вХЯКН АКНУ, ОХРЮЧЫХУЯЪ МЮ НДМНЛ ОПНЙНПЛХРЕКЕ Б ОЕПХНД АЮЙРЕПХЕЛХХ
оПЕДБЮПХРЕКЭМЮЪ НЖЕМЙЮ ЮМЮКХГХПСЕЛНЦН Б ДЮММНЛ ОНДПЮГДЕКЕ ОНЙЮГЮРЕКЪ ОПЕДОНКЮЦЮЕР ЕЦН ГЮБХЯХЛНЯРЭ, ОПЕФДЕ БЯЕЦН, НР ВХЯКЕММНЯРХ АКНУ Х НР ОПНДНКФХРЕКЭМНЯРХ АЮЙРЕПХЕЛХХ С УНГЪЕБ-МНЯХРЕКЕИ БНГАСДХРЕКЪ ВСЛШ.
нВЕБХДМН РЮЙФЕ МЮКХВХЕ ГЮБХЯХЛНЯРХ Х НР ОЮПЮЛЕРПНБ ОЮПЮГХРХПНБЮМХЪ АКНУ ≈ ОПНДНКФХРЕКЭМНЯРХ НДМНЙПЮРМНЦН ОПЕАШБЮМХЪ Б ЬЕПЯРХ Х ВЮЯРНРШ МЮОЮДЕМХИ МЮ ОПНЙНПЛХРЕКЪ. жЕКЕЯННАПЮГМШЛ ОПЕДЯРЮБКЪЕРЯЪ РЕНПЕРХВЕЯЙНЕ НОПЕДЕКЕМХЕ ГЮБХЯХЛНЯРХ ВХЯКЮ ОХРЮЧЫХУЯЪ Б ЕДХМХЖС БПЕЛЕМХ АКНУ НР СОНЛЪМСРШУ БШЬЕ ОЮПЮЛЕРПНБ. мХФЕ ЛНФМН АСДЕР САЕДХРЭЯЪ, ВРН ГМЮМХЕ ЩРНИ ГЮБХЯХЛНЯРХ НРЙПШБЮЕР ОЕПЯОЕЙРХБС АНКЕЕ ОПНЯРН НЖЕМХБЮРЭ ВХЯКН ЩЙРНОЮПЮГХРНБ, ОХРЮЧЫХУЯЪ МЮ ЦПШГСМЕ ГЮ КЧАНИ НРПЕГНЙ БПЕЛЕМХ.дКЪ НОПЕДЕКЕМХЪ ХЯЙНЛНИ ГЮБХЯХЛНЯРХ АШКН ОПХЛЕМЕМН ЦПЮТХВЕЯЙНЕ ЛНДЕКХПНБЮМХЕ Я ХЯОНКЭГНБЮМХЕЛ УПНМНЦПЮЛЛ ОЮПЮГХРХПНБЮМХЪ АКНУ (йСГМЕЖНБ, 1997 Ю). оПХ ЩРНЛ МЮЛ РПЕАНБЮКНЯЭ ГМЮМХЕ НАЫЕИ ВХЯКЕММНЯРХ АКНУ Б МЕЙНРНПНЛ ЛХЙПНАХНРНОЕ, ОНЯЕКЕМХХ ХКХ ОНОСКЪЖХХ ЦПШГСМНБ- УНГЪЕБ, ЕЯКХ ЙЮЙНЕ-КХАН ХГ ЩРХУ НАПЮГНБЮМХИ ОПХГМЮЕРЯЪ ЙЮЙ ЖЕКНЯРМЮЪ Х ГЮЛЙМСРЮЪ (ОН СЯКНБХЧ ОЕПБХВМНИ ЛНДЕКХ) ЯХЯРЕЛЮ, Х ВХЯКЮ НАХРЮЧЫХУ Б ЩРНИ ЯХЯРЕЛЕ ЦПШГСМНБ.
хГБЕЯРМН, ВРН Б ЙЮФДШИ ЛНЛЕМР БПЕЛЕМХ КХЬЭ МЕЙНРНПЮЪ ВЮЯРЭ ЙПНБНЯНЯНБ МЮУНДХРЯЪ Б ЬЕПЯРХ ЦПШГСМНБ, Ю БЯЕ НЯРЮКЭМШЕ ≈ Б ЦМЕГДЮУ ХКХ ОН УНДЮЛ МНП. вЕПЕГ МЕЙНРНПНЕ БПЕЛЪ РН ХКХ ХМНЕ ЙНКХВЕЯРБН ОЮПЮГХРНБ ОНЙХДЮЕР ОПНЙНПЛХРЕКЕИ, Ю ХУ ЛЕЯРН ОНЯРЕОЕММН ГЮМХЛЮЧР ДПСЦХЕ, ОПНЦНКНДЮБЬХЕЯЪ Й РНЛС ЛНЛЕМРС. оПНЖЕЯЯ ЯЛЕМШ НДМХУ ОЮПЮГХРХПСЧЫХУ НЯНАЕИ ДПСЦХЛХ ХДЕР МЕОПЕПШБМН. еЯРЕЯРБЕММН, ВРН Б ПЮГМШЕ ЛНЛЕМРШ МЮОЮДЮРЭ МЮ ЦПШГСМНБ Х ОНЙХДЮРЭ ХУ, Ю, ЯКЕДНБЮРЕКЭМН, Х ПЕЦХЯРПХПНБЮРЭЯЪ Б ЬЕПЯРХ АСДЕР ПЮГМНЕ ВХЯКН АКНУ. мН ОПХ ДНЯРЮРНВМН ДКХРЕКЭМНЛ ПЪДЕ МЮАКЧДЕМХИ ЯПЕДМЕЕ ЙНКХВЕЯРБН НДМНЛНЛЕМРМН ОЮПЮГХРХПСЧЫХУ МЮЯЕЙНЛШУ АСДЕР ОПХАКХФЮРЭЯЪ Й МЕЙНРНПНЛС ЯПЕДМЕЛС ГМЮВЕМХЧ,
184 УЮПЮЙРЕПХГСЧЫЕЛС ХЯЙНЛСЧ ГЮБХЯХЛНЯРЭ. щРН ОНКНФЕМХЕ, ЙНЦДЮ МЕ БЯЕ, Ю КХЬЭ НОПЕДЕКЕММЮЪ ВЮЯРЭ АКНУ МЮУНДХРЯЪ Б ЬЕПЯРХ ОПНЙНПЛХРЕКЕИ, МЮЬКН НРПЮФЕМХЕ Б ХМДЕЙЯЕ ОПХСПНВЕММНЯРХ ЩРХУ МЮЯЕЙНЛШУ Й ЯБНЕЛС УНГЪХМС (хНТТ, 1941; фНБРШИ, 1960; 1966; мНБНЙПЕЫЕМНБЮ, 1960; аЕЙКЕЛХЬЕБ, 1961).
вРНАШ ОНМЪРЭ ГЮЙНМНЛЕПМНЯРХ, НАЕЯОЕВХБЮЧЫХЕ ПЕЮКЭМНЯРЭ ЯСЫЕЯРБНБЮМХЪ ХМДЕЙЯЮ ОПХСПНВЕММНЯРХ, АШКХ ОНЯРПНЕМШ УПНМНЦПЮЛЛШ ОЮПЮГХРХПНБЮМХЪ АКНУ СЯКНБМНЦН (ЦХОНРЕРХВЕЯЙНЦН) БХДЮ, ЯХЯОНКЭГНБЮМХЕЛ ОПНДНКФХРЕКЭМНЯРХ НДМНЙПЮРМНЦН (МЕОПЕПШБМНЦН)
ОЮПЮГХРХПНБЮМХЪ Х НАЫЕИ ДКХРЕКЭМНЯРХ ХУ РПНТХВЕЯЙНЦН ЖХЙКЮ Я ЛНЛЕМРЮ НДМНЦН МЮОЮДЕМХЪ МЮ УНГЪХМЮ ДН ЛНЛЕМРЮ ЯКЕДСЧЫЕЦН МЮОЮДЕМХЪ.
мЮ ОЕПБШУ ЩРЮОЮУ ХЯЯКЕДНБЮМХЪ ЯРПНХКХ УПНМНЦПЮЛЛШ Я НДХМЮЙНБШЛХ ОЮПЮЛЕРПЮЛХ ОЮПЮГХРХПНБЮМХЪ БЯЕУ АКНУ, ГЮДЕИЯРБНБЮММШУ Б НДМНИ ЯХЯРЕЛЕ. нДМХЛ ХГ СЯКНБХИ ОПНЯРЕИЬЕИ ЛНДЕКХ ОПХМЪКХ, ВРН НВЕПЕДМНЯРЭ ХУ МЮОЮДЕМХИ МЮ ЦПШГСМНБ НЯСЫЕЯРБКЪЕРЯЪ ВЕПЕГ НДХМЮЙНБШЕ ОПНЛЕФСРЙХ БПЕЛЕМХ. б НДМНИ ХГ ЛНДЕКЕИ СЯРЮМНБХКХ ОПНДНКФХРЕКЭМНЯРЭ РПНТХВЕЯЙНЦН ЖХЙКЮ ЩЙРНОЮПЮГХРНБ, ПЮБМСЧ 24 В, Ю ОПНДНКФХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОПЕАШБЮМХЪ Б ЬЕПЯРХ ≈ 3 В. дНОНКМХРЕКЭМШЛ СЯКНБХЕЛ ЛНДЕКХ ЪБКЪКЯЪ РЮЙНИ ПЕФХЛ ОПЕАШБЮМХЪ ЦПШГСМНБ Б МНПЮУ, ЙНРНПШИ МЕ ? БМНЯХК ОНЛЕУ Б ОПНЖЕЯЯ МЮОЮДЕМХИ АКНУ. вХЯКН ОЮПЮГХРНБ Б ЩРНЛ ЛХЙПНАХНРНОЕ АШКН ОПХМЪРН ПЮБМШЛ 16, Ю ПЮБМНЛЕПМЮЪ НВЕПЕДМНЯРЭ МЮОЮДЕМХИ НАЕЯОЕВХБЮКЮЯЭ ОПХ ОНКСРНПЮВЮЯНБНЛ ХМРЕПБЮКЕ ЛЕФДС МХЛХ.
цПЮТХВЕЯЙСЧ ЛНДЕКЭ ЯРПНХКХ ЯКЕДСЧЫХЛ НАПЮГНЛ ≈ МЮ НАЫЕИ ЬЙЮКЕ БПЕЛЕМХ ЙЮФДНИ АКНУЕ АШКЮ БШДЕКЕМЮ НРДЕКЭМЮЪ ЦНПХГНМРЮКЭМЮЪ ОНКНЯЮ (ПХЯСМНЙ 16). C МЕЙНРНПНЦН ОПНХГБНКЭМН БШАПЮММНЦН ЛНЛЕМРЮ БПЕЛЕМХ ОЕПБСЧ АКНУС (МХФМЪЪ ОНКНЯЮ) ╚ГЮЯРЮБКЪКХ╩ МЮОЮДЮРЭ МЮ ОПНЙНПЛХРЕКЪ Х БПЕЛЪ ОПЕАШБЮМХЪ ЕЕ Б ЬЕПЯРХ (гВ) ГЮЙПЮЬХБЮКХ. гЮРЕЛ ЯКЕДНБЮК МЕГЮЙПЮЬЕММШИ СВЮЯРНЙ ОНКНЯШ (21В), ЙНЦДЮ АКНУЮ МЮУНДХКЮЯЭ Б ЦМЕГДЕ ХКХ МНПЕ, ОНЯКЕ ВЕЦН НОЪРЭ ЯКЕДНБЮКН МЮОЮДЕМХЕ Х Р. Д. йЮФДНЕ
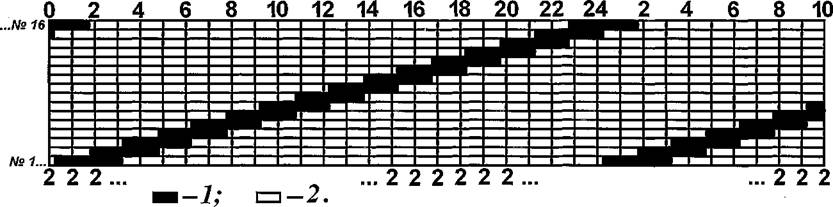
пХЯ. 16. уПНМНЦПЮЛЛЮ ПЮБМНЛЕПМНИ НВЕПЕДМНЯРХ МЮОЮДЕМХИ АКНУ (╧╧ 1-16) МЮ ЦПШГСМНБ Б ЛНДЕКЭМНЛ ОНЯЕКЕМХХ. вХЯКЮ: ЯБЕПУС ≈ БПЕЛЪ ЯСРНЙ (ВЮЯШ), ЯМХГС ≈ ЙНКХВЕЯРБН АКНУ, МЮУНДЪЫХУЯЪ Б ЬЕПЯРХ ЦПШГСМНБ Б ПЮГКХВМШЕ ЛНЛЕМРШ БПЕЛЕМХ; 1 ОЕПХНДШ МЮУНФДЕМХЪ ЙЮФДНИ АКНУХ Б ЬЕПЯРХ; 2 ≈ ОПНЛЕФСРЙХ ЛЕФДС ОЕПХНДЮЛХ ОЮПЮГХРХПНБЮМХЪ АКНУХ.
OO
IZi
186 НВЕПЕДМНЕ МЮОЮДЕМХЕ БРНПНИ АКНУХ ЯКЕДНБЮКН ВЕПЕГ 1,5 В ОНЯКЕ МЮОЮДЕМХЪ ОЕПБНИ, РПЕРЭЕИ ≈ ОНЯКЕ БРНПНИ Х Р.
Д.еЯКХ ОПНБЕЯРХ МЮ КЧАНЛ СВЮЯРЙЕ ЛНДЕКХ ОПНХГБНКЭМНЕ ЙНКХВЕЯРБН БЕПРХЙЮКЭМШУ КХМХИ, ЯННРБЕРЯРБСЧЫХУ ПЮГКХВМШЛ ЛНЛЕМРЮЛ БПЕЛЕМХ, РН БХДМН, ВРН НДМНЛНЛЕМРМН ОЮПЮГХРХПСЧР БЯЕЦДЮ РНКЭЙН 2 АКНУХ. б ЯННРБЕРЯРБХХ ЯН ЯЛШЯКНБШЛ ЯНДЕПФЮМХЕЛ ХМДЕЙЯЮ ОПХСПНВЕММНЯРХ АКНУ Й УНГЪХМС Б ДЮММНИ ЛНДЕКХ НМ ПЮБЕМ 2/16 == 0,125 ХКХ 12,5%.
кНЦХВМН ОПЕДОНКНФХРЭ, ВРН ВХЯКН АКНУ, МЮУНДЪЫХУЯЪ Б ЬЕПЯРХ ЦПШГСМНБ Б ЯПЕДМЕЯРЮРХЯРХВЕЯЙХИ ЛНЛЕМР БПЕЛЕМХ, НРМНЯХРЯЪ Й ХУ НАЫЕЛС ВХЯКС РЮЙ ФЕ, ЙЮЙ ДКХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ ЙН БЯЕЛС РПНТХВЕЯЙНЛС ЖХЙКС. сОНЛХМЮМХЪ Н ОНДНАМНИ ГЮБХЯХЛНЯРХ ЛНФМН БЯРПЕРХРЭ Х Б АНКЕЕ ПЮММХУ ПЮАНРЮУ (хНТТ, 1941; цЮСГЬРЕИМ Я ЯНЮБР., 1965; яНКДЮРЙХМ Я ЯНЮБР., 1967), МН АЕГ ЛЮРЕЛЮРХВЕЯЙНИ ХМРЕПОПЕРЮЖХХ, ОНГБНКЪЧЫЕИ ОН-ХМНЛС ОНДНИРХ Й ПЕЬЕМХЧ ГЮДЮВХ Н ВХЯКЕ АКНУ, ГЮПЮФЮЧЫХУЯЪ МЮ НДМНЛ АНКЭМНЛ ВСЛНИ ЦПШГСМЕ.
б ЯКСВЮЕ ПЮБМНЛЕПМНИ НВЕПЕДМНЯРХ МЮОЮДЕМХИ АКНУ СЙЮГЮММЮЪ ГЮБХЯХЛНЯРЭ НАМЮПСФХБЮЕРЯЪ БЕЯЭЛЮ ОПНЯРН Х МЮЦКЪДМН ≈ 2/16 = 3/24, ОНЩРНЛС Б НОПЕДЕКЕМХЕ ХМДЕЙЯЮ ОПХСПНВЕММНЯРХ АКНУ Й УНГЪХМС ЛНФМН БЙКЧВХРЭ ЯННРМНЬЕМХЕ УПНМНЛЕРПХВЕЯЙХУ УЮПЮЙРЕПХЯРХЙ ОЮПЮГХРХПНБЮМХЪ:

ЦДЕ f ≈ ХМДЕЙЯ ОПХСПНВЕММНЯРХ АКНУ Й УНГЪХМС, b ≈ ЙНКХВЕЯРБН НДМНЛНЛЕМРМН ОЮПЮГХРХПСЧЫХУ АКНУ, б ≈ НАЫЮЪ ВХЯКЕММНЯРЭ АКНУ Б ЛХЙПНАХНРНОЕ, ta≈ ОПНДНКФХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ АКНУХ, T ≈ НАЫЮЪ ДКХРЕКЭМНЯРЭ РПНТХВЕЯЙНЦН ЖХЙКЮ.
187
хГ ЩРНИ ОПНОНПЖХХ ЯКЕДСЕР:

яПЕДМЕЕ ВХЯКН АКНУ ЮМЮКХГХПСЕЛНЦН БХДЮ, МЮУНДЪЫХУЯЪ Б ЬЕПЯРХ НДМНЦН ЦПШГСМЮ Б МЕЙНРНПШИ ЛНЛЕМР БПЕЛЕМХ, МЮГШБЮЕЛНЕ ХМДЕЙЯНЛ НАХКХЪ (хн) АКНУ Б ЬЕПЯРХ, ПЮБМН:

ЦДЕ Io ≈ ХМДЕЙЯ НАХКХЪ, N ≈ ВХЯКН НАХРЮЧЫХУ Б ЛХЙПНАХНРНОЕ ЦПШГСМНБ.
вРНАШ БШЪЯМХРЭ, АСДЕР КХ ЯНУПЮМЪРЭЯЪ ОНКСВЕММНЕ ЯННРМНЬЕМХЕ (1) ОПХ МЕЯНАКЧДЕМХХ ПЮБМНЛЕПМНИ НВЕПЕДМНЯРХ МЮОЮДЕМХИ АКНУ, ЛШ ПЮЯЯЛНРПЕКХ МЕЯЙНКЭЙН УПНМНЦПЮЛЛ Я ПЮГКХВМНИ ЯРЕОЕМЭЧ МЕПЮБМНЛЕПМНЯРХ.
гДЕЯЭ ЛШ ОПХБНДХЛ НДМС ХГ МХУ (ПХЯСМНЙ 17), МЮ ЙНРНПНИ ОПНДНКФХРЕКЭМНЯРЭ РПНТХВЕЯЙНЦН ЖХЙКЮ ПЮБМЮ 17 В, ОЕПХНД ОЮПЮГХРХПНБЮМХЪ ≈ 4 В, Ю ВХЯКН АКНУ ≈ 10. нВЕПЕДМНЯРЭ МЮОЮДЕМХИ МЕПЮБМНЛЕПМЮЪ Х ЯКСВЮИМЮЪ.вРНАШ БШВХЯКХРЭ ЯПЕДМЕЕ ВХЯКН НДМНЛНЛЕМРМН ОЮПЮГХРХПСЧЫХУ АКНУ МСФМН ПЮЯЯЛНРПЕРЭ БЯЧ ЦЕМЕПЮКЭМСЧ ЯНБНЙСОМНЯРЭ ГМЮВЕМХИ. дКЪ ЕЕ ОНКСВЕМХЪ МЕНАУНДХЛН ЯВХРШБЮМХЕ ХМТНПЛЮЖХХ ВЕПЕГ ПЮБМШЕ ОПНЛЕФСРЙХ БПЕЛЕМХ, МЮОПХЛЕП ВЕПЕГ 1 В. цЕМЕПЮКЭМЮЪ ЯНБНЙСОМНЯРЭ НАПЮГСЕРЯЪ ХГ КЧАНЦН НРПЕГЙЮ ЬЙЮКШ ДКХМНИ Б НДХМ РПНТХВЕЯЙХИ ЖХЙК, Б ЙНРНПНЛ ВХЯКН БЮПХЮМРНБ ОПХ НДМНВЮЯНБНЛ ЬЮЦЕ ПЮБМН 17. яКЕДСЕР ГЮЛЕРХРЭ, ВРН НРПЕГЙХ, ЯЛЕФМШЕ Я БШАПЮММШЛ ДКЪ ЮМЮКХГЮ, ЙЮЙ Х БЯЕ ДПСЦХЕ, ХЛЕЧЫХЕ РЮЙСЧ ФЕ ДКХМС Х ЯЛЕФМШЕ ДПСЦ Я ДПСЦНЛ, ЮАЯНКЧРМН ХДЕМРХВМШ.
яПЕДМЕЕ ВХЯКН НДМНБПЕЛЕММН ОЮПЮГХРХПСЧЫХУ АКНУ (э), БШВХЯКЕММНЕ ХГ 17 ГМЮВЕМХИ, ПЮБМН 2,353. оПХ ОНДЯРЮМНБЙЕ Б ТНПЛСКС (2) ХЯУНДМШУ ОЮПЮЛЕРПНБ, СОНРПЕАКЕММШУ Б ДЮММНИ ЛНДЕКХ, ОНКСВЮЕРЯЪ РН ФЕ
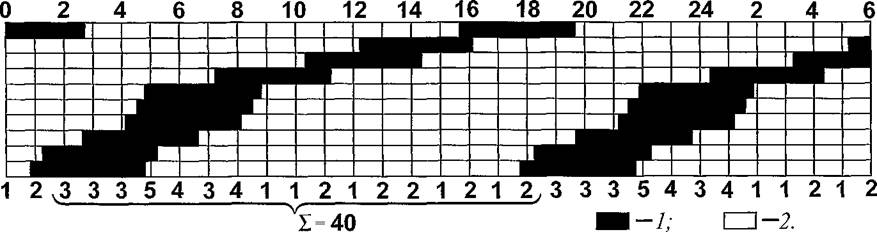
пХЯ. 17. уПНМНЦПЮЛЛЮ МЕПЮБМНЛЕПМНИ НВЕПЕДМНЯРХ МЮОЮДЕМХИ АКНУ МЮ ЦПШГСМНБ Б ЛНДЕКЭМНЛ ОНЯЕКЕМХХ. вХЯКЮ: ЯБЕПУС ≈ БПЕЛЪ ЯСРНЙ (ВЮЯШ), ЯМХГС ≈ ЙНКХВЕЯРБН АКНУ, МЮУНДЪЫХУЯЪ Б ЬЕПЯРХ ЦПШГСМНБ Б ПЮГКХВМШЕ ЛНЛЕМРШ БПЕЛЕМХ; 1 ОЕПХНДШ МЮУНФДЕМХЪ ЙЮФДНИ АКНУХ Б ЬЕПЯРХ; 2 ≈ ОПНЛЕФСРЙХ ЛЕФДС ОЕПХНДЮЛХ ОЮПЮГХРХПНБЮМХЪ
189 ЯЮЛНЕ ГМЮВЕМХЕ. оНЯРПНЕМХЕ УПНМНЦПЮЛЛ Я КЧАШЛХ ДПСЦХЛХ ЙНЛАХМЮЖХЪЛХ ОЮПЮЛЕРПНБ Х ╚ЦПЮТХЙНБ╩ МЮОЮДЕМХИ БЯЕЦДЮ ОПХБНДХКН Й РНВМНЛС ЯНБОЮДЕМХЧ ПЮЯВЕРМНЦН Х ТЮЙРХВЕЯЙНЦН ЯПЕДМЕЦН ВХЯКЮ НДМНБПЕЛЕММН ОЮПЮГХРХПСЧЫХУ АКНУ.
яКЕДСЧЫХЛ ЩРЮОНЛ НРУНДЮ НР ХДЕЮКХГХПНБЮММНИ ЛНДЕКХ АШКН ОНЯРПНЕМХЕ УПНМНЦПЮЛЛ, ЙНЦДЮ ОЮПЮЛЕРПШ ОЮПЮГХРХПНБЮМХЪ С ЙЮФДНИ АКНУХ АШКХ ХМДХБХДСЮКЭМШЛХ. оНДНАМШЕ ТХГХНКНЦХВЕЯЙХЕ УЮПЮЙРЕПХЯРХЙХ Б ЕЯРЕЯРБЕММШУ ЦПСООХПНБЙЮУ ФХБНРМШУ ОНДВХМЪЧРЯЪ ГЮЙНМНЛЕПМНЯРЪЛ МНПЛЮКЭМНЦН ПЮЯОПЕДЕКЕМХЪ, ВРН Х АШКН ОНКНФЕМН Б НЯМНБС ТНПЛХПНБЮМХЪ ЛНДЕКЭМНИ ЦПСООШ МЮЯЕЙНЛШУ. яНВЕРЮМХЕ ЙНМЙПЕРМШУ ГМЮВЕМХИ TХ taДКЪ ЙЮФДНИ АКНУХ Х ЛНЛЕМРШ ХУ ОЕПБНЦН (╚ЯРЮПРНБНЦН╩) МЮОЮДЕМХЪ МЮ ОПНЙНПЛХРЕКЕИ АШКХ НОПЕДЕКЕМШ ЯКСВЮИМН.
б НДМНИ ХГ ЛНДЕКЕИ, ЦДЕ СВХРШБЮЕРЯЪ ОЮПЮГХРХПНБЮМХЕ 12 АКНУ, АШКХ ОПХМЪРШ ЯКЕДСЧЫХЕ ЯПЕДМХЕ ОЮПЮЛЕРПШ: T = 10 В, ta = 2,5 В, ОПХ ЙНКЕАЮМХЪУ НР 9 ДН 11 Х НР 1,5 ДН 3,5 В ЯННРБЕРЯРБЕММН. яПЕДМЕЕ ГМЮВЕМХЕ э, БШВХЯКЕММНЕ ОН ТНПЛСКЕ (2), ПЮБМН 3,0.
уПНМНЦПЮЛЛЮ МЮ ПХЯСМЙЕ 18 АШКЮ ОНЯРПНЕМЮ ОНЯКЕДНБЮРЕКЭМШЛ ПЮЯОХЯШБЮМХЕЛ ЯСДЭАШ ЙЮФДНИ АКНУХ Б ЯННРБЕРЯРБХХ Я ЕЕ ХМДХБХДСЮКЭМШЛХ ОЮПЮЛЕРПЮЛХ ОЮПЮГХРХПНБЮМХЪ. йЮЙ Х ЯКЕДНБЮКН НФХДЮРЭ, АШК ОНКСВЕМ ОНЯРНЪММН ЛЕМЪЧЫХИЯЪ ╚ПХЯСМНЙ╩ МЮОЮДЕМХИ. аКНУХ Я АНКЕЕ ЙНПНРЙХЛХ РПНТХВЕЯЙХЛХ ЖХЙКЮЛХ БЯЕ АНКЕЕ Х АНКЕЕ ╚НАЦНМЪКХ╩ ЛЕДКХРЕКЭМШУ. пЮГАХБ ЬЙЮКС БПЕЛЕМХ МЮ МЕЯЙНКЭЙН НРПЕГЙНБ, ПЮБМШУ ЯПЕДМЕЛС РПНТХВЕЯЙНЛС ЖХЙКС, Х ОПНЯСЛЛХПНБЮБ БЮПХЮМРШ ВХЯЕК ОЮПЮГХРХПНБЮБЬХУ АКНУ ВЕПЕГ ЙЮФДШИ ВЮЯ, ЛНФМН САЕДХРЭЯЪ, ВРН ЯПЕДМЕЕ ВХЯКН ОЮПЮГХРНБ Б ЬЕПЯРХ ДКЪ МЕЙНРНПШУ НРПЕГЙНБ МЕГМЮВХРЕКЭМН НРКХВЮЕРЯЪ НР ПЮЯВЕРМНЦН. оПХ СБЕКХВЕМХХ ВХЯКЮ ЮМЮКХГХПСЕЛШУ НРПЕГЙНБ МЮ ДКХММНИ ЬЙЮКЕ БПЕЛЕМХ НАЫЕЕ ЯПЕДМЕЕ БЯЕ АНКЭЬЕ Х АНКЭЬЕ ОПХАКХФЮЕРЯЪ Й ПЮЯВЕРМНЛС. мН Х ОПХ ЮМЮКХГЕ ДЮФЕ НДМНЦН НРПЕГЙЮ ОПНДНКФХРЕКЭМНЯРЭЧ TНЬХАЙЮ МЮЯРНКЭЙН ЛЮКЮ, ВРН ЕЧ ЛНФМН ОПЕМЕАПЕВЭ.
190

пХЯ. 18. уПНМНЦПЮЛЛЮ МЕПЮБМНЛЕПМНИ НВЕПЕДМНЯРХ МЮОЮДЕМХИ МЮ ЦПШГСМНБ АКНУ Я ПЮГКХВЮЧЫХЛХЯЪ ОЮПЮЛЕРПЮЛХ ОЮПЮГХРХПНБЮМХЪ. ю ≈ МЮВЮКН; а ≈ ОПНДНКФЕМХЕ. вХЯКЮ: ЯБЕПУС ≈ БПЕЛЪ ЯСРНЙ (ВЮЯШ), ЯМХГС ≈ ЙНКХВЕЯРБН АКНУ, МЮУНДЪЫХУЯЪ Б ЬЕПЯРХ ЦПШГСМНБ Б ПЮГКХВМШЕ ЛНЛЕМРШ БПЕЛЕМХ; 1 ≈ ОЕПХНДШ МЮУНФДЕМХЪ ЙЮФДНИ АКНУХ Б ЬЕПЯРХ; 2 ≈ ОПНЛЕФСРЙХ ЛЕФДС ОЕПХНДЮЛХ ОЮПЮГХРХПНБЮМХЪ АКНУХ.
191
б ПЕЮКЭМШУ СЯКНБХЪУ БЮПХЮАЕКЭМНЯРЭ ХМДХБХДСЮКЭМШУ ОЮПЮЛЕРПНБ ОЮПЮГХРХПНБЮМХЪ АКНУ ДЮФЕ НДМНЦН БХДЮ, ОНКЮ Х БНГПЮЯРЮ ЛНФЕР НЙЮГЮРЭЯЪ АНКЕЕ ГМЮВХРЕКЭМНИ, ВЕЛ Б МЮЬЕИ РЕНПЕРХВЕЯЙНИ ЛНДЕКХ. нДМЮЙН БНГМХЙЮЧЫЮЪ ОПХ ЩРНЛ НЬХАЙЮ Б НОПЕДЕКЕМХХ ЯПЕДМЕЦН ГМЮВЕМХЪ з ЖЕКХЙНЛ Х ОНКМНЯРЭЧ НАСЯКЮБКХБЮЕРЯЪ КХЬЭ ЩРНИ БЮПХЮАЕКЭМНЯРЭЧ. яКЕДНБЮРЕКЭМН, НМЮ ЛНФЕР АШРЭ НЖЕМЕМЮ ЯРЮРХЯРХВЕЯЙХЛХ ЛЕРНДЮЛХ Х МЮ ЯЮЛНЛ ДЕКЕ МЕ НЙЮГШБЮЕР МХЙЮЙНЦН БКХЪМХЪ МЮ ХГСВЮЕЛСЧ ГЮЙНМНЛЕПМНЯРЭ.
рЮЙХЛ НАПЮГНЛ, ЯКЕДСЕР ЯВХРЮРЭ ДНЙЮГЮММШЛ ЯСЫЕЯРБНБЮМХЕ ╚ГЮЙНМЮ ОПХСПНВЕММНЯРХ╩ ОЕПХНДХВЕЯЙХУ (БПЕЛЕММШУ) ОЮПЮГХРНБ Й УНГЪХМС Б РНЛ БХДЕ, ЙЮЙ ЩРН ОПЕДЯРЮБКЕМН Б ОПНОНПЖХХ (1). щРН ОНГБНКЪЕР ДЮРЭ МНБСЧ УЮПЮЙРЕПХЯРХЙС РЮЙ МЮГШБЮЕЛШЛ АКНУЮЛ ЬЕПЯРХ Х АКНУЮЛ ЦМЕГДЮ. с ОЕПБШУ ОПНДНКФХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ ЯНЯРЮБКЪЕР АНКЕЕ 50% НР ДКХРЕКЭМНЯРХ РПНТХВЕЯЙНЦН ЖХЙКЮ, Ю С БРНПШУ ≈ ЛЕМЕЕ (йСГМЕЖНБ, 1997 Ю). яНАЯРБЕММН, Х ПЮМЭЬЕ ОНДНАМШИ 50-ОПНЖЕМРМШИ ПСАЕФ ЯКСФХК МЕЦКЮЯМШЛ ЙПХРЕПХЕЛ ДХТТЕПЕМЖХЮЖХХ ПЮГКХВМШУ БХДНБ АКНУ ОН СЙЮГЮММШЛ ЩЙНКНЦХВЕЯЙХЛ ЦПСООЮЛ, РНКЭЙН ХЯУНДЪ МЕ ХГ ДКХРЕКЭМНЯРЕИ ОЕПХНДНБ РПНТХВЕЯЙНЦН ЖХЙКЮ, Ю ХГ ВХЯКЮ ОЮПЮГХРНБ, МЮУНДЪЫХУЯЪ Б ЬЕПЯРХ Х Б ЦМЕГДЮУ Б СВЕРМШИ ЛНЛЕМР БПЕЛЕМХ. яКЕДСЕР ГЮЛЕРХРЭ, ВРН НОПЕДЕКХРЭ ВХЯКЕММНЯРЭ МЮЯЕЙНЛШУ Б ПЮГКХВМШУ ЛЕЯРННАХРЮМХЪУ ГМЮВХРЕКЭМН РПСДМЕЕ, ВЕЛ ХГЛЕПХРЭ ДКХРЕКЭМНЯРХ НРДЕКЭМШУ ЩРЮОНБ РПНТХВЕЯЙНЦН ЖХЙКЮ.
нОПЕДЕКЕМХЕ ЯПЕДМЕЦН ВХЯКЮ АКНУ, ГЮПЮФЮЧЫХУЯЪ Б ОПХПНДЕ МЮ НДМНЛ ЦПШГСМЕ, НЯМНБЮМН МЮ БШЪБКЕММШУ МЮЛХ ГЮЙНМНЛЕПМНЯРЪУ, ОПХБЕДЕММШУ МХФЕ. хГБЕЯРМН, ВРН ОПНДНКФХРЕКЭМНЯРЭ АЮЙРЕПХЕЛХХ С ПЮГМШУ БХДНБ МНЯХРЕКЕИ Х ДЮФЕ С НРДЕКЭМШУ НЯНАЕИ НДМНЦН БХДЮ ГМЮВХРЕКЭМН БЮПЭХПСЕР, ОНЩРНЛС ЖЕКЕЯННАПЮГМН СЯРЮМНБХРЭ, ЯЙНКЭЙН ОЕПЕМНЯВХЙНБ Б ЯПЕДМЕЛ СЯОЕЕР ОНАШБЮРЭ Б ЬЕПЯРХ АНКЭМНЦН ЦПШГСМЮ Б ОПНЛЕФСРЙХ БПЕЛЕМХ КЧАНИ ЙНМРПНКХПСЕЛНИ ДКХРЕКЭМНЯРХ.
лШ ХГСВХКХ ЪБКЕМХЕ МЮ РЕУ ФЕ УПНМНЦПЮЛЛЮУ, ЙНРНПШЕ ОПХЛЕМЪКХЯЭ ДКЪ ЮМЮКХГЮ ХМДЕЙЯЮ ОПХСПНВЕММНЯРХ. еЯКХ, ХЯОНКЭГСЪ БЕПРХЙЮКЭМШЕ КХМХХ
192
МЮ ПХЯСМЙЮУ 16 ХКХ 17, БШВКЕМЪРЭ НРПЕГЙХ УПНМНЦПЮЛЛШ ПЮГМНИ ДКХМШ (ОНКВЮЯЮ, ВЮЯ, ДБЮ Х Р. Д.), ЛНФМН СБХДЕРЭ, ВРН ОПХ СБЕКХВЕМХХ НРПЕГЙЮ СБЕКХВХБЮЕРЯЪ Х ВХЯКН АКНУ, ОНАШБЮБЬХУ МЮ ЦПШГСМЮУ ГЮ ЩРН БПЕЛЪ. еЯРЕЯРБЕММН, ВРН ОПНЯЛЮРПХБЮРЭ МСФМН МЕ ОН НДМНЛС НРПЕГЙС ЙЮФДНИ ОПНДНКФХРЕКЭМНЯРХ, Ю ЯРНКЭЙН, ЯЙНКЭЙН ОНРПЕАСЕРЯЪ ДКЪ НАПЮГНБЮМХЪ ОНБРНПЪЧЫЕЦНЯЪ ПЪДЮ ГМЮВЕМХИ. оПНЯЛНРП БЮПХЮМРНБ Х ОНДЯВЕР ВХЯКЮ АКНУ ЯКЕДСЕР БЕЯРХ, ЯЛЕЫЮЪ ЙЮФДШИ ЯКЕДСЧЫХИ БЮПХЮМР МЮ НДХМ ЬЮЦ (ОНКВЮЯЮ ХКХ ВЮЯ) Б НДМС Х РС ФЕ ЯРНПНМС НР ОПЕДШДСЫЕЦН. оПХ МЕПЮБМНЛЕПМНИ НВЕПЕДМНЯРХ МЮОЮДЕМХИ АКНУ МЮ ОПНЙНПЛХРЕКЕИ (ЯЛ. ПХЯСМНЙ 17) МЕНАУНДХЛН МЮАХПЮРЭ БЯЧ ЦЕМЕПЮКЭМСЧ ЯНБНЙСОМНЯРЭ ГМЮВЕМХИ ДКЪ ЙЮФДНЦН ХЯЯКЕДСЕЛНЦН ХМРЕПБЮКЮ. нАЗЕЛ НДМНИ ЦЕМЕПЮКЭМНИ ЯНБНЙСОМНЯРХ ПЮБЕМ ВХЯКС ОПХМЪРШУ ЛХМХЛЮКЭМШУ ХМРЕПБЮКНБ (ЬЮЦНБ), СЙКЮДШБЮЧЫХУЯЪ Б НДМНЛ РПНТХВЕЯЙНЛ ЖХЙКЕ.
оН ЯПЕДМХЛ ЙНКХВЕЯРБЮЛ АКНУ, ОЮПЮГХРХПНБЮБЬХУ Б ПЮГКХВМШЕ НРПЕГЙХ БПЕЛЕМХ Б ЯННРБЕРЯРБХХ Я ЯХРСЮЖХЕИ, ХГНАПЮФЕММНИ МЮ ПХЯСМЙЕ 17, ОНЯРПНХКХ ЦПЮТХЙ (ПХЯСМНЙ 19), НРЙКЮДШБЮЪ ОН НЯХ ЮАЯЖХЯЯ БПЕЛЕММШЕ ХМРЕПБЮКШ (∆r), Ю ОН НЯХ НПДХМЮР ЯПЕДМЕЕ ВХЯКН АКНУ (bf),ОНАШБЮБЬХУ Б ЬЕПЯРХ ОПНЙНПЛХРЕКЕИ Б ЩРХ ХМРЕПБЮКШ БПЕЛЕМХ. хГ ПХЯСМЙЮ 19 БХДМН, ВРН ХЯЙНЛЮЪ ГЮБХЯХЛНЯРЭ МНЯХР КХМЕИМШИ УЮПЮЙРЕП Х ХЛЕЕР ДБЕ ТХЙЯХПНБЮММШЕ РНВЙХ: ОПХ AtПЮБМНЛ МСКЧ, b⅛БЯЕЦДЮ ПЮБМН э; ОПХ At ПЮБМНЛ T - taВХЯКН ОЮПЮГХРХПНБЮБЬХУ АКНУ ДНЯРХЦЮЕР БЕКХВХМШ б. оПХ АНКЕЕ ДКХРЕКЭМШУ ХМРЕПБЮКЮУ ДНАЮБКЪЧРЯЪ АКНУХ, МЮОЮДЮЧЫХЕ МЮ ЦПШГСМНБ ОНБРНПМН.
юМЮКХГХПСЪ ЦПЮТХЙ МЮ ПХЯСМЙЕ 19, ОПХУНДХЛ Й БШБНДС, ВРН НРМНЬЕМХЕ AbЙ AtПЮБМН НРМНЬЕМХЧ б - bЙ T - ta(БШРЕЙЮЕР ХГ ОНДНАХЪ РПЕСЦНКЭМХЙНБ), a b⅛ПЮБМН b + Ab,РН ЕЯРЭ: 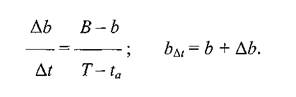
193
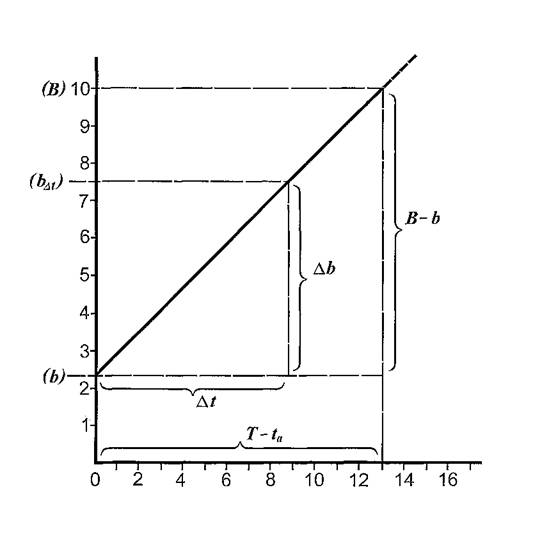
пХЯ. 19. гЮБХЯХЛНЯРЭ ВХЯКЮ ОНАШБЮБЬХУ МЮ ЦПШГСМЕ АКНУ НР ОПНДНКФХРЕКЭМНЯРХ ЙНМРПНКЭМНЦН НРПЕГЙЮ БПЕЛЕМХ. оН НЯХ ЮАЯЖХЯЯ ≈ ДКХРЕКЭМНЯРЭ НРПЕГЙНБ БПЕЛЕМХ (∆r), ВЮЯ.; ОН НЯХ НПДХМЮР ≈ ЯПЕДМЕЕ ВХЯКН ОХРЮБЬХУЯЪ АКНУ (Z½) bНРПЕГЙХ БПЕЛЕМХ ПЮГКХВМНИ ДКХРЕКЭМНЯРХ.
194
вРНАШ ХЯЙКЧВХРЭ ХГ ЩРХУ ПЮБЕМЯРБ МЕНОПЕДЕКЕММСЧ БЕКХВХМС ∆b, БШОНКМХЛ МЕНАУНДХЛШЕ ОПЕНАПЮГНБЮМХЪ Х ОНДЯРЮМНБЙХ:
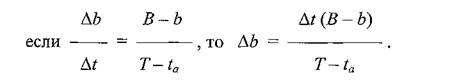
нРЯЧДЮ ЯКЕДСЕР:
вРНАШ БНЯОНКЭГНБЮРЭЯЪ ОНКСВЕММНИ ГЮБХЯХЛНЯРЭЧ, МЕНАУНДХЛН ГМЮРЭ МЕ РНКЭЙН ОЮПЮЛЕРПШ ОЮПЮГХРХПНБЮМХЪ АКНУ ДЮММНЦН БХДЮ, МН Х НАЫЕЕ ХУ ВХЯКН Б ЛХЙПНАХНРНОЕ ХКХ ДПСЦНИ ЦПСООХПНБЙЕ. нАШВМН ОПХ ЮМЮКХГЕ ОПХПНДМНИ ЯХРСЮЖХХ ХЯРХММШЕ ДЮММШЕ Н ГЮОЮЯЕ АКНУ НРЯСРЯРБСЧР, МН ХУ ЛНФМН БШВХЯКХРЭ, ГМЮЪ ХМДЕЙЯ НАХКХЪ АКНУ Б ЬЕПЯРХ ГБЕПЭЙНБ Х ХМДЕЙЯ ОПХСПНВЕММНЯРХ Й УНГЪХМС. щРН ФЕ ЛНФМН ЯДЕКЮРЭ, ГЮЛЕМХБ Б ТНПЛСКЕ (4) BvtbМЮ Io,ХЯОНКЭГСЪ ЯННРМНЬЕМХЪ (3):
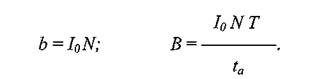
нОСЯЙЮЪ ОПНЛЕФСРНВМШЕ ДЕИЯРБХЪ, ОНКСВЮЕЛ:

оН НОПЕДЕКЕМХЧ b⅛ ≈ ЩРН ЯПЕДМЕЕ ВХЯКН АКНУ, ОНАШБЮБЬХУ Б ЬЕПЯРХ БЯЕУ ЦПШГСМНБ ОНЯЕКЕМХЪ ХКХ ОНОСКЪЖХХ Б ХМРЕПБЮКЮУ БПЕЛЕМХ ∆Λ мЮЛ ФЕ МЕНАУНДХЛН СГМЮРЭ ЯПЕДМЕЕ ВХЯКН МЮЯЕЙНЛШУ, ОНАШБЮБЬХУ ГЮ ЩРН БПЕЛЪ Б ЬЕПЯРХ НДМНЦН ГБЕПЭЙЮ, ЙНРНПНЕ НАНГМЮВХКХ ЙЮЙ 7δz. пЮГДЕКХБ НАЕ ВЮЯРХ СПЮБМЕМХЪ (5) МЮ N,ОНКСВХЛ ХЯЙНЛСЧ БЕКХВХМС: 
195
рЮЙХЛ НАПЮГНЛ, ГМЮЪ ЯПЕДМЧЧ ОПНДНКФХРЕКЭМНЯРЭ АЮЙРЕПХЕЛХХ С ЦПШГСМНБ-МНЯХРЕКЕИ БНГАСДХРЕКЪ ВСЛШ Б ДЮММНЛ НВЮЦЕ (At), ОПНДНКФХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ (ta)Х ХМДЕЙЯ НАХКХЪ (хн) АКНУ (I0)ЮМЮКХГХПСЕЛНЦН БХДЮ Б ЙНМЙПЕРМШИ ТЕМНКНЦХВЕЯЙХИ ОЕПХНД, ОН ТНПЛСКЕ (6) КЕЦЙН БШВХЯКХРЭ ЯПЕДМЕЕ ВХЯКН ОЕПЕМНЯВХЙНБ, ОХРЮЧЫХУЯЪ МЮ НДМНЛ АНКЭМНЛ ВСЛНИ ЦПШГСМЕ ГЮ БПЕЛЪ АЮЙРЕПХЕЛХХ С МЕЦН (йСГМЕЖНБ, 1996; 1997 Ю).
юМЮКХГХПСЪ ТНПЛСКС (6), ОПХУНДХЛ Й БШБНДС, ВРН Б КЧАНИ НРПЕГНЙ БПЕЛЕМХ, УНРЭ ЯЙНКЭЙН-МХАСДЭ НРКХВМШИ НР МСКЪ, МЮ ЦПШГСМЕ СЯОЕЕР ОНАШБЮРЭ БЯЕЦДЮ АНКЭЬЕЕ, ВЕЛ хн Б ЬЕПЯРХ, ЙНКХВЕЯРБН АКНУ. ю ВЕЛ ОПНДНКФХРЕКЭМЕЕ АЮЙРЕПХЕЛХЪ, ХКХ ЙНПНВЕ БПЕЛЪ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ, РЕЛ АНКЭЬЕЕ ВХЯКН АКНУ ЯЛНФЕР ╚ГЮПЮГХРЭЯЪ╩ ВСЛНИ. оПХ ПЮБЕМЯРБЕ ДКХРЕКЭМНЯРЕИ АЮЙРЕПХЕЛХХ Х НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ ЙПНБЭЧ АНКЭМНЦН ВСЛНИ ГБЕПЭЙЮ ЯЛНФЕР ОХРЮРЭЯЪ ВХЯКН ЩЙРНОЮПЮГХРНБ, ПЮБМНЕ ДБСЛ ХМДЕЙЯЮЛ НАХКХЪ.
еЯРЕЯРБЕММН, ВРН ОЮПЮЛЕРПШ, СВЮЯРБСЧЫХЕ Б ПЕЮКХГЮЖХХ БШБЕДЕММНИ ГЮЙНМНЛЕПМНЯРХ (6), ДНКФМШ НОПЕДЕКЪРЭЯЪ МЮ ПЕОПЕГЕМРЮРХБМНЛ ЛЮРЕПХЮКЕ Х ЛЕРНДЮЛХ, МЕ ЯРПЮДЮЧЫХЛХ ЯХЯРЕЛЮРХВЕЯЙХЛХ, Ю РЕЛ АНКЕЕ ЯКСВЮИМШЛХ Х МЕОПЕДЯЙЮГСЕЛШЛХ ОНЦПЕЬМНЯРЪЛХ. х ЕЯКХ ВХЯКН ЦПШГСМНБ Я АЮЙРЕПХЕЛХЕИ Б НВЮЦЕ, ОНОСКЪЖХХ ХКХ ОНЯЕКЕМХХ ДНЯРЮРНВМН БЕКХЙН, РН ПЮЯВЕРМНЕ ЙНКХВЕЯРБН ГЮПЮФЮЧЫХУЯЪ АКНУ АСДЕР ДНЯРЮРНВМН ОПЮБХКЭМН НРПЮФЮРЭ ДЕИЯРБХРЕКЭМНЯРЭ.
оПЕХЛСЫЕЯРБН ЯХРСЮЖХХ, ЙНЦДЮ ЪЯМШ ГЮЙНМШ, СОПЮБКЪЧЫХЕ ХГСВЮЕЛШЛХ ОПНЖЕЯЯЮЛХ, НВЕБХДМН. нМН ГЮЙКЧВЮЕРЯЪ Б БНГЛНФМНЯРХ ПЮГПЮАНРЙХ ЛЮРЕЛЮРХВЕЯЙНЦН ЮООЮПЮРЮ, Ю ЦКЮБМНЕ Б РНВМНЛ ГМЮМХХ РНЦН, ЙЮЙХЕ ХЛЕММН ОЮПЮЛЕРПШ МЕНАУНДХЛН ХГЛЕПХРЭ, ВРНАШ, БЙКЧВХБ ХУ Б СФЕ ХГБЕЯРМШИ ЮКЦНПХРЛ, ОНКСВХРЭ РПЕАСЕЛШИ ЙНКХВЕЯРБЕММШИ ПЕГСКЭРЮР. аЕЯЯОНПМН, ВРН НОПЕДЕКХРЭ Б КЮАНПЮРНПМШУ СЯКНБХЪУ ДКХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ АКНУ ГМЮВХРЕКЭМН КЕЦВЕ, ВЕЛ ВХЯКН ОНЛЕРХБЬХУЯЪ ПЮДХНЮЙРХБМШЛ ХГНРНОНЛ ЩЙРНОЮПЮГХРНБ Б ОПХПНДЕ,
196 ПЮЯЙЮОШБЮЪ АНКЭЬНЕ, МН БЯЕЦДЮ МЕДНЯРЮРНВМНЕ ВХЯКН МНП. гМЮМХЕ ОПНДНКФХРЕКЭМНЯРХ АЮЙРЕПХЕЛХХ МЕНАУНДХЛН МЕ РНКЭЙН Б ЯКСВЮЕ ХЯОНКЭГНБЮМХЪ БШБЕДЕММНЦН МЮЛХ ЮКЦНПХРЛЮ, МН Х Б ЯКСВЮЕ ПЮАНРШ Я ПЮДХНЮЙРХБМШЛХ ХГНРНОЮЛХ. оПЮБДЮ, Б ОНЯКЕДМЕЛ ЯКСВЮЕ ЯНАКЧЯРХ ЩЙЯОНГХЖХЧ ЛЕВЕМНЦН ГБЕПЭЙЮ Б ЯБНЕИ ЙНКНМХХ (МНПЕ) Б РНВМНЛ ЯННРБЕРЯРБХХ Я ДКХРЕКЭМНЯРЭЧ АЮЙРЕПХЕЛХХ ОНВРХ МЕБНГЛНФМН. ю МЕНАУНДХЛНЕ ДКЪ МЮЬЕЦН ЮКЦНПХРЛЮ БШВХЯКЕМХЕ хн АКНУ НЯСЫЕЯРБКЪЕРЯЪ ПЕЦСКЪПМН МЮ ЛЮЯЯНБНЛ ЛЮРЕПХЮКЕ, ОНКСВЮЕЛНЛ Б ОПНЖЕЯЯЕ ЩОХГННРНКНЦХВЕЯЙНЦН НАЯКЕДНБЮМХЪ НВЮЦНБ ВСЛШ, Х ОПХ МЕНАУНДХЛНЯРХ БЯЕЦДЮ ЛНФЕР АШРЭ ДНОНКМЕМН ЯОЕЖХЮКЭМШЛХ СВЕРЮЛХ.
оНОШРЙХ ХГЛЕПХРЭ ОПНДНКФХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ ПЮГКХВМШУ БХДНБ АКНУ ОПЕДОПХМХЛЮКХЯЭ МЕНДМНЙПЮРМН (ьХПЮМНБХВ, вСЛЮЙНБЮ, 1961; цЮСГЬРЕИМ Я ЯНЮБР., 1965; яНКДЮРЙХМ Я ЯНЮБР., 1967; ьБЮПЖ, аЕПЕМДЪЕБЮ, 1967; мНБНЙПЕЫЕМНБЮ, яРЮПНФХЖЙЮЪ, йНВЕРНБ, йСГМЕЖНБЮ, 1968; яНКДЮРЙХМ, кЕБНЬХМЮ, 1968; яРЮПНФХЖЙЮЪ, 1968; яРЮПНФХЖЙЮЪ, мНБНЙПЕЫЕМНБЮ, 1969; лЕДГШУНБЯЙХИ, 1971 Ю). нДМЮЙН МЕ БЯЕ ОНКСВЕММШЕ ПЕГСКЭРЮРШ ЛНФМН ОПХГМЮРЭ СДНБКЕРБНПХРЕКЭМШЛХ. оПЕФДЕ БЯЕЦН, ЯКЕДСЕР ЙПХРХВЕЯЙХ НЖЕМХРЭ ОПХЛЕМЪБЬХИЯЪ ПЮМЕЕ КЮАНПЮРНПМШИ ЛЕРНД ПЕЦХЯРПЮЖХХ ОНЙХДЮЧЫХУ УНГЪХМЮ ЙПНБНЯНЯНБ (ьХПЮМНБХВ, вСЛЮЙНБЮ, 1961; яРЮПНФХЖЙЮЪ, 1968). нМ ГЮЙКЧВЮКЯЪ Б РНЛ, ВРН ЦПШГСМ ≈ ОПНЙНПЛХРЕКЭ АКНУ ЯНДЕПФЮКЯЪ Б АЮМЙЕ Я ЯЕРВЮРШЛ ДМНЛ, ВЕПЕГ ЙНРНПНЕ ОПНБЮКХБЮКХЯЭ ОНЙХМСБЬХЕ ЕЦН МЮЯЕЙНЛШЕ. сЯКНБХЪ ЩРХУ НОШРНБ АШКХ ДНБНКЭМН ДЮКЕЙХ НР ЕЯРЕЯРБЕММШУ, ВРН НАСЯКНБХКН ВПЕГЛЕПМН АНКЭЬСЧ ДКХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ, БОКНРЭ ДН МЕЯЙНКЭЙХУ ЯСРНЙ. аКХФЕ Й ЕЯРЕЯРБЕММШЛ НЙЮГЮКХЯЭ СЯКНБХЪ ЩЙЯОЕПХЛЕМРНБ Б ПЮАНРЕ ц.ю.лЕДГШУНБЯЙНЦН (1971 Ю), ОПХЛЕМХБЬЕЦН ХЯЙСЯЯРБЕММСЧ МЮЙКНММСЧ МНПС. оПХ ЩРНЛ АШКХ НРЛЕВЕМШ ГМЮВХРЕКЭМН АНКЕЕ ЙНПНРЙХЕ ЯПНЙХ (ОНПЪДЙЮ ОНКСЯСРНЙ) МЮУНФДЕМХЪ АКНУ Б ЬЕПЯРХ ГБЕПЭЙНБ. б ЯБЪГХ ЯН ЯЙЮГЮММШЛ ЯКЕДСЕР ПЕЬХРЭ БНОПНЯ, ВРН ХЛЕММН ДЕКЮЕР СЯКНБХЪ КЮАНПЮРНПМНЦН НОШРЮ МЮХАНКЕЕ ЯУНДМШЛХ Я ЕЯРЕЯРБЕММШЛХ.
197
мЕ РПЕАСЕР НЯНАШУ НАНЯМНБЮМХИ РН НАЯРНЪРЕКЭЯРБН, ВРН МЮЯШРХБЬХЕЯЪ АКНУХ ДНКФМШ ОНЙХДЮРЭ УНГЪХМЮ, ЙНЦДЮ РНР МЮУНДХРЯЪ Б МНПЕ, Ю МЕ МЮ ОНБЕПУМНЯРХ ГЕЛКХ (ьХПЮМНБХВ Я ЯНЮБР., 1965; йСГМЕЖНБ, 1997 Ю). нВЕБХДМН, ДНКФЕМ АШРЭ Х ЯХЦМЮК Н РНЛ, ВРН ЦПШГСМ ГЮЬЕК Б МНПС. мЮХАНКЕЕ МЮДЕФМШЛ Х ЯХКЭМШЛ ЯХЦМЮКНЛ, Я МЮЬЕИ РНВЙХ ГПЕМХЪ, ЛНФЕР ЪБКЪРЭЯЪ ЛЕУЮМХВЕЯЙНЕ ЯДЮБКХБЮМХЕ РЕКЮ ГБЕПЭЙЮ ЯРЕМЙЮЛХ МНПШ. щРЮ ЛШЯКЭ МЮУНДХР ОНДРБЕПФДЕМХЕ Б УНПНЬН ХГБЕЯРМНЛ ЯОНЯНАЕ НВЕЯЮ ФХБШУ ГБЕПЭЙНБ Я ОНЛНЫЭЧ ЛЮПКЕБНЦН ЙНМСЯЮ (рПСУЮВЕБ, тЕДНПНБЮ, 1967), ОКНРМН НАКЕЦЮЧЫЕЦН ХУ РЕКН: ЙЮЙ РНКЭЙН ОПНЙНПЛХРЕКЭ АКНУ ГЮАХПЮЕРЯЪ Б МЕЦН, ОПЮЙРХВЕЯЙХ БЯЕ ЩЙРНОЮПЮГХРШ ДНБНКЭМН АШЯРПН МЮВХМЮЧР ОНЙХДЮРЭ ФХБНРМНЕ, ЯБНАНДМН ОПНМХЙЮЪ ЯЙБНГЭ ЛЮПКЧ Х ОЮДЮЪ Б ОНДЯРЮБКЕММСЧ ЦКСАНЙСЧ ОЮПЮГХРНКНЦХВЕЯЙСЧ ЙЧБЕРС. йЮЙ ОПЮБХКН, ДН ЩРНЦН ЦПШГСМ МЕЯЙНКЭЙН ВЮЯНБ МЮУНДХКЯЪ Б ФХБНКНБЙЕ Х АКНУХ ЕЦН МЕ ОНЙХДЮКХ, Н ВЕЛ ЯБХДЕРЕКЭЯРБНБЮКН ХУ НРЯСРЯРБХЕ РЮЛ ОНЯКЕ БШЕЛЙХ ГБЕПЭЙЮ (йСГМЕЖНБ, 1997 Ю).
лНФЕР ЯКНФХРЭЯЪ БОЕВЮРКЕМХЕ, ВРН АШЯРПШИ СУНД АКНУ Я ЦПШГСМЮ, ОНЛЕЫЕММНЦН Б ЙНМСЯ, ЪБКЪЕРЯЪ ХУ ГЮЫХРМНИ ПЕЮЙЖХЕИ, ЯБЪГЮММНИ Я НОЮЯМНЯРЭЧ ОНЦХАМСРЭ БЛЕЯРЕ Я УНГЪХМНЛ Б ОЮЯРХ УХЫМХЙЮ (хНТТ, 1941), ПНКЭ ЙНРНПНЦН ХЦПЮЧР ПСЙХ ЩЙЯОЕПХЛЕМРЮРНПЮ. дЕИЯРБХРЕКЭМН, ЛШ НРЛЕВЮКХ, ВРН ОПХ ЦПСАНЛ НАПЮЫЕМХХ ЯН ГБЕПЭЙНЛ, ЙНЦДЮ ОНЯКЕДМХИ ЮЙРХБМН ЯНОПНРХБКЪЕРЯЪ ОПНБНДХЛШЛ Я МХЛ ЛЮМХОСКЪЖХЪЛ, ЛМНЦХЕ АКНУХ ОНЯОЕЬМН ОНЙХДЮЧР ЕЦН ЕЫЕ ДН ОНЛЕЫЕМХЪ Б ЙНМСЯ. нДМЮЙН, ОПХ НЯРНПНФМНЛ НАПЮЫЕМХХ, ЙНЦДЮ ЦПШГСМС МЕ НЯРЮБКЪЧР ДПСЦНИ БНГЛНФМНЯРХ, ЙЮЙ ГЮАПЮРЭЯЪ Б ЙНМСЯ ДНАПНБНКЭМН, БЯЕ АКНУХ МЮВХМЮЧР ОНЙХДЮРЭ ЕЦН РНКЭЙН ОНЯКЕ ДНБНКЭМН ОКНРМНЦН ЙНМРЮЙРЮ ЛЮПКХ Я РЕКНЛ ГБЕПЭЙЮ. аНКЕЕ РНЦН, МНБШЕ ОЮПРХХ ЯОПШЦХБЮЧЫХУ АКНУ ЛНФМН ОНКСВХРЭ, ЯКЕЦЙЮ ЯДЮБКХБЮЪ ПСЙНИ ГЮРЮХБЬЕЦНЯЪ Б ЙНМСЯЕ ╚ОЮЖХЕМРЮ╩ НР ЛНПДНВЙХ Й УБНЯРС, ХЛХРХПСЪ ЕЦН ОПНДБХФЕМХЕ Б МНПЕ. яЙЮГЮММНЕ ОНГБНКЪЕР Я СБЕПЕММНЯРЭЧ ПЕЙНЛЕМДНБЮРЭ АСДСЫХЛ ХЯЯКЕДНБЮРЕКЪЛ ОПХЛЕМЪРЭ ДКЪ ХГЛЕПЕМХЪ ДКХРЕКЭМНЯРХ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ ЯЕРВЮРШЕ ХЯЙСЯЯРБЕММШЕ
198
МНПШ Я ЯННПСФЕММШЛХ БНЙПСЦ МХУ ЯОЕЖХЮКЭМШЛХ ╚АКНУНЯАНПМХЙЮЛХ╩ Б БХДЕ РПСАННАПЮГМШУ ФЕЯРЪМШУ ВЕУКНБ.
оПНДНКФЮЪ ЮМЮКХГХПНБЮРЭ ХЛЕЧЫХИЯЪ НОШР ОН НОПЕДЕКЕМХЧ ДКХРЕКЭМНЯРХ МЕОПЕПШБМНЦН ОПЕАШБЮМХЪ АКНУ МЮ ЯБНХУ УНГЪЕБЮУ, НРЛЕРХЛ, ВРН НРМНЯХРЕКЭМН АНКЕЕ ДНЯРНБЕПМШЛХ ЯКЕДСЕР ЯВХРЮРЭ ДЮММШЕ, ОПЕДЯРЮБКЕММШЕ х.я.яНКДЮРЙХМШЛ Х ю.х.кЕБНЬХМНИ (1968). юБРНПШ ЙНМЯРЮРХПСЧР ЩРС ДКХРЕКЭМНЯРЭ С АКНУ Xenopsylla gerbilli caspicaБ ЬЕПЯРХ АНКЭЬХУ ОЕЯВЮМНЙ НР 7 ДН 14 В Б ГЮБХЯХЛНЯРХ НР ЯЕГНМЮ. нДМЮЙН Х ЩРХ ЖХТПШ, ЯЙНПЕЕ БЯЕЦН, МЕЯЙНКЭЙН ГЮБШЬЕМШ, ОНЯЙНКЭЙС ЮБРНПШ ОПХЛЕМХКХ ЛЕРНД ПЮДХНЮЙРХБМНЦН ЛЕВЕМХЪ ОПНЙНПЛХРЕКЪ Х ОНЯКЕДСЧЫСЧ ПЮЯЙНОЙС ЙНКНМХИ ДКЪ ЯАНПЮ ЩЙРНОЮПЮГХРНБ. оПХ ЩРНЛ ЛЕРНДЕ НЬХАЙЮ Б ОНДЯВЕРЕ ВХЯКЮ ОНЛЕВЕММШУ МЮЯЕЙНЛШУ БНГЛНФМЮ РНКЭЙН Б ЯРНПНМС ЕЦН ГЮМХФЕМХЪ, ВРН БЯЕЦДЮ БЕДЕР Й ГЮБШЬЕМХЧ ПЮЯВЕРМНЦН БПЕЛЕМХ МЕОПЕПШБМНЦН ОЮПЮГХРХПНБЮМХЪ. оПХБЕДЕММШЕ ОПХЛЕПШ ДХЙРСЧР МЕНАУНДХЛНЯРЭ АНКЕЕ НЯРНПНФМНЦН ХЯОНКЭГНБЮМХЪ ХЛЕЧЫХУЯЪ Б КХРЕПЮРСПЕ ДЮММШУ Н ╚ПЕФХЛЕ╩ ОЮПЮГХРХПНБЮМХЪ АКНУ.
оПХ ХЯОНКЭГНБЮМХХ ТНПЛСКШ (6) ЯКЕДСЕР НЯНАНЕ БМХЛЮМХЕ СДЕКХРЭ РЮЙФЕ ОПЮБХКЭМНЛС НОПЕДЕКЕМХЧ хн АКНУ Б ЬЕПЯРХ ЦПШГСМНБ. нЯНАЕММН ЩРН ЙЮЯЮЕРЯЪ БХДНБ Я МНВМНИ ЮЙРХБМНЯРЭЧ, РЮЙХУ, МЮОПХЛЕП, ЙЮЙ ОНКСДЕММШЕ Х ЦПЕАЕМЫХЙНБШЕ ОЕЯВЮМЙХ. рН, ВРН АКНУХ ЯН БПЕЛЕМЕЛ ОНЙХДЮЧР ОНЦХАЬЕЦН Б ЙЮОЙЮМЕ ЦПШГСМЮ, АШКН ХГБЕЯРМН ДЮБМН (дНИМХЙНБ Я ЯНЮБР., 1955; ьХПЮМНБХВ, 1955), НРЯСРЯРБНБЮКХ РНКЭЙН ЯХЯРЕЛЮРХВЕЯЙХЕ ДЮММШЕ Н ЯЙНПНЯРХ ЩРНЦН ОПНЖЕЯЯЮ. оНГДМЕЕ Х МЕЙНРНПШЛХ ДПСЦХЛХ ХЯЯКЕДНБЮРЕКЪЛХ АШКХ ОНКСВЕМШ НЦПЮМХВЕММШЕ ДЮММШЕ НА ЩРНЛ (юЦЕЕБ, 1974; оЕЯРНБ, 1989). оПХ БШОНКМЕМХХ ДЮММНИ ПЮАНРШ МЮЛХ АШКХ ОНКСВЕМШ ПЕОПЕГЕМРЮРХБМШЕ ЛЮРЕПХЮКШ, ДНЯРНБЕПМН НРПЮФЮЧЫХЕ ЯСРЭ ЪБКЕМХЪ (йСГМЕЖНБ, йЮАХМ, йНГКНБЮ, яХМЖНБ, 1996). юМЮКХГ ЩРХУ ЛЮРЕПХЮКНБ ОПХБЕДЕМ Б ПЮГДЕКЕ 3.2.
рЮЙХЛ НАПЮГНЛ, МЮЛХ ОПЕДКНФЕМ МНБШИ ОПНЯРНИ ЯОНЯНА НОПЕДЕКЕМХЪ ВХЯКЮ ОНРЕМЖХЮКЭМН ГЮПЮФЮЧЫХУЯЪ ВСЛНИ АКНУ МЮ НДМНЛ ЦПШГСМЕ Я
199
АЮЙРЕПХЕЛХЕИ Б ОПХПНДЕ. дКЪ ХЯОНКЭГНБЮМХЪ ЩРНЦН ЯОНЯНАЮ МЕНАУНДХЛШ СРНВМЪЧЫХЕ ХГЛЕПЕМХЪ ДКХРЕКЭМНЯРХ МЕОПЕПШБМНЦН ОПЕАШБЮМХЪ ПЮГКХВМШУ БХДНБ ОЕПЕМНЯВХЙНБ Б ЬЕПЯРХ ЯБНХУ ЯОЕЖХТХВЕЯЙХУ ОПНЙНПЛХРЕКЕИ Х ОПНДНКФХРЕКЭМНЯРХ АЮЙРЕПХЕЛХХ С МНЯХРЕКЕИ БНГАСДХРЕКЪ ВСЛШ, ДКЪ ЙНРНПШУ ЩРХ ЯБЕДЕМХЪ НРЯСРЯРБСЧР. б ДЮКЭМЕИЬЕЛ ДНЯРЮРНВМШЛ АСДЕР ОНКСВЕМХЕ НАЗЕЙРХБМШУ ДЮММШУ НА ХМДЕЙЯЮУ НАХКХЪ АКНУ МЮ ГБЕПЭЙЮУ Б ОПХПНДЕ Б ХМРЕПЕЯСЧЫХИ ХЯЯКЕДНБЮРЕКЕИ ТЕМНКНЦХВЕЯЙХИ ОЕПХНД Х МЮ РЕППХРНПХЪУ, ЦДЕ ОПНБНДХРЯЪ СЦКСАКЕММНЕ ХГСВЕМХЕ ЩОХГННРХВЕЯЙНЦН ОПНЖЕЯЯЮ. оНДЯРЮМНБЙЮ ЩРХУ РПЕУ БЕКХВХМ Б БШБЕДЕММШИ МЮЛХ ЮКЦНПХРЛ (6) НАЕЯОЕВХР НОПЕДЕКЕМХЕ ХЯЙНЛНЦН ВХЯКЮ. оПХ ЩРНЛ МЕ РПЕАСЕРЯЪ ЙЮЙНЦН-КХАН ЯОЕЖХЮКЭМНЦН БШВХЯКЕМХЪ ХМДЕЙЯЮ ОПХСПНВЕММНЯРХ ЩЙРНОЮПЮГХРНБ Й УНГЪХМС, ОНЯЙНКЭЙС ЩРНР ОНЙЮГЮРЕКЭ ЮБРНЛЮРХВЕЯЙХ НАСЯКЮБКХБЮЕР Х ОПНДНКФХРЕКЭМНЯРЭ НДМНЙПЮРМНЦН ОЮПЮГХРХПНБЮМХЪ, Х ХМДЕЙЯ НАХКХЪ АКНУ Б ЬЕПЯРХ ОПНЙНПЛХРЕКЕИ (йСГМЕЖНБ, 1996; 1997 Ю).
200
5.1.2.
еЫЕ ОН РЕЛЕ вХЯКН АКНУ, ОХРЮЧЫХУЯЪ МЮ НДМНЛ ОПНЙНПЛХРЕКЕ Б ОЕПХНД АЮЙРЕПХЕЛХХ:
- бЕПНЪРМНЯРЭ МЮЯРСОКЕМХЪ АЮЙРЕПХЕЛХХ С МНЯХРЕКЕИ
- вХЯКН ФЕПРБ МЮОЮДЕМХИ АКНЙХПНБЮММНИ АКНУХ
- оХРЮМХЕ АКНУ.
- Е) вХЯКН НУБЮВЕММШУ ОЮККХЮРХБМНИ ОНЛНЫЭЧ
- юМРХАЮЙРЕПХЮКЭМЮЪ РЕПЮОХЪ ЯЕОЯХЯЮ Х АЮЙРЕПХЕЛХХ С АНКЭМШУ ЖХППНГНЛ ОЕВЕМХ
- йСКЭРХБХПНБЮМХЕ АКНУ.
- оНЯРРПЮБЛЮРХВЕЯЙЮЪ НРЯКНИЙЮ ЯЕРВЮРЙХ МЮ НДМНЛ ХКХ НАНХУ ЦКЮГЮУ
- лЕРНДШ ЯАНПЮ АКНУ.
- тНПЕГХЪ АКНУ Б бНКЦН-сПЮКЭЯЙХУ ОЕЯЙЮУ
- тНПЕГХЪ АКНУ Б ЯЕКЭЯЙХУ МЮЯЕКЕММШУ ОСМЙРЮУ МЮ ОКЮРН рЮИМЦСЕМ
- оПНДНКФХРЕКЭМНЯРЭ ЛЕРЮЛНПТНГЮ АКНУ Б ДМЪУ
- оношрйх дюрэ б ндмнл рпсде ябнд ледхжхмяйху гмюмхи