЮЙ, я.т. ьЕМДЕПНБХВ Х к.ъ. гЮЙЯРЕКЭ- ЯЙЮЪ (1967 Ц.) ОПНБНДХКХ ХГСВЕМХЕ БГЮХЛНДЕИЯРБХЪ ПЮГКХВМШУ БХ╜ПСЯНБ (ЦПХООЮ A(H2,N2) Х ОЮПЮЦПХООЮ 2 Х 3 ЯЕПНРХОНБ, Ю РЮЙФЕ БХПСЯЮ A(H2N2) Х ЮДЕМНБХПСЯЮ 2 ЯЕПНРХОЮ) ОПХ ЮЯЯНЖХХПНБЮММНЛ ГЮПЮФЕМХХ ЙСКЭРСПШ РЙЮМХ КЕЦЙХУ ЩЛАПХНМЮ ВЕКНБЕЙЮ. юБРНПШ СЯРЮМНБХКХ, ВРН Б ПЕГСКЭРЮРЕ ЯНБЛЕЯРМНЦН ПЮГЛМНФЕМХЪ БХПСЯНБ ЦПХООЮ Х ОЮПЮЦПХООЮ 3 ПЕОПНДСЙЖХЪ ЦПХООНГМНЦН БНГАСДХРЕКЪ ПЕГЙН ЯМХФЮЕРЯЪ, ОПХВЕЛ ЯРЕОЕМЭ ОНДЮБКЪЧЫЕИ ЮЙРХБМНЯРХ БХПС╜ЯЮ ОЮПЮЦПХООЮ 3 ГЮБХЯХР НР ОНЯКЕДНБЮРЕКЭМНЯРХ ХМТХЖХПНБЮМХЪ ЙСКЭРСПШ РЙЮМХ Х НР ХМРЕПБЮКНБ ЛЕФДС ГЮПЮФЕМХЕЛ ЕЕ ОЕПБШЛ Х БРНПШЛ ЮЦЕМРНЛ. оПХ ГЮПЮФЕМХХ ЙСКЭРСПШ РЙЮМХ ЯМЮВЮКЮ БХПСЯНЛ ОЮПЮЦПХООЮ 3, Ю ГЮРЕЛ ЦПХООЮ ОНДЮБКЪЧЫЕЕ ДЕИЯРБХЕ ОЕПБНЦН МЮ БРНПНИ МЮХАНКЕЕ БШПЮФЕМН. бКХЪМХЕ БХПСЯЮ ОЮПЮЦПХООЮ 3 МЮ ПЕ╜ОПНДСЙЖХЧ БХПСЯЮ ЦПХООЮ A ЯРЮМНБХРЯЪ ЯКЮАЕЕ ОПХ НДМНБПЕЛЕМ╜МНЛ ГЮПЮФЕМХХ ХКХ ГЮПЮФЕМХХ ЙСКЭРСПШ РЙЮМХ ЯМЮВЮКЮ БХПСЯНБ ЦПХООЮ, Ю ВЕПЕГ 24 В - БХПСЯНЛ ОЮПЮЦПХООЮ 3. оПХ ЩРНЛ ЯМХФЮЕРЯЪ РЮЙФЕ Х ХМРЕМЯХБМНЯРЭ ПЕОПНДСЙЖХХ ОЮПЮЦПХООЮ 3.
б ОНДНАМШУ НОШРЮУ Я БХПСЯЮЛХ A(H2N2) Х ОЮПЮЦПХООЮ 2 ОН╜КСВЕМШ ЮМЮКНЦХВМШЕ ПЕГСКЭРЮРШ: ОНДЮБКЪЧЫХЛ АШК БХПСЯ, ЙНРН╜ПШИ ББНДХКЯЪ ОЕПБШЛ ОПХ ПЮГДЕКЭМНЛ ХМТХЖХПНБЮМХХ. б ПЕГСКЭ╜РЮРЕ НДМНБПЕЛЕММНЦН ГЮПЮФЕМХЪ ОПНХЯУНДХКН ВЮЯРХВМНЕ СЦМЕРЕМХЕ ПЕОПНДСЙЖХХ НАНХУ БХПСЯНБ. оПХ БНЯОПНХГБЕДЕМХХ ЮЯЯНЖХХПНБЮМ╜МНИ ХМТЕЙЖХХ БХПСЯНБ ЦПХООЮ A Х ЮДЕМНБХПСЯНБ СЯРЮМНБКЕМН, ВРН РЕ Х ДПСЦХЕ КСВЬЕ ПЕОПНДСЖХПСЧРЯЪ Б ВХЯРНИ ЙСКЭРСПЕ. йНЦДЮ БНЯ╜ОПНХГБНДХРЯЪ ОНЯКЕДНБЮРЕКЭМНЕ ГЮПЮФЕМХЕ, ХМРЕМЯХБМЕЕ ПЮГЛМН╜ФЮЕРЯЪ РНР БХПСЯ, ЙНРНПШИ АШК ББЕДЕМ ОЕПБШЛ. оПХ НДМНБПЕЛЕМ╜МНЛ ГЮПЮФЕМХХ ЙСКЭРСПШ ЩРХЛХ ЮЦЕМРЮЛХ ОПНХЯУНДХР ВЮЯРХВМНЕ СЦМЕРЕМХЕ ПЕОПНДСЙЖХХ НАНХУ БХПСЯНБ. яКЕДНБЮРЕКЭМН, ЯНВЕРЮМ╜МЮЪ ХМТЕЙЖХЪ, ЙЮЙ ЩРН АШКН ОНЙЮГЮМН МЮЛХ Б ЦК. 7 Х Б ЯКСВЮЕ Я ЦПХООНГМНИ ХМТЕЙЖХЕИ, БХДХЛН, ХЦПЮЕР Б ЩОХДЕЛХВЕЯЙНЛ ОПНЖЕЯ╜ЯЕ ПЕЦСКХПСЧЫСЧ ПНКЭ
бННАЫЕ ЯЛЕМЮ ЬРЮЛЛНБ БНГАСДХРЕКЪ Б ЩОХДЕЛХВЕЯЙНЛ ОПН╜ЖЕЯЯЕ ОПХЯСЫЮ, ЙЮЙ ХГБЕЯРМН, МЕ РНКЭЙН ЦПХООНГМНИ ХМТЕЙЖХХ. х 224
НМЮ, ГЮВЮЯРСЧ, МЕ ЯБЪГЮМЮ Я ХГЛЕМЕМХЕЛ ЯРЕОЕМХ БНЯОПХХЛВХБНЯРХ ОНОСКЪЖХХ. мЮОПХЛЕП, ЯЛЕМЮ ЯЕПНРХОНБ ЯРПЕОРНЙНЙЙЮ ЦПСООШ ю ОПХБНДХР Й ОНЪБКЕМХЧ АНКЕЕ РЪФЕКШУ ТНПЛ АНКЕГМХ Х НАНЯРПЕ╜МХЧЩОХДЕЛХНКНЦХВЕЯЙНИ ЯХРСЮЖХХ [37]. яЛЕМЮ ЬРЮЛЛНБ Б УНДЕ ПЮГБХРХЪ що Б ЛМНЦНКЕРМЕИ ДХМЮЛХЙЕ ОПНЯКЕФЕМЮ Х ОПХ ЛЕМХМ╜ЦНЙНЙЙНБНИ ХМТЕЙЖХХ [38]. оНДЗЕЛ ГЮАНКЕБЮЕЛНЯРХ Б 90-У ЦНДЮУ ОПНЬКНЦН БЕЙЮ ДХТРЕПХЕИ Б пНЯЯХХ ЯНБОЮК МЕ РНКЭЙН Я ПЕГЙХЛ ЯМХ╜ФЕМХЕЛ НУБЮРЮ ОПХБХБЙЮЛХ, МН Х Я БНГБПЮРНЛ Б ОНОСКЪЖХЧ АНКЕЕ ЮЦПЕЯЯХБМНЦН АХНБЮПЮ - ЦПЮБХЯ [39]. нДМЮЙН ОНДНАМШЕ ХГЛЕМЕМХЪ УЮПЮЙРЕПМШ Х ДКЪ ХМТЕЙЖХИ Я ДПСЦХЛХ, ЛЕМЕЕ ЮЙРХБМШЛХ, ЛЕУЮ╜МХГЛЮЛХ ОЕПЕДЮВХ. мЮОПХЛЕП, НАЫЕХГБЕЯРМЮ ЯХРСЮЖХЪ Я ХГЛЕМЕМХ╜ЕЛ БЕДСЫЕЦН БЮПХЮМРЮ ОПХ ЬХЦЕККЕГЮУ [40].
хМРЕПЕЯМШЕ Б ЩРНЛ ОКЮМЕ ДЮММШЕ МЮЛХ ОНКСВЕМШ ОПХ ХГСВЕ╜МХХ ЩРХНКНЦХВЕЯЙНИ ЯРПСЙРСПШ БХПСЯМШУ ЦЕОЮРХРНБ Б оПХЛНПЯЙНЛ ЙПЮЕ Х пЕЯОСАКХЙЕ яЮУЮ (ъЙСРХЪ) [41, 42] ГЮ НДХМ Х РНР ФЕ БПЕЛЕМ╜МНИ ОПНЛЕФСРНЙ (ПХЯ. 10.2).
рЮЙ, ЯННРМНЬЕМХЕ ДНКХ цю, ХЛЕЧЫЕЦН, ЙЮЙ ХГБЕЯРМН, ТЕЙЮКЭ╜МН-НПЮКЭМШИ ЛЕУЮМХГЛ ОЕПЕДЮВХ, Я СДЕКЭМШЛ БЕЯНЛ ЦЕОЮРХРНБ б Х я, ПЮЯОПНЯРПЮМЪЧЫХУЯЪ Я ОНЛНЫЭЧ ЦЕЛНЙНМРЮЙРМНЦН ЛЕУЮМХГ╜ЛЮ, ХЛЕКН НОПЕДЕКЕММСЧ ГЮЙНМНЛЕПМНЯРЭ. оПХ ЩРНЛ, ЙЮЙ БХДМН МЮ ПХЯ. 10.2, ХГЛЕМЕМХЪ Б ЩРХНКНЦХВЕЯЙНИ ЯРПСЙРСПЕ БХПСЯМШУ ЦЕОЮ╜РХРНБ, ЖХПЙСКХПСЧЫХУ ЯПЕДХ МЮЯЕКЕМХЪ йПЮИМЕЦН яЕБЕПЮ Х ЧЦЮ дЮКЭМЕЦН бНЯРНЙЮ, АШКХ ОПЮЙРХВЕЯЙХ ЯХМУПНММШЛХ, ВРН ЯКНФМН НАЗЪЯМХРЭ БКХЪМХЕЛ МЮ МЕ╦ РНКЭЙН ЮЙРХБМНЯРХ РЕУ ХКХ ХМШУ ОСРЕИ ОЕПЕДЮВХ СЙЮГЮММШУ ХМТЕЙЖХИ.
оПХБЕДЕММШЕ ДЮММШЕ, ОН МЮЬЕЛС ЛМЕМХЧ, ЯБХДЕРЕКЭЯРБСЧР Н РНЛ, ВРН ХГЛЕМЕМХЪ Б ЩРХНКНЦХВЕЯЙНИ ЯРПСЙРСПЕ БНГАСДХРЕКЕИ МЕГЮБХЯХЛН НР ХУ ОПХМЮДКЕФМНЯРХ Й РНИ ХКХ ХМНИ ЦПСООЕ ХМТЕЙ╜ЖХИ, БХДХЛН, - ЪБКЕМХЕ НАЫЕАХНКНЦХВЕЯЙНЕ Х НАСЯКНБКЕМН ЛЕУЮ╜МХГЛЮЛХ ЯЮЛНПЕЦСКЪЖХХ ОЮПЮГХРЮПМШУ ЯХЯРЕЛ Б ТНПЛХПСЕЛНЛ ХЛ АХНЦЕНЖЕМНГЕ (ЯЛ. ЦК. 8). б ЙНМЕВМНЛ ХРНЦЕ ЩРН МЮОПЮБКЕМН МЮ ЯН╜УПЮМЕМХЕ ЯРЮАХКЭМНЯРХ ЯСЫЕЯРБНБЮМХЪ ОЮПЮГХРЮПМШУ ЯХЯРЕЛ [43]. дПСЦНЕ ДЕКН, ВРН ОПХ ЦПХООЕ СЙЮГЮММШЕ ОПНЖЕЯЯШ ХДСР ЦНПЮГДН АН╜КЕЕ ЮЙРХБМН, ВРН БН ЛМНЦНЛ НАСЯКНБКЕМН АНКЕЕ БШЯНЙНИ ЙНМРЮЦХНГ- МНЯРЭЧ БХПСЯЮ Х ЕЦН БШПЮФЕММНИ ЯОНЯНАМНЯРЭЧ Й ХГЛЕМВХБНЯРХ. оНЩРНЛС ЯЛЕМЮ ЖХПЙСКХПСЧЫХУ ЬРЮЛЛНБ ОПХ ЦПХООЕ ХЛЕЕР АНКЕЕ
ЯСЫЕЯРБЕММСЧ ЩОХДЕЛХНКНЦХВЕЯЙСЧ ГМЮВХЛНЯРЭ Х ЯННРБЕРЯРБЕМ-
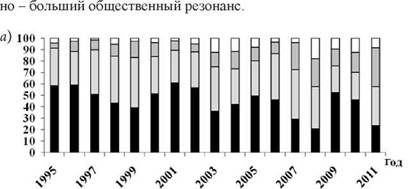
■цЕОЮРОРю ? цЕОЮРХР б ? ΓeπaτnτC ? оПНВХЕ БХПСЯМШЕ ЦЕОЮРХРШ
пХЯ. 10.2.щРХНКНЦХВЕЯЙЮЪ ЯРПСЙРСПЮ БХПСЯМШУ ЦЕОЮРХРНБ Б оПХЛНПЯЙНЛ ЙПЮЕ (Ю) Х пЕЯОСАКХЙЕ яЮУЮ (ъЙСРХЪ) (А) ГЮ 1995-2012 ЦЦ.
яКЕДСЕР ГЮЛЕРХРЭ, ВРН МЕЙСЧ БЮФМСЧ ПЕЦСКХПСЧЫСЧ ПНКЭ БН БГЮХЛННРМНЬЕМХЪУ ЛЕФДС БХПСЯЮЛХ ЦПХООЮ, БНГЛНФМН, ХЦПЮЧР ОЮМ╜ДЕЛХХ. б ВЮЯРМНЯРХ, ОНЯКЕ ОЮМДЕЛХХ ЦПХООЮ Б ЙНМЖЕ XIX Б., БХПСЯ ЦПХООЮ СЙНПЕМХКЯЪ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ, Ю ГЮЙНМНЛЕПМНЯРХ ЕЦН ЩОХДЕЛХНКНЦХХ ОПХМЪКХ ЯНБПЕЛЕММШИ УЮПЮЙРЕП [1]. оНЯКЕ ОЮМ╜
ДЕЛХХ 1977 Ц. ОПНХГНЬКХ ЙЮПДХМЮКЭМШЕ ХГЛЕМЕМХЪ Б ЩРХНКНЦХВЕ╜ЯЙНИ ЯРПСЙРСПЕ ЦПХООЮ Х ЕЦН ЩОХДЕЛХНКНЦХВЕЯЙНИ УЮПЮЙРЕПХЯРХЙЕ. хЛЕММН ОНЯКЕ ОЮМДЕЛХХ 2009-2010 Ц. БХПСЯ A(H1N1) БМНБЭ ХЯВЕГ ХГ ОНОСКЪЖХХ. оНЯЙНКЭЙС Б XX Б. НМ НРЙПШБЮК ЯВЕР ОЮМДЕЛХЪЛ, Ю ГЮРЕЛ БШГШБЮК ЩОХДЕЛХХ Б 40-50-У ЦНДЮУ ОПНЬКНЦН БЕЙЮ, ХЯВЕГЮК Х БМНБЭ БНГБПЮЫЮКЯЪ Б ОНОСКЪЖХЧ, РН, ОН-БХДХЛНЛС, РНКЭЙН АСДСЫЕЕ ОНЙЮФЕР, ПЮЯЯРЮКХЯЭ КХ ЛШ Я ЩРХЛ БХПСЯНЛ МЮБЯЕЦДЮ? хЛЕММН РЮЙ ЛШ ОХЯЮКХ Б ЯБНЕИ ЯРЮРЭЕ Б 2012 Ц., ЮМЮКХГХПСЪ ХРНЦХ ОЮМДЕЛХХ ЦПХООЮ 2009-2010 ЦЦ. [43]. нДМЮЙН СФЕ Б 2014 Ц. БХПСЯ A(H1N1) Б ЕДХМХВМШУ ЯКСВЮЪУ БМНБЭ АШК БШДЕКЕМ КЮАНПЮРНПХЪЛХ МЕЙНРНПШУ ЯРПЮМ чФМНЦН ОНКСЬЮПХЪ. х, МЮ МЮЬ БГЦКЪД, ЯКНФМН ОПНЦМНГХПН╜БЮРЭ, ЙЮЙ ЕЦН БНГБПЮЫЕМХЕ Б ОНОСКЪЖХЧ ЛНФЕР ЯЙЮГЮРЭЯЪ МЮ ЩОХ╜ДЕЛХНКНЦХВЕЯЙНИ ЯХРСЮЖХХ. й РНЛС ФЕ ЩРНР БЮПХЮМР МЕ БУНДХР Б ЯНЯРЮБ ЯНБПЕЛЕММШУ БЮЙЖХМ ОПНРХБ ЦПХООЮ.
яКЕДСЕР ГЮЛЕРХРЭ, ВРН, ОН ЛМЕМХЧ й.ц. сЛЮМЯЙНЦН [44], БХПС╜ЯШ - ОНКМНОПЮБМШЕ, БЕГДЕЯСЫХЕ Х МЕНАУНДХЛШЕ ГБЕМЭЪ АХНЯТЕПШ, ПНКЭ ЙНРНПШУ (Б НЯМНБМНЛ) ЯБНДХРЯЪ Й СВЮЯРХЧ Б ЮДЮОРЮЖХНММШУ ОПНЖЕЯЯЮУ, ОПХВЕЛ МЕ РНКЭЙН Б ЮДЮОРЮЖХХ Й СЯКНБХЪЛ ЯПЕДШ НАХРЮ╜МХЪ, МН Х Й БГЮХЛМНИ ЮДЮОРЮЖХХ ПЮГКХВМШУ НПЦЮМХГЛНБ (ЯЛ. ЦК. 8). йЮЙ ЯВХРЮЕР б.м. ъЦНДХМЯЙХИ, ╚ ...ЛНФЕР АШРЭ ЩОХДЕЛХХ БХПСЯ╜МШУ Х ДПСЦХУ ХМТЕЙЖХИ ХЛЕЧР ХЯЙКЧВХРЕКЭМН БЮФМНЕ ГМЮВЕМХЕ, ОНЯЙНКЭЙС Б ЩРН БПЕЛЪ ОЕПЕМНЯХРЯЪ ЦЕМЕРХВЕЯЙЮЪ ХМТНПЛЮЖХЪ НР ЛХЙПННПЦЮМХГЛНБ Й КЧДЪЛ Х НАПЮРМН╩ [45]. б ЩРНИ ЯБЪГХ ЛНФМН ДС╜ЛЮРЭ, ВРН ОЮМДЕЛХВЕЯЙНЕ ПЮЯОПНЯРПЮМЕМХЕ БХПСЯЮ ЦПХООЮ ЪБКЪЕРЯЪ ТЮЙРНПНЛ, НФХБКЪЧЫХЛ (ПЮЯЙЮВХБЮЧЫХЛ) ЯЙКНММШИ Й ЯЮЛНОПН╜ХГБНКЭМНЛС ГЮРСУЮМХЧ ОПНЖЕЯЯ ЮДЮОРЮЖХХ. оНДРБЕПФДЕМХЕЛ ЩРНИ БЕПЯХХ ЯКСФЮР ЛЮРЕПХЮКШ Н ЯСЫЕЯРБЕММШУ ХГЛЕМЕМХЪУ Б ЩОХДЕ╜ЛХНКНЦХХ ЦПХООЮ Б ЙНМЖЕ ОПНЬКНЦН - МЮВЮКЕ МШМЕЬМЕЦН БЕЙЮ, ВРН ОНГБНКХКН ч.ц. хБЮММХЙНБС Я ЯНЮБР. [1] МЮ НЯМНБЕ ЮМЮКХГЮ ГЮАН╜КЕБЮЕЛНЯРХ МЮЯЕКЕМХЪ ЦПХООНЛ A ДЮФЕ ГЮЪБХРЭ Н БНГЛНФМНЛ ОН╜ЯРЕОЕММНЛ ОПЕЙПЮЫЕМХИ ЩОХДЕЛХИ ЦПХООЮ, БШГБЮММШУ ЩРХЛ БХПС╜ЯНЛ. б ЩРНЛ ОКЮМЕ ЯКЕДСЕР РЮЙФЕ НРЛЕРХРЭ Х РНР ТЮЙР, ВРН Б ОЕПХНД ОЮМДЕЛХХ БХПСЯЮ A(H1N1)pdm09 БХПСЯНКНЦХВЕЯЙЮЪ КЮАНПЮРНПХЪ ╚жЕМРПЮ ЦХЦХЕМШ Х ЩОХДЕЛХНКНЦХХ ОН оПХЛНПЯЙНЛС ЙПЮЧ╩ БШДЕ╜КЪКЮ МЮХАНКЭЬЕЕ ЙНКХВЕЯРБН ПЮГКХВМШУ ЬРЮЛЛНБ БХПСЯЮ ЦПХООЮ, ОПХВЕЛ МЮ ОПНРЪФЕМХХ БЯЕЦН ЦНДЮ, Р.Е. Х Б КЕРМХИ ОЕПХНД, Б НРКХВХЕ НР ДПСЦХУ КЕР.
оНДБНДЪ ХРНЦ БШЬЕХГКНФЕММНЛС, ЛНФМН ОПХИРХ Й ГЮЙКЧВЕ╜МХЧ, ВРН Б РЕНПЕРХВЕЯЙНЛ ОКЮМЕ ОЮМДЕЛХЪ ЦПХООЮ 2009-2010 ЦЦ. ЯОН╜ЯНАЯРБНБЮКЮ ОЕПЕНЯЛШЯКЕМХЧ МЕЙНРНПШУ БНОПНЯНБ ЩОХДЕЛХНКНЦХХ ЩРНИ ХМТЕЙЖХХ. бЛЕЯРЕ Я РЕЛ, МЕЯЛНРПЪ МЮ РН, ВРН БНГНАКЮДЮКЮ ГН- НЮМРПНОНМНГМЮЪ РЕНПХЪ ТНПЛХПНБЮМХЪ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ ЦПХООЮ, ЛНФМН КХ АШРЭ СБЕПЕММШЛ, ВРН Б ОНОСКЪЖХЧ СФЕ МХЙНЦДЮ МЕ БЕПМЕРЯЪ БХПСЯ A(H2N2)? ю ПЮГБЕ ХЯЙКЧВЕМЮ БНГЛНФМНЯРЭ СБЕ╜КХВЕМХЪ ЩОХДЕЛХВЕЯЙНЦН ОНРЕМЖХЮКЮ БХПСЯЮ ЦПХООЮ я? мЕЪЯМШЛХ НЯРЮЧРЯЪ Х БНОПНЯШ, ЙЮЯЮЧЫХЕЯЪ МЕЙНРНПШУ ДПСЦХУ НЯНАЕММНЯРЕИ ЩОХДЕЛХНКНЦХХ ЦПХООЮ. б ЩРНИ ЯБЪГХ ЛШ ОНКЮЦЮЕЛ, ВРН МЮ НЯМНБЕ РНКЭЙН РПЮДХЖХНММШУ ОПЕДЯРЮБКЕМХИ НА що ЩРХ БНОПНЯШ МЕ ПЕ╜ЬХРЭ. мЕКЭГЪ МЕ ЯНЦКЮЯХРЭЯЪ Я ЛМЕМХЕЛ б.х. оНЙПНБЯЙНЦН Я ЯНЮБР. [46], ВРН ╚ ЦКСАНЙХЕ ЩЙНКНЦХВЕЯЙХЕ ╚ЙНПМХ╩ ХМТЕЙЖХНММНИ ОЮРН╜КНЦХХ ДН ЯХУ ОНП МЕ ОПХБКЕЙЮЧР ДНКФМНЦН БМХЛЮМХЪ, НЯРЮБЮЪЯЭ ЯКЮАНХГСВЕММШЛХ╩, Б ВЮЯРМНЯРХ ╚СУНД╩ Х ╚ОПХУНД╩ ПЮГКХВМШУ ХМ╜ТЕЙЖХНММШУ АНКЕГМЕИ, ЙЮЙ ОПЮБХКН, МЕ НЯЛШЯКХБЮЕРЯЪ Я ЩЙНКНЦХ╜ВЕЯЙХУ ОНГХЖХИ.
аХАКХНЦПЮТХВЕЯЙХИ ЯОХЯНЙ
1. хБЮММХЙНБ ч.ц., хЯЛЮЦСКНБ ю.р. щОХДЕЛХНКНЦХЪ ЦПХО╜ОЮ. - юКЛЮ-юРЮ, 1983. - 204 Я.
2. нМХЫЕМЙН ц.ц., еФКНБЮ е.а., кЮГХЙНБЮ ц.т. Х ДП. оЮМДЕЛХЪ ЦПХООЮ A/H1N1/09 Б ЛХПЕ Х пНЯЯХИЯЙНИ тЕДЕПЮЖХХ Б 2009-2010 ЦЦ. Х ОПНЦМНГ МЮ 2010-2011 ЦЦ. // фСПМ. ЛХЙПНАХНК.
- 2010. - ╧ 6. - я. 12-17.
3. хБЮММХЙНБ ч.ц. нЯНАЕММНЯРХ ЩОХДЕЛХИ ЦПХООЮ Х РЕМ╜ДЕМЖХХ ЕЕ ХГЛЕМЕМХЪ // бЕЯРМХЙ пюлм. - 1995. - ╧ 9. - я. 3-7.
4. щМДПЧЯ й. еЯРЕЯРБЕММЮЪ ХЯРНПХЪ БХПСЯНБ. - л., 1969.
- я. 64-100.
5. аЕКЪЙНБ б.д., цНКСАЕБ д.а., йЮЛХМЯЙХИ ц.д., рЕЖ б.б. яЮЛНПЕЦСКЪЖХЪ ОЮПЮГХРЮПМШУ ЯХЯРЕЛ. - к., 1987. - 239 Я.
6. аЕКЪЙНБ б.д., ъТЮЕБ п.у. щОХДЕЛХНКНЦХЪ. - л., 1989.
- я. 186-200.
7. яКЕОСЬЙХМ ю.м., кЭБНБ д.й., лЮПХМХВ х.ц. Х ДП. щОХДЕЛХНКНЦХВЕЯЙХЕ НЯНАЕММНЯРХ ЦПХООЮ ОНЯКЕДМХУ КЕР // бНОП. БХПСЯНК. - 1998. - ╧ 2. - я. 59-62.
8. яКЕОСЬЙХМ ю.м. яНБПЕЛЕММШЕ НЯНАЕММНЯРХ ЩОХДЕЛХН╜КНЦХХ Х ОПНТХКЮЙРХЙХ ЦПХООЮ // фСПМ. ЛХЙПНАХНК. - 2001. - ╧ 1.
- я. 95-99.
9. аЕКНБ ю.а., нЦЮПЙНБ о.х. гННМНГМШИ ОРХВХИ ЦПХОО. оПНЦМНГШ ОЮМДЕЛХХ Х ПЕЮКЭМНЯРЭ // фСПМ. ЛХЙПНАХНК. - 2008.
- ╧ 1. - я. 90-95.
10. цЕМДНМ ч.г. оЮМДЕЛХЪ ЦПХООЮ: ОПЕДОНКНФЕМХЪ Х ТЮЙРШ // фСПМ. ЛХЙПНАХНК. - 2008. - ╧ 5. - я. 109-118.
11. цКНАЮКЭМШИ ОКЮМ бнг ОН ОНДЦНРНБЙЕ Й АНПЭАЕ Я ЦПХО╜ОНЛ. - WHO/CDS/CSR/GIP/2005/5/.
12. вЕПЙЮЯЯЙХИ а.к. цКНАЮКЭМЮЪ ЩОХДЕЛХНКНЦХЪ. - л., 2008.
- 447 Я.
13. цЕМДНМ ч.г. яБХМНИ ЦПХОО H1N1 / йЮКХТНПМХЪ - ЯРПЮ╜ЯРХ Х ТЮЙРШ // фСПМ. ЛХЙПНАХНК. - 2010. - ╧ 34. - я. 105-114.
14. йХЯЕКЕБ н.х., йНЛХЯЯЮПНБ ю.а., яРСЙНБЮ л.ю. Х ДП. оЮМДЕЛХВЕЯЙХИ ЦПХОО 2009 Ц. Б пНЯЯХХ. дХЮЦМНЯРХЙЮ Х ЛНКЕЙСКЪП╜МН-АХНКНЦХВЕЯЙХЕ УЮПЮЙРЕПХЯРХЙХ БХПСЯЮ // бНОП. БХПСЯНК. - 2011.
- ╧ 1. - я. 17-21.
15. кЭБНБ д.й., аСПЖЕБЮ е.х., ыЕКЙЮМНБ л.ч. Х ДП. пЮЯОПНЯРПЮМЕМХЕ МНБНЦН ОЮМДЕЛХВЕЯЙНЦН БХПСЯЮ ЦПХООЮ A(H1N1)v Б пНЯЯХХ // бНОП. БХПСЯНК. - 2010. - ╧ 3. - я. 4-9.
16. кЭБНБ д.й., аСПЖЕБЮ е.х., кЮБПХЫЕБЮ б.б. хМТНПЛЮЖХЪ жЕМРПЮ ЩЙНКНЦХХ Х ЩОХДЕЛХНКНЦХХ ЦПХООЮ хМЯРХРСРЮ БХПСЯНКНЦХХ ХЛ. д.х. хБЮМНБЯЙНЦН пюлм НА ХРНЦЮУ ЩОХДЕЛХВЕЯЙНЦН ЯЕГНМЮ 2009-2010 ЦЦ. ОН ЦПХООС Х нпбх (Я 40-И МЕДЕКХ 2009 Ц. ОН 22-Ч МЕДЕКЧ 2010 Ц.) Б ЛХПЕ Х пНЯЯХХ // бНОП. БХПСЯНК. - 2011. - ╧ 1.
- я. 44-49.
17. нМХЫЕМЙН ц.ц. щОХДЕЛХВЕЯЙЮЪ ЯХРСЮЖХЪ ОН ЦПХООС, БШГ╜БЮММНЛС БШЯНЙНОЮРНЦЕММШЛ БХПСЯНЛ РХОЮ A(H1N1), Б пНЯЯХИЯЙНИ тЕДЕПЮЖХХ Х Б ЛХПЕ // фСПМ. ЛХЙПНАХНК. - 2010. - ╧ 1. - я. 3-9.
18. нЯРЕПУЮСЯ щ., нОЕМЬНС о., лНМРН ю. цПХОО // ESWI.
- юМРБЕПОЕМ, аЕКЭЦХЪ, 2010. - 19 Я.
19. пНЛЮМНБЯЙХИ ю.ю., хКЭХВЕБЮ р.м., дСПШЛЮМНБ ю.ц. Х ДП. оЮМДЕЛХВЕЯЙХИ БХПСЯ ЦПХООЮ ю (H1N1) Б юЛСПЯЙНИ НАКЮЯРХ НЯЕ╜МЭЧ 2009 Ц. // фСПМ. ЛХЙПНАХНК. - 2011. - ╧ 1. - я. 35-39.
20. ыЕКЙЮМНБ л.ч., кЭБНБ д.м., тЕДЪЙХМЮ х.р. Х ДП. дХМЮЛХЙЮ ПЮЯОПНЯРПЮМЕМХЪ ОЮМДЕЛХВЕЯЙНЦН ЦПХООЮ A/H1N1 swl МЮ дЮКЭМЕЛ бНЯРНЙЕ Б 2009 Ц. // бНОП. БХПСЯНК. - 2010. - ╧ 3.
- я. 10-15.
21. гСЕБЮ к.о., ъТЮЕБ п.у. щОХДЕЛХНКНЦХЪ: СВЕАМХЙ. - яоА., 2006. - я. 325-342.
22. оНЙПНБЯЙХИ б.х., нМХЫЕМЙН ц.ц., вЕПЙЮЯЯЙХИ а.к. щБНКЧЖХЪ ХМТЕЙЖХНММШУ АНКЕГМЕИ Б пНЯЯХХ Б XX БЕЙЕ. - л., 2003.- я. 184-214.
23. яЕЛЕМНБ а.т., оНЙПНБЯЙХИ б.х. бЮЙЖХМНОПНТХКЮЙРХЙЮ ХМТЮПЙРЮ, ХМЯСКЭРЮ Х КЕРЮКЭМНЯРХ ОПХ ЩОХДЕЛХНКНЦХВЕЯЙНЛ ОНДЗ╜ЕЛЕ ЦПХООЮ // фСПМ. ЛХЙПНАХНК. - 2004. - ╧ 2. - я. 95-99.
24. яНЙНКНБ л.х. нЯРПШЕ ПЕЯОХПЮРНПМШЕ БХПСЯМШЕ ХМТЕЙ╜ЖХХ. - л., 1968. - 259 Я.
25. яНКНБЭЕБ б.д. цПХОО Х ДПСЦХЕ ЯУНДМШЕ ГЮАНКЕБЮМХЪ // нАЫЮЪ Х ВЮЯРМЮЪ ЩОХДЕЛХНКНЦХЪ: ПСЙНБНДЯРБН ДКЪ БПЮВЕИ / ОНД ПЕД. х.х. еКЙХМЮ. - л., 1973. - р 2- я. 3-19.
26. яНЛМХМЮ ю.ю, жШАЮКНБЮ к.л., йЮПОНБЮ к.я. Х ДП. цЕМЕРХВЕЯЙЮЪ ОПЕДПЮЯОНКНФЕММНЯРЭ ДЕРЕИ Я ЦПСООНИ ЙПНБХ (EIII) Й КЮРЕМРМНЛС МНЯХРЕКЭЯРБС БХПСЯЮ ЦПХООЮ ю - БНГЛНФМЮЪ ОПХВХМЮ БНГМХЙМНБЕМХЪ МНБШУ ЩОХДЕЛХВЕЯЙХУ ЬРЮЛЛНБ Б ЯРПЮМЮУ чЦН╜бНЯРНВМНИ юГХХ // бЕЯРМ. пюлм. - 1994. - ╧ 9. - я. 21-24.
27. йХКАСПМ щ. щОХДЕЛХНКНЦХЪ ЦПХООЮ. - л., 1978.
28. фДЮМНБ б.л. щБНКЧЖХЪ БХПСЯНБ. - л., 1990. - 373 Я.
29. яНОПСМНБ т.т. лНКЕЙСКЪПМШЕ НЯМНБШ ОЮПЮГХРХГЛЮ.
- л., 1986. - 173 Я.
30. Garten R., Davis C., Russel C. et al. Antigenic and genetic characteristics of swine-origin 2009 (H1N1) jnfluenza virus circulating in humans // Science. - 2009. - Vol. 325. - P. 197-201.
31. аПХЙН м.х., оНЙПНБЯЙХИ б.х. цКНАЮКХГЮЖХЪ Х ЩОХДЕЛХ╜ВЕЯЙХИ ОПНЖЕЯЯ // щОХДЕЛХНК. Х ХМТЕЙЖ. АНК. - 2010. - ╧ 4. - я. 4-8.
32. Simonsen L., Clarke M., Schonberger L. et al. Pandemic versus influenza mortality: a pattern of changing age distribution // J. Infect. Dis. - 1998. - Vol. 178. - P. 53-60.
33. Scholtissek C., von Hoyninger V., Rott R. Genetic relatedness between the new 1977 epidemic strains (H1N1) isclated between 1947 and 1957 (H1N1) // Virology. - 1978. - Vol. 89.
- P. 613-617.
34. ъЖШЬХМЮ я.а., лХМЕМЙН, ю.м., бНКНЬХМЮ о.б. Х ДП. лНКЕЙСКЪПМН-ЦЕМЕРХВЕЯЙЮЪ УЮПЮЙРЕПХЯРХЙЮ БХПСЯНБ ОЮМДЕЛХВЕ╜ЯЙНЦН ЦПХООЮ A/H1N1(swl 2009), ХГНКХПНБЮММНЦН Б 2009-2010 ЦЦ. Б пНЯЯХХ // фСПМ. ЛХЙПНАХНК. - 2011. - ╧ 1. - я. 26-34.
35. пСАХЯ к.б., лЮЯЧЙ б.я., уСПЖХКЮБЮ н.ц., ыЕПАН ю.о. оПХПНДМШЕ Х ЯНЖХЮКЭМШЕ ТЮЙРНПШ ПХЯЙЮ ГЮАНКЕБЮЕЛНЯРХ НЯРПШ╜ЛХ ПЕЯОХПЮРНПМШЛХ ХМТЕЙЖХЪЛХ // щОХДЕЛХНК. Х ХМТЕЙЖ. АНК.
- 2008. - ╧ 5. - я. 34-37.
36. нА ЩОХДЯХРСЮЖХХ ОН ГЮАНКЕБЮЕЛНЯРХ нпбх Х ЦПХООНЛ Б ЛХПЕ Х пт ГЮ 4-8 МЕДЕКХ 2011 Ц. - http://rospotrebnadzor.ru
37. оНЙПНБЯЙХИ б.х., аПХЙН м.х., пЪОХЯ к.ю. яРПЕОРНЙНЙЙХ Х ЯРПЕОРНЙНЙЙНГШ. - л.: цщнрюп-лЕДХЮ, 2006. - 544.
38. ъТЮЕБ п.у., дСДЮПЕБЮ б.б., хБЮМНБЮ р.ц., йПЮИМНБЮ р.х. щТТЕЙРХБМНЯРЭ ЯХЯРЕЛШ ЩОХДЕЛХНКНЦХВЕЯЙНЦН МЮДГНПЮ ГЮ ЛЕМХМ╜ЦНЙНЙЙНБНИ ХМТЕЙЖХЕИ Б яЮМЙР-оЕРЕПАСПЦЕ // бЕЯРМ. пНЯ. БНЕМ.- ЛЕД. ЮЙЮДЕЛХХ. - 2008. - ╧ 2 (22), ОПХКНФЕМХЕ. - я. 381.
39. яЕПЦХЕБ б.о. аНКЕГМХ ВЕКНБЕЙЮ ЙЮЙ НРПЮФЕМХЕ ЛЕФБХДН╜БНИ АНПЭАШ // фСПМ. ЛХЙПНАХНК. - 2007. - ╧ 3. - я. 97-102.
40. вЕПМНЫЕЙНБ й.ю. щБНКЧЖХЪ ЩРХНКНЦХВЕЯЙНИ ЯРПСЙРСПШ ЬХЦЕККЕГНБ // фСПМ. ЛХЙПНАХНК. - 2010. - ╧ 3. - я. 114-118.
41. ъЙНБКЕБ ю.ю., оНГДЕЕБЮ е.я. хМРЕЦПЮЖХНММЮЪ ЩОХДЕ╜ЛХНКНЦХЪ ЦЕОЮРХРНБ б Х я Б оПХЛНПЯЙНЛ ЙПЮЕ. - бКЮДХБНЯРНЙ: лЕДХЖХМЮ дб, 2011. - 116 Я.
42. кЮОРЕБЮ м.х. хМРЕЦПЮЖХНММЮЪ ЩОХДЕЛХНКНЦХЪ ХМТЕЙ╜ЖХИ Я ЦЕЛНЙНМРЮЙРМШЛ ЛЕУЮМХГЛНЛ ОЕПЕДЮВХ (бхв, ЦЕОЮРХРШ б Х я) Б пЕЯОСАКХЙЕ яЮУЮ (ъЙСРХЪ): ЮБРНПЕТ. ДХЯ.. ЙЮМД. ЛЕД. МЮСЙ.
- нЛЯЙ, 2014. - 26 Я.
43. ъЙНБКЕБ ю.ю., аЮПЮМНБ м.х. рЕНПЕРХВЕЯЙХЕ ЮЯОЕЙРШ ЩОХ╜ДЕЛХНКНЦХВЕЯЙНИ НЖЕМЙХ ОЮМДЕЛХХ ЦПХООЮ A(H1N1) Б 2009-2010 ЦЦ. // щОХДЕЛХНК. Х ХМТЕЙЖ. АНК. - 2012. - ╧ 3. - я. 10-17.
44. сЛЮМЯЙХИ й.ц. оПЕГСЛОЖХЪ МЕБХМНБМНЯРХ БХПСЯНБ // уХЛХЪ Х ФХГМЭ. - 1979. - ╧ 3. - я. 76.
45. ъЦНДХМЯЙХИ б.м. аСДСЫЕЕ ДПЕБМЕИ МЮСЙХ. - л.: гМЮМХЕ, 1982. - 176 Я.
46. оНЙПНБЯЙХИ б.х., вЕПЙЮЯЯЙХИ а.к., яНКНДНБМХЙНБ ч.о. щОХДЕЛХВЕЯЙХИ ОПНЖЕЯЯ Б ЩОНУС МЮСВМН-РЕУМХВЕЯЙНЦН ОПНЦПЕЯЯЮ // пСЙНБНДЯРБН ОН ЩОХДЕЛХНКНЦХХ ХМТЕЙЖХНММШУ АНКЕГМЕИ / ОНД ПЕД. б.х. оНЙПНБЯЙНЦН. - л., 1993. - р 1: бНОПНЯШ НАЫЕИ ЩОХДЕ╜ЛХНКНЦХХ. - я. 25-37.
еЫЕ ОН РЕЛЕ цкюбю 10 ренперхвеяйхе юяоейрш щохделхнкнцхвеяйни нжемйх оюмделхх цпхоою A(H1N1)pdm09 2009-2010 ЦЦ.:
- рЕНПХХ БНГМХЙМНБЕМХЪ ОЮМДЕЛХИ ЦПХООЮ Х ОРХВХИ ЦПХОО.
- яСОНРМХЖЙХИ л. б.. щБНКЧЖХНММЮЪ ОЮРНКНЦХЪ. й БНОПНЯС Н ЛЕЯРЕ бхв-ХМТЕЙЖХХ Х бхв/яохд-ОЮМДЕЛХХ ЯПЕДХ ДПСЦХУ ХМТЕЙЖХНММШУ, ЩОХДЕЛХВЕЯЙХУ Х ОЮМДЕЛХВЕЯЙХУ ОПНЖЕЯЯНБ: ЛНМНЦПЮТХЪ / л. б, яСОНРМХЖЙХИ. ≈ M.: бСГНБЯЙЮЪ ЙМХЦЮ,2009. ≈ 400 Я., 2009
- цКЮБЮ 1. РЕНПЕРХВЕЯЙХЕ ЮЯОЕЙРШ тСМЙЖХНМХПНБЮМХЪ ПШМЙЮ ЛЕДХЖХМЯЙХУ СЯКСЦ
- рЕНПЕРХВЕЯЙХЕ Х ОПХЙКЮДМШЕ ЮЯОЕЙРШ ХГСВЕМХЪ ЯНБКЮДЮЧЫЕЦН ОНБЕДЕМХЪ Б ЯНБПЕЛЕММНИ ОЯХУНКНЦХХ
- щОХДЕЛХНКНЦХВЕЯЙХЕ ЮЯОЕЙРШ бхв-ХМТЕЙЖХХ
- йКХМХЙН-ЩОХДЕЛХНКНЦХВЕЯЙХЕ ЮЯОЕЙРШ ТСМЙЖХНМЮКЭМШУ ЙХЯР
- яНЖХЮКЭМН-ЦХЦХЕМХВЕЯЙХЕ Х ЩОХДЕЛХНКНЦХВЕЯЙХЕ ЮЯОЕЙРШ ЯХТХКХЯЮ
- 1.щОХДЕЛХНКНЦХВЕЯЙХЕ ЮЯОЕЙРШ бхв-ХМТЕЙЖХХ Х РСАЕПЙСКЕГЮ
- 1.4 щОХДЕЛХНКНЦХВЕЯЙХЕ ЮЯОЕЙРШ ХЬЕЛХВЕЯЙНИ АНКЕГМХ ЯЕПДЖЮ Х ЮРЕПНЯЙКЕПНГЮ
- ьКНОНБ б.ц.. оЮРНКНЦХВЕЯЙЮЪ ЮМЮРНЛХЪ: сВЕАМХЙ. ≈ дНМЕЖЙ: йЮЬРЮМ,2010. ≈ 472 Я., 2010
- нЖЕМЙЮ ЩОХДЕЛХНКНЦХВЕЯЙНИ НАЯРЮМНБЙХ ОН УНКЕПЕ Б пНЯЯХХ
- аСГСМНБ ю. т.. тНПЛХПНБЮМХЕ ЯНЛЮРХВЕЯЙХУ ОНЯКЕДЯРБХИ ЮДЮОРЮЖХНММНЦН ЯХМДПНЛЮ. жЕМЮ ЖХБХКХГЮЖХХ. ≈ л.: оПЮЙРХВЕЯЙЮЪ ЛЕДХЖХМЮ,2010. - 352 Я., 2010
- юМДПЕЕБЮ р.б.. яНГДЮМХЕ ОПЕГЕМРЮЖХИ Б Microsoft Power Point // сВЕАМНЕ ОНЯНАХЕ. - л.,2009. - 104 Я., 2009
- б.б. йЮГЮЙЕБХВ. бНЕММН-ОНКЕБЮЪ РЕПЮОХЪ Х ЦЕЛЮРНКНЦХЪ. хГДЮРЕКЭЯРБН цнс бон дЮКЭМЕБНЯРНВМШИ ЦНЯСДЮПЯРБЕММШИ ЛЕДХЖХМЯЙХИ СМХБЕПЯХРЕР - 2010, 2010
- хМДХБХДСЮКХГЮЖХЪ КЕЙЮПЯРБЕММНИ РЕПЮОХХ АНКЭМШУ ЮПРЕПХЮКЭМНИ ЦХОЕПРЕМ╜ГХЕИ : СВЕА.-ЛЕРНД. ОНЯНАХЕ / е. я. аЕКЭЯЙЮЪ. - лХМЯЙ : ацлс,2010. - 36 Я., 2010
- б.ъ. цЮПЛЮЬ, н.л. сПЪЯЭЕБ, й.б.рПСТЮМНБ. цхоепрнмхвеяйюъ анкегмэ. сВЕАМНЕ ОНЯНАХЕ ДКЪ ЯРСДЕМРНБ КЕВЕАМНЦН ТЮЙСКЭРЕРЮ. - пЪГЮМЭ, 2010, 2010
- хЯЙЕМДЕПНБ а.ц.. щКЕЙРПХВЕЯЙЮЪ МЕЯРЮАХКЭМНЯРЭ ЯЕПДЖЮ ОПХ ЮПРЕПХЮКЭМНИ ЦХОЕПРЕМГХХ: лНМНЦПЮТХЪ. - оЕМГЮ,2009. - 208Я.: ХК., 2009
- вЕПМНБ м.м.. лЕРНДШ Х ЯПЕДЯРБЮ ЮМЮКХРХВЕЯЙНЦН ЙНМРПНКЪ: сВЕАМНЕ ОНЯНАХЕ. √ рЮЦЮМПНЦ: хГД-БН ррх чтс,2009. √ 60 Я., 2009
- б.т. дЮМХКХВЕБ. яНБПЕЛЕММЮЪ НТРЮКЭЛНКНЦХЪ: пСЙНБНДЯРБН. 2-Е ХГД. / оНД ПЕД. б.т. дЮМХКХВЕБЮ. ≈ яоА.: оХРЕП,2009. ≈ 688 Я.: ХК., 2009