СЕЗОННОСТЬ И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНЕННЫХ циклов
Жизненные циклы клещей, обитающих в разнообразных климатических зонах и заселяющих разные биотопы, значительно отличаются по общей продолжительности и сезонности нападения на хозяев, размножения и линек.
Как и у других групп наземных беспозвоночных, они обеспечивают адаптацию вида к циклическим изменениям климата путем синхронизации с последними сезонных проявлений своей жизнедеятельности. Вопросы сезонности жизненных циклов клещей в связи с их историческим развитием рассмотрены Г. В. Сердюковой (I960), определяющей продолжительность цикла развития как адаптацию к ландшафту с определенным чередованием сезонов года и набором экологически близких групп хозяев. В цитируемой работе выделяются непрерывный, одно-, двух- и трехгодичные циклы развития иксодовых клещей, но благодаря исследованиям Е. М. Хейсина с сотрудниками (1955а, 19596) доказана возможность и более продолжительных циклов в северной части ареалов клещей Ixodes ricinus и /. persulcatus. Наиболее простым и примитивным Г. В. Сердюкова считает непрерывный цикл, когда отсутствует синхронизация отдельных стадий развития с определенными сезонами года. Предполагалось, что подобный тип развития был свойствен предкам современных видов иксодид, обитавшим в районах с влажным тропическим климатом, а в настоящее время встречается у норовых видов. К сожалению, недостаточная изученность тропических видов клещей не дает возможности судить о характере жизненных циклов в тропиках, тогда как у всех изученных в этом отношении гнездово-норовых видов умеренных широт обнаружены различные сезонноциклические адаптации, так что нет оснований говорить о существовании видов с непрерывным циклом развития.При одно- и многолетних циклах синхронность развития с сезонностью климата у многих изученных в этом отношении видов, как и у насекомых, обеспечивается возникновением диапаузы. Она впервые была обнаружена
H.
И. Алфеевым (1948, 1954) у клещей Dermacentor marginatus, D. ріс- tus, Ixodes ricinus и I. persulcatus. В дальнейшем особенности диапаузы и механизм ее возникновения изучался многими исследователями (Нец- кий и Ольшевская, 1950; Разумова, 1960, 1965, 19666; Белозеров, 1963, 1965а, 19656, 19666; Белозеров и Михайлова, 1966). Диапауза проявляется в форме задержки метаморфоза напитавшихся личинок и нимфI. ricinus и I. persulcatus или остановке оогенеза у напитавшихся в летнеосенний период самок Dermacentor. У голодных имаго рода Dermacentor и в меньшей степени у нимф Ixodes ricinus наблюдается так называемая поведенческая диапауза, проявляющаяся в угнетении агрессивности. Роль внешних факторов в возникновении диапаузы рассматривается в связи с особенностями жизненных циклов клещей родов Ixodes и Derma- centor (см. стр. 60—65).
У гнездово-норовых видов аргасовых клещей, живущих в условиях относительно стабильного микроклимата, сезонность основных процессов жизнедеятельности (сроки питания, размножения и линек) выражена слабее, чем у обитателей открытых биотопов. В тропических районах нападение на хозяев и размножение, судя по наблюдениям над Ornithodo- ros moubata, происходит круглогодично, но при чередовании периодов засухи и дождей также наблюдается определенная сезонность жизненного ритма (Walton, 1957, 1959). В субтропических и умеренных широтах в зимние месяцы активность клещей обычно прекращается вследствие воздействия низких температур. Размножение происходит в летнеосенний период, а в зимнее время у некоторых видов отмечено торможение развития яичников (Остроумова, 1936). За теплый период самки проходят один (О. papillipes, О. verrucosus), а при благоприятных условиях до двух (О. tartakovskyi) и более гонотрофических циклов (Argas persicus). Линька неполовозрелых стадий также происходит в теплое время года. Нападение различных возрастных групп голодных клещей на хозяев наблюдается в различные сроки. По наблюдениям М. В. Поспело- вой-Штром (19596), в одном из горных кишлаков Таджикистана весной на хозяев нападали главным образом личинки, нимфы Ornithodoros papillipes младших возрастов, взрослые и, реже, нимфы старших возрастов.
Летом при общем снижении числа нападений питались преимущественно нимфы средних возрастов, а осенью интенсивность нападения увеличивалась за счет личинок и нимф младших возрастов нового поколения. В этих условиях значительная часть популяции завершила цикл развития за 1 год.Отличительной особенностью жизненных циклов аргазид служит неравномерность развития особей одного поколения. Последнее обусловлено непостоянством числа нимфальных возрастов и исключительной способностью к многолетнему голоданию. По данным лабораторного разведения клещей и наблюдениям в природных биотопах, развитие большинства видов фауны СССР в оптимальных условиях продолжается от полугода до 2 лет, но у части особей таких видов, как Ornithodoros papillipes при недостатке прокормителей может затягиваться до 20 лет (Павловский и Скрынник, 1956, 1960).
Весьма четко сезонность проявляется в жизненном цикле Alveonasus lahorensis. Вылупившиеся летом личинки не способны к питанию и нападают на овец или, реже, других сельскохозяйственных животных только в осенне-зимний период. Питание личинок и нимф трех возрастов занимает 3—6 недель, а отпавшие насосавшиеся нимфы III линяют на взрослых особей только весной. Первый гонотрофический цикл самки проходят за счет пищевых резервов от нимфального питания. Последующие гонотрофические циклы связаны с имагинальныч питанием и яйцекладкой в весенне-летний сезон (Поспелова-Штром, 1953; Кусов, 1957).
Среди иксодовых клещей фауны СССР наиболее близок к непрерывному циклу характер развития однохозяинного Boophilus calcaratus. В тропических странах у видов этого рода наблюдается круглогодичное паразитирование на скоте, тогда как в южных районах СССР (республики Средней Азии, Закавказье, Северный Кавказ, Южная Украина) в зависимости от климатических условий за год успевают развиться 2—3 генерации, а в зимний период нападение клещей прекращается (Марков и Богородицкий, 1935; «Потоцкий, 1945; Галузо, 1949). Число поколений в год лимитируется условиями, необходимыми для развития яйцекладущих самок, яиц и личинок.
В зимний период развитие яиц и нападение личинок тормозится низкими температурами, что связано с переходом их в холодовое оцепенение, тогда как диапауза не обнаружена. Отсутствием диапаузы в жизненном цикле можно объяснить обнаруженную И. Г. Галузо (1949) значительную асинхронность в развитии особей одной популяции В. саі- caratus, так что потомство одной самки может давать за год различное число поколений. В то же время в умеренных широтах у В. calcaratus уже существует определенная приспособленность жизненного цикла к климатическому ритму, проявляющаяся в повышенной холодостойкости у зимующих личинок и яиц.Продолжительность паразитической жизни на теле хозяина в значительно меньшей степени зависит от климатических факторов. По данным многих авторов, от нападения голодных личинок до отпадения напитавшихся самок В. calcaratus проходило в среднем 20—30 дней. Сходные данные были получены для близких видов: В. microplus в Австралии — 22 дня (Hitchcock, 1955b) и 5. caudatus в Японии — 22—24 дня (Kitaoka a. Yajima, 1958а), причем понижение температуры воздуха лишь в незначительной степени удлиняло эти сроки.
Детальные исследования жизненных циклов гнездово-норовых иксо- дид, выполненные за последние годы, не подтвердили существования у них примитивных «непрерывных» циклов развития. Как и у пастбищных видов, жизненные циклы этой биологической группы характеризуются четко выраженными сезонно-циклическими адаптациями, общая продолжительность развития одной генерации занимает не менее года.
По наблюдениям Л. В. Глащинской-Бабенко (1956), жизненный цикл Ixodes plumbeus — паразита ласточек-береговушек согласован с сезонными явлениями в жизни хозяина. В покинутых гнездах зимуют голодные личинки. Весной они нападают на прилетевших ласточек и отпадают с них внутри гнезда. Высокая температура в жилом гнезде (30—32°) обеспечивает быстрое протекание метаморфоза, так что через 15—18 суток после отпадения напитавшихся личинок появляются активные голодные нимфы. Последние через 12—15 дней после питания на птенцах линяют на имаго.
Самки тоже успевают напитаться на заканчивающих гнездовой период птенцах и через 3—8 дней после отпадения приступают к яйцекладке. Отложенные яйца в том же сезоне дают уходящих на зимовку неактивных личинок, хорошо переносящих переохлаждение и длительное, до 9 месяцев, голодание. В результате все развитие — от нападения голодных личинок на хозяина до появления неактивных личинок следующего поколения — занимает 4—4.5 месяца — время гнездования перелетных птиц, а за год развивается одно поколение со строгой сезонной последовательностью стадий развития.Довольно сходен с описанным и жизненный цикл Bhipicephalus schul- zei — обитателей нор грызунов (преимущественно сусликов) сухих степей и полупустынь (Нельзина и Данилова, 1960). Нападение клещей на хозяев, метаморфоз неполовозрелых фаз, яйцекладка и развитие яиц ограничены теплым периодом года (с апреля по август). В этот промежуток наблюдается последовательная смена нападающих фаз (максимум для личинок в мае, нимф — в июне и имаго — в июле), и на зимовку уходят голодные личинки, остающиеся неактивными весь осенний период, несмотря на достаточно высокую температуру внутри норы.
У обитателей нор с холодным микроклиматом, как в случае с Ixodes crenulatus — паразитом сурков Marmota baibacina centralis в горах Тянь- Шаня на высоте 1800—3200 м над ур. м., жизненный цикл даже при наиболее благоприятных условиях продолжается не менее 3 лет (Кошеч- кина, 1950; Афанасьева, 1951). За летний период успевает развиться только одна фаза и одновременно в гнезде встречаются особи нескольких поколений.
Значительная часть иксодовых клещей с пастбищным типом паразитизма, включая однохозяинных Hyalomma scupense и двуххозяинных Н. plumbeum, Н. detritum, Н. anatolicum, Rhipicephalus bursa, имеет одногодичные циклы. Наиболее детально особенности одногодичного жизненного цикла изучены у Dermacentor pictus (Олсуфьев, 1953), D. таг- ginatus (Покровская, 1951, 1953) и D. nuttalli (Меринов, 1964). Для этих трех видов характерны зимовка и весенняя волна паразитирования имаго, тогда как в летние месяцы у особей, не сумевших найти прокормителей, развивается состояние «неактивности» (Яшкуль, 1960).
Напитавшиеся весной самки вскоре приступают к яйцекладке и уже с июня начинается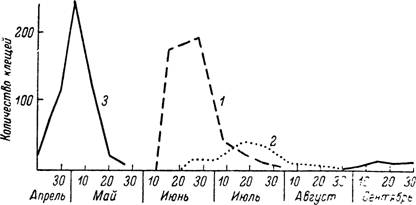
Рис. 155. Сезонность паразитирования различных фаз развития клеща Dermacentor pictus в Московской обл.
1 — средняя численность личинок на пяти полевках; 2 — то же для нимф; 3 — средняя численность половозрелых клещей на одной корове (по Олсуфьеву, 1953).
массовое нападение личинок. Напитавшиеся личинки быстро линяют на нимф, паразитирующих на грызунах с конца июля по август и уже в августе начинающих линять на имаго (рис. 155). Большая часть последних приступает к питанию после перезимовки, но в некоторых районах наблюдается и осенняя волна паразитирования клещей на скоте. В этом случае откладка яиц происходит только у перезимовавших особей (Нецкий и Ольшевская, 1950; Ламанова, 1962).
Общей особенностью видов рода Dermacentor служит короткий период существования в природе голодных личинок и нимф, не способных к длительному голоданию, и исключительная долговечность взрослых клещей. У D. pictus, по наблюдениям Н. Г. Олсуфьева (1953), имаго при отсутствии хозяев переживает в природных биотопах 3—4 года. Вследствие этого активизирующаяся весной или осенью популяция клещей представляет смесь из особей не менее трех поколений.
Сезонная регуляция одногодичного цикла обеспечивается существованием летней неактивности у голодных и диапаузы у напитавшихся самок, изученных экспериментально у D. pictus (Разумова, 1960, 1965, 19666) и у D. marginatus (Белозеров, 1963, 1965а, 19656). Впадение клещей этих видов в состояние летней неактивности и возникновение несколько позднее потенциальной способности к диапаузе у напитавшихся самок служит надежной биологической адаптацией, предотвращающей появление в конце лета и осенью не способных к зимовке неполовозрелых фаз развития. По данным трехлетних наблюдений над природной популяцией имаго D. pictus в Московской обл. (Разумова, 19666), развитие неактивности и потенциальной. способности к диапаузе у напитавшихся самок происходило независимо. Первый процесс происходил с первой-второй декады июня, т. е. на 1—2 недели ранее возникновения потенциальной диапаузы. В то же время летняя неактивность исчезала в первой половине августа — за 1—2 месяца до прекращения состояния потенциальной диапаузы.
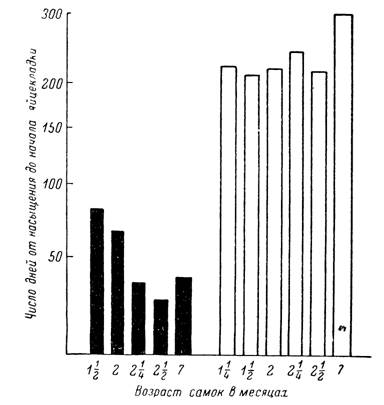
Рис. 156. Средние сроки от насыщения до начала яйцекладки (при 18°) у самок Dermacentor marginatus (волгоградская популяция) в зависимости от возраста и фотопериодических условий их содержания до кормления (по Белозерову, 1965а).
Описанный двойной контролирующий механизм, с одной стороны, предотвращает нежелательное для вида насыщение самок в начале лета, когда у них еще не развилось состояние потенциальной диапаузы. С другой стороны, исчезновение летней неактивности в конце августа также не ведет к неблагоприятным последствиям, так как напитавшиеся осенью самки перезимовывают в состоянии диапаузы и приступают к яйцекладке вместе с особями, напитавшимися весной следующего года.
Черные столбики — 9 час. света; белые столбики — 18 час. света.
Возникновение диапаузы у напитавшихся самок контролируется световым режимом содержания голодных имаго. Так, в условиях эксперимента без- диапаузная откладка яиц происходила только у самок D. marginatus, содержавшихся до питания в условиях короткого дня (9 —12 час. света), а после нахождения при длительном дне (16—24 часа) начало яйцекладки задерживалось на несколько месяцев (рис. 156). Сходный короткодневный тип фото- периодической реакции был свойствен и самкам D. pictus. На прочность диапаузы влияли также температурный режим содержания голодных имаго и возраст самок после линьки. Снятие диапаузы к весеннему периоду обеспечивалось реактивацией самок низкими температурами во время зимовки.
При бездиапаузном развитии сезонность жизненных циклов выражена слабее и возможно частичное налегание последовательных поколений. Подобный тип развития свойствен клещу Hyalomma asiaticum. В пустынях Туркмении, по многолетним наблюдениям А. В. Кочкаревой- Бахаевой (1964) над численностью клещей на хозяевах и в их основном биотопе — норах песчанок, а также на основании наших исследований возрастного состава имаго в природных популяциях (табл. 2), можно считать, что основная масса Н. asiaticum развивается по одногодичному циклу, несмотря на круглогодичное нахождение всех фаз развития на хозяевах и в норах (рис. 157).
В марте—мае наблюдается массовое паразитирование имаго Н. asiaticum на скоте (особенно верблюдах). В июне—августе численность поло-
возрелых клещей на животных и на поверхности почвы становится очень низкой. Основную их массу составляют сильно истощенные особи (табл. 2). Несомненно, что с наступлением летней жары и увеличением дефицита насыщения в припочвенном слое воздуха не нашедшие хозяев клещи в основном погибают. Лишь незначительная часть популяции продолжает нападать на скот в течение всего летнего периода и доживает до осени.
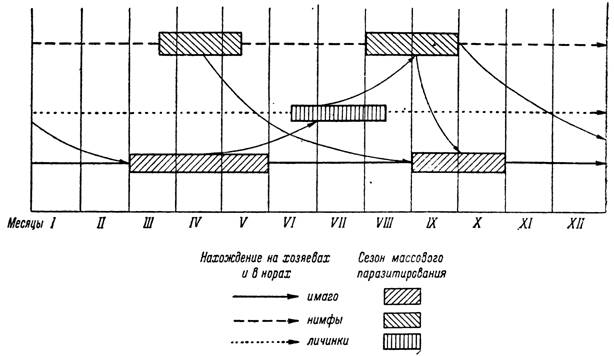
Рис. 157. Цикл развития клеща Нуаіошта asiaticum в Южной Туркмении.
Напитавшиеся в апреле—мае самки в условиях высоких средних температур без задержки приступают к яйцекладке. В июне—августе наблюдается годовой максимум численности активных личинок в норах и на хозяевах. Напитавшиеся личинки уже с августа линяют в нимф и обеспечивают позднелетний подъем численности последних на песчанках. Паразитирование неполовозрелых фаз на песчанках продолжается весь осенне-зимний период, прекращаясь только в наиболее холодное время года — в декабре—январе. Значительная часть популяции нимф, однако,
Таблица 2
Возрастной состав голодных самок Нуаіошта asiaticum, собранных на поверхности почвы в Каракумах в 1955 и 1958 гг.
% особей определенной степени истощения по месяцам
| Степень истощения клещей | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII |
| 2-я | 70 | 58 | 24 | 6 | 4 | 22 | 58 | 59 | ||||
| 3-я | — | — | 11 | 42 | 49 | 19 | 13 | 22 | 25 | 12 | — | — |
| 4-я | 19 | ~ | 27 | 75 | 83 | 56 | 17 | 29 | ||||
| Всего собрано клещей . | — | — | 27 | 159 | 87 | 65 | 23 | 9 | 12 | 17 | — | — |
переживает осенне-зимние месяцы в голодном состоянии и обеспечивает весенний пик численности на грызунах.
Нимфы весеннего питания линяют в течение лета на имаго, но последние в значительной массе остаются в норах до весны. Лишь часть их активизируется и нападает на скот осенью того же года. Нимфы, напитавшиеся в конце лета, вероятно, также успевают перелинять в имаго в том же сезоне или же при осеннем питании их линька может задерживаться низкими температурами до следующей весны. Таким образом, сезонность паразитирования определенных фаз развития Н. asiaticum на хозяевах находится под непосредственным воздействием климатических факторов, а наличие активных голодных особей зависит от доступности прокорми-
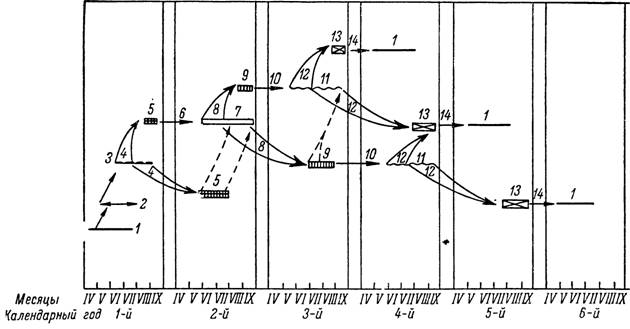
Рис. 158. Цикл развития клеща Ixodes persulcatus в Карелии (по Хейсину, Павловской,
Малаховой и Рыбак, 1955).
1 — голодные самки; 2 — сытые самки; 3 — яйцекладка; 4 — эмбриональное развитие; 5 — вылуп- ление личинок; 6 — переход личинок в активное состояние; 7 — голодные личинки; 8 — питание и метаморфоз личинок; 9 — выход нимф; Ю — переход нимф в активное состояние; 11 — голодные нимфы; 12 — питание и метаморфоз нимф; 13 — выход имаго; 14 — переход имаго в активное состояние.
телей — позвоночных и способности к многомесячному голоданию не только на имагинальной, но и на неполовозрелых фазах (Афанасьева, 1956). В связи с этими биологическими особенностями наблюдается частичное смешение особей двух последовательных генераций в составе одной популяции при сохранении синхронности развития и одногодичном цикле развития у основной массы клещей.
Наиболее сложные и продолжительные жизненные циклы свойственны клещам Ixodes persulcatus и I. ricinus, продвинувшимся на север дальше других видов с пастбищным типом паразитизма. В серии исследований Е. М. Хейсина с сотрудниками (Хейсин, 1955а, 19556; Хейсин и Лебешева, 1955; Хейсин, Бочкарева, Лаврененко и Михайлова, 1955; Хейсин, Бочкарева и Лавренко, 1955; Хейсин, Павловская, Малахова и Рыбак, 1955) установлено, что на территории Карелии развитие каждой фазы этих видов занимает не менее 1 года, но может продолжаться и до
2 лет. Минимальная продолжительность жизненного цикла составляет
3 года, но может достигать 6 лет.
Развитие I. persulcatus протекает следующим образом (рис. 158). Имаго нападают на скот с конца апреля до конца июля с максимумом
паразитирования во второй половине мая—первой половине июня. Отпавшие в мае—июне самки через 2—3 недели приступают к яйцекладке, которая продолжается 1—3 месяца. Из яиц, отложенных до середины июля, в августе—сентябре того же сезона успевают вылупиться личинки, а яйца, отложенные в более поздние сроки, перезимовывают и дают личинок на следующий год. Личинки и нимфы паразитируют на мелких животных в продолжение всего весенне-летнего периода с максимумом численности в конце июня—июля. Неполовозрелые особи, напитавшиеся до середины июля, успевают перелинять в том же сезоне, а отпавшие с хозяев в более поздние сроки впадают в диапаузу и линяют на следующий год. Активация личинок и нимф позднелетнего вылупления или линьки происходит только после перезимовки весной следующего года, а нимфы ранне летней линьки могут активизироваться в том же сезоне. Имаго, перелинявшие в летне-осенний период, остаются неактивными до весны следующего года. Большая часть популяции имеет трехгодичный цикл развития, так как большинство кладок дает личинок в том же году, а для личиночного и нимфального метаморфоза требуется еще по одному году. Однако в более холодных микробиотопах и особенно в неблагоприятные в климатическом отношении годы развитие одной или даже двух фаз не укладывается в 1 год и соответственно развитие части популяции может продолжаться 4 или 5 лет. В Московской обл. (Бабенко, 1956) и Красноярском крае (Бабенко и Рубина, 1961), несмотря на более жаркое лето, минимальная продолжительность развития также составляет 3 года, тогда как на юге Хабаровского края (Моисеенко, 1957) и в Приморском крае (Сердюкова, 1960) личинки могут питаться в год яйцекладки и жизненный цикл сокращается до 2 лет.
Развитие /. ricinus протекает сходным образом, но в связи с его большей теплолюбивостью развитие каждой фазы в Карелии продолжается в основном по 2 года при общей продолжительности цикла в 5—6 лет. При продвижении на юг сроки развития соответственно сокращаются и в Ленинградской обл. большая часть популяции имеет 4-летний цикл (Сердюкова, 1960), хотя возможно и более длительное развитие (Алфеев, 1948). В Московской обл. (Бабенко, 1956) и в Англии (Lees a. Milne, 1951) развитие продолжается 3 года и не исключено, что в южной части ареала оно проходит в еще более сжатые сроки.
Обращает внимание совершенство сезонной регуляции жизненного цикла, обеспечивающей синхронность жизненных явлений у особей одной фазы развития, несмотря на возможную принадлежность к разным поколениям. Последнее обеспечивается существованием яйцевой, личиночной и нимфальной диапауз (Алфеев, 1948; Сердюкова, 1951, 1952), из которых изучены только личиночная и нимфальная (Белозеров, 1964а, 19666; Белозеров и Михайлова, 1966). Температурный порог бездиапауз- ного развития для ленинградской популяции близок к 18°. В условиях длинного дня (18—24 час. света) напитавшиеся личинки без задержки линяют в нимф, а при коротком дне (9—14 час. света) у всех личинок возникала диапауза и метаморфоз заканчивался через 150—200 дней. При 25° наблюдается бездиапаузное развитие независимо от светового режима. Длина дня и температура детерминируют возникновение диапаузы на стадии голодных личинок, но чувствительность к этим факторам сохраняется и у напитавшихся особей, так что в условиях длинного дня и высоких температур всегда наблюдалось бездиапаузное развитие. У личинок из Ленинградской обл. и из Молдавии порог фотопериодической реакции оказался различным и соответствовал особенностям климата этих двух районов. Наряду с другими факторами на возникновение диапаузы влиял и возраст клещей. С увеличением возраста личинок наблюдалось усиление тенденции к возникновению диапаузы.
У нимф, в* отличие от личинок, диапауза характеризуется сменой механизмов регуляции. До питания она детерминирована в соответствии с нормами короткодневной реакции, а после питания — нормами длиннодневной реакции. Описанный механизм возникновения диапаузы делает понятным задержку метаморфоза при позднелетнем питании личинок и уход на зимовку большинства напитавшихся за летний сезон нимф. Перезимовка диапаузирующих особей не вызывала их реактивации, и они линяли в нихмф или имаго одновременно с особями весеннего кормления текущего года. Последнее обеспечивало сохранение сезонности паразитирования при многолетнем цикле и разновозрастных популяциях. У нимф сезонность паразитирования, возможно, регулируется угнетением агрессивности в условиях короткого дня. Однако подобная реакция наблюдалась только у особей не старше 5—(^месячного возраста.
Еще по теме СЕЗОННОСТЬ И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНЕННЫХ циклов:
- Общие особенности
- Описания жизненных циклов
- Синхронизация жизненных циклов с сезонными климатическими ритмами
- Природная очаговость клещевых инфекций
- Введение ...................................................................................................................... 5
- СЕЗОННОСТЬ И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНЕННЫХ циклов
- ИНДИВИДУАЛЬНАЯ ДОЛГОВЕЧНОСТЬ И ФИЗИОЛОГИЧЕСКИЙ ВОЗРАСТ
- ПРОДОЛЖИТЕЛЬНОСТЬ ПИТАНИЯ
- ГОНОТРОФИЧЕСКИЕ ОТНОШЕНИЯ
- ЗАКЛЮЧЕНИЕ
- Введение
- УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
- Род Ornithodorus С. Ъ. Koch
- Питание блох.
- Плазмодии малярии