Синхронизация жизненных циклов с сезонными климатическими ритмами
Анализ данных о продолжительности и механизмах регуляции жизненных циклов у разных видов иксодовых клещей свидетельствует об их высокой лабильности. У видов с достаточно обширными ареалами в зависимости от климатических условий их местообитаний географические популяции могут различаться по таким важным параметрам, как фенология активности, зимующие стадии, механизм индукции диапаузы, время развития одного поколения.
Продолжительность жизненного цикла у некоторых видов может варьировать даже, для потомства одной самки, что создает исключительную возрастную гетерогенность клещевых популяций.Оптимальные гигротермические условия для непрерывного развития иксодовых клещей в течение всего года, по-видимому, обеспечиваются влажным тропическим климатом. В субтропическом климате чрезмерно жаркий и сухой летний сезон или же холодный зимний могут замедлять или приостанавливать развитие, а в умеренном климате зимний перерыв в развитии иксодид становится правилом. Как и у насекомых, жизненные циклы иксодовых клещей приспособлены к особенностям климата их местообитаний. В условиях сезонных климатических ритмов периоды активной жизнедеятельности прерываются периодами покоя, особой формой которого является диапауза. На этой стадии у клещей повышается устойчивость к неблагоприятным факторам среды (низкие или высокие температуры, дефицит влажности) и благодаря существенному снижению уровня метаболизма удлиняются сроки жизни голодающих особей.
В. Н. Белозеров (1981) различает у иксодовых клещей 2 формы диапаузы. При морфогенетической диапаузе наблюдается задержка развития отложенных яиц, уд-Фотопериодическая регуляция развития и поведения иксодовых клещей
| Вид | Диапау- зирую- щая фаза | Процессы, регулируемые фотопериодом | Тип реакции | Литературный источник |
| АтЫуотта атегісапит | Л, Н, И | Подстерегание, линька, оогенез | да | Barnard et al., 1985; Barnard, 1988; Pound, George, 1988 |
| А. triguttatum | Н | Линька | да | Guglielmone, Moorhouse, 1986 |
| A. variegatum | И | Оогенез | да | Pegram et al., 1988 |
| Dermacentor albipictus | л | Нападение | кд | Wright, 1971 |
| D. andersoni | н | Линька | дд | Pound, George, 1991 |
| D. marginatus | и | Нападение, оогенез | кд | Белозеров, 1981 |
| D. niveus | и | То же | кд | Jiang Zai-jie et al., 1991 |
| D. reticulatus | и | » » | кд | Белозеров, 1981 |
| D. silvarum | и | Задержка питания | кд | Белозеров, 1973 |
| D. sinicus | и | Нападение, оогенез | кд | Jiang Zai-jie et al., 1991 |
| D. variabilis | л, н | Нападение | да | Smith, Cole, 1941 |
| Haemaphysalis concinna | л, н, и | Нападение, линька | дд | Белозеров, 1988 |
| H. longicomis | л, н, и | То же | дд | Белозеров, 1988; Sutherst, Bourne, 1991; Fujimoto, 1995a |
| Hyalomma anatolicum | н, и | Линька, оогенез | дд | Белозеров, 1981 |
| Ixodes ricinus | л, н, и | Подстерегание, линька, оогенез | дд, кд- дц, кд | To же |
| /. persulcatus | л | Линька | дд | Белозеров, 1988; Fujimoto, 1995b |
| /. nipponensis | и | Нападение | дд | Fujimoto, 1996 |
| Rhipicephalus appendiculatus | и | » | да | Rechav, 1981 |
| R. pumilio | л, н | Нападение, линька | дд, кд | Белозеров, 1988 |
| R. t uranic us | и | Подстерегание | дд | To же |
Примечание. Л — личинка, Н — нимфа, И — имаго. ДЦ — длиннодневная, КД — корот- кодиевиая, КД-ДД — двухступенчатая реакция короткодиевио-длиииодиевиого типа.
линение периода подготовки к линьке у напитавшихся личинок, нимф или оогенеза у самок. Поведенческая диапауза проявляется в отсутствии агрессивности (способности к нападению и питанию) у голодных клещей. Этот тип диапаузы может возникнуть сразу после вылупления или линьки и связан с уходом на зимовку, как в случае с имаго I. persulcatus и D.
andersoni, или же с переживанием летнего сезона голодными личинками клещей Н. detritum или D. albipictus. В описанных случаях поведенческая диапауза накладывается на стадию послеличиночного до- развития. Последняя затягивается на несколько недель или месяцев, и для активации клещей необходимо сочетание определенных температурных условий и длины светового дня. У имаго D. reticulatus, D. marginatus и некоторых других видов, голодные имаго которых живут 2 года и более, поведенческая диапауза возможна после каждого периода активности и может повторяться в жизни одной особи 2 и даже 3 раза (Cerny et al., 1982; Разумова, 1986).В согласовании во времени циклов развития иксодид с годовыми климатическими ритмами решающая роль принадлежит соотношению длины дня и ночи в суточном цикле (фотопериод). Реакцию организма на фотопериод, индуцирующую или предотвращающую развитие диапаузы, называют фотопериодической реакцией, и она обнаружена у многих видов иксодид (табл. 6). У клещей обнаружены длин-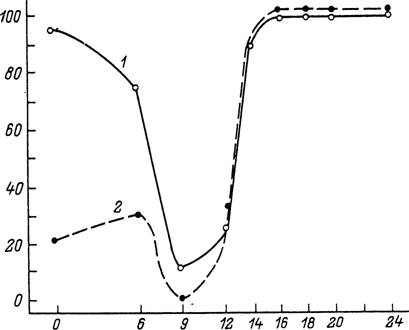
Рис. 62. Индукция репродуктивной диапаузы у напитавшихся самок Dermacentor marginatus (1 — из Дагестана, 2 — из Казахстана) в зависимости от фотопериодических условий (при 18 °С) содержания самок до
кормления (по: Белозеров, 1981).
По оси абсцисс — продолжительность фотопериода, в ч; по оси ординат — диапаузирующие самки, в %.
нодневные фотопериодические реакции, когда активность или развитие стимулируется длинным днем, короткодневные реакции с противоположным эффектом и двухступенчатые реакции при реагировании на короткий и длинный день у нимф I. ricinus. Важной экологической характеристикой для организмов, обладающих фотопериодической реакцией, служит величина критического или порогового фотопериода, при которой соотношение особей с бездиапаузным развитием и диапаузой примерно одинаково.
У клещей с длиннодневной фотопериодической реакцией, как например у нимф Н. anatolicum, при фотопериоде короче критического (13 ч) возникает Диапауза.
У видов с короткодневной фотопериодической реакцией, как например у напитавшихся самок D. marginatus, репродуктивная диапауза наступает при длине дня более 13 ч (рис. 62). Величина критического фотопериода может варьировать у разных географических популяций одного вида, увеличиваясь с юга на север. Например, у напитавшихся самок /. ricinus из Молдавии она составляет 15—16 ч света, а у личинок из Ленинградской области — 17—18 ч (рис. 63). Напротив, у D. marginatus этот показатель не был подвержен географической изменчивости и был равен 13 ч у популяций из Дагестана, Поволжья и Казахстана. При решающей роли фотопериода конечный результат фотопериодической реакции может зависеть от температуры (/. ricinus, Н. anatolicum), от возраста голодных особей (D. reticulatus, I. ricinus) и от других факторов (Белозеров, 1988).Важной особенностью иксодид, обладающих многолетними циклами развития, является возможность повторного и независимого возникновения диапаузы на разных стадиях онтогенеза и в разные годы жизни клеща. Другой их особенностью является, в отличие от насекомых, частое совмещение на одной и той же стадии развития способности к восприятию фотопериода и ответа на него, как обнаружено у напитавшихся личинок и нимф /. ricinus или нимф и самок Н. anatolicum. Восприятие фотопериода на одной стадии развития, а реализация — на другой у иксодид встречаются реже, и примером этого могут служить голодные и напитавшиеся самки D. marginatus.
С возникновением диапаузы у клещей возникает повышенная устойчивость к неблагоприятным внешним условиям и особенно температуре. Наряду с по-
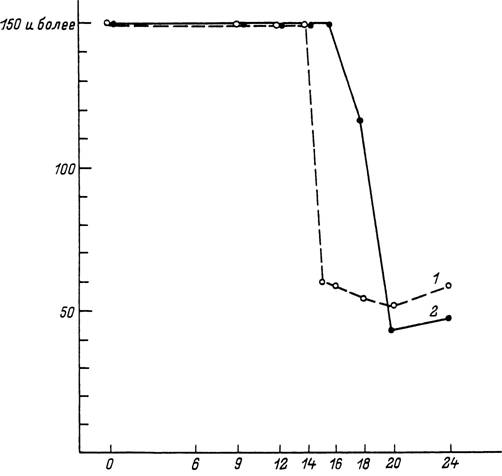
Рис. 63. Продолжительность метаморфоза у напитавшихся самок Ixodes ricinus (1 — из Молдавии, 2 — из Ленинградской области) при 18 °С и различных фотопериодах. Голодные личинки содержались при режиме 12 ч света—12 ч темноты (по: Белозеров, 1981).
По оси абсцисс — продолжительность фотопериода, в ч; по оси ординат — продолжительность метаморфоза, в сут.
нижением уровня метаболизма и экономным расходованием энергетических резервов организма клещ в этот период может проходить холодовую реактивацию.
Последняя обеспечивает прекращение диапаузы в конце зимовки и стимулирует высокую активность перезимовавших голодных особей или яйцекладку и линьку у напитавшихся. Процесс холодовой реактивации достаточно длинен, и для ее завершения у напитавшихся самок D. marginatus при температуре 0—6 °С требуется 3—4 мес.Наиболее полно исследована зимняя диапауза, характерная для иксодовых клещей, обитающих в регионах с достаточно холодной и продолжительной зимой. Стадии, на которых осуществляется зимовка, и факторы, индуцирующие диапаузу, в определенной степени видоспецифичны. Например, у /. persulcatus в пределах обширнейшего ареала этого вида самки, перелинявшие со второй половины лета, всегда остаются в состоянии облигатной диапаузы до весны следующего года. Напротив, у таких видов, как I. ricinus, постоянным признаком является лишь способность к возникновению диапаузы практически на всех стадиях развития. Конкретная же реализация этого признака определяется фотопериодом, метеорологическими условиями и физиологическим возрастом самих клещей. В результате реальное число стадий, на которых возникла диапауза, и общая продолжительность жизненного цикла могут быть различными не только для разных географических популяций, но даже для особей из потомства одной самки. Очевидно, что жизненные циклы иксодовых клещей представляют одну из важнейших адаптаций к их условиям существования. По аналогии с насекомыми мы полагаем, что более простыми следует считать
гомодинамные или, как их называют у клещей, «непрерывные» циклы развития, свойственные некоторым тропическим и субтропическим видам. Более сложные многолетние циклы с диапаузирующими стадиями мы рассматриваем как вторичные, возникшие в связи с приспособлением клещей к сезонным климатическим циклам умеренной зоны. Процессы, определяющие эволюцию жизненных циклов у клещей, исключительно лабильны и допускают возможность быстрого реагирования на флюктуации климата за счет перестройки жизненных циклов по их продолжительности, числу хозяев в развитии и срокам активности отдельных стадий.
Табл. I. Внешний вид клеща Hyalomma cisiciticum.
1 — голодная самка; 2 — нимфы, питающиеся на ушастом еже; 3 — самки, питающиеся на спине кролика; 4 — напитавшаяся и отпавшая с хозяина самка.
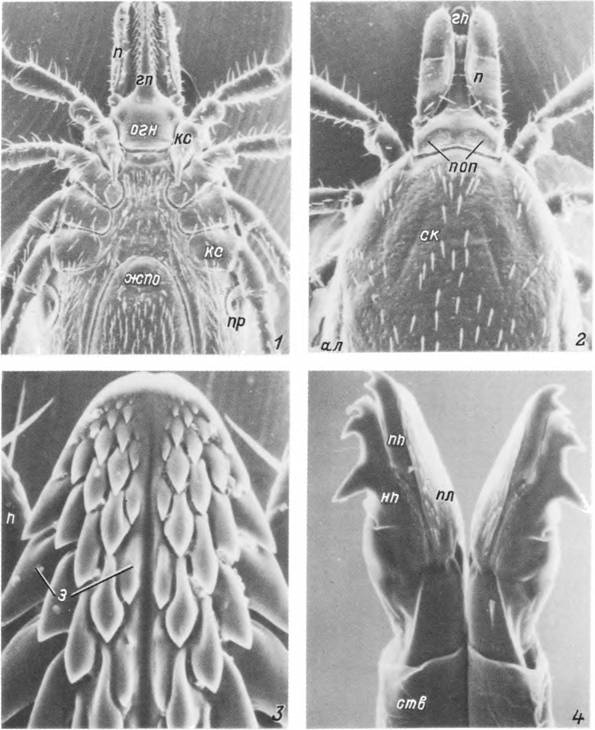
Табл. II. Самка Ixodes persulcatus.
1,2 — передняя половина тела с вентральной (/) и дорсальной (2) стороны; 3 — дистальный конец гипостома; 4 — передние концы хелішер. ал — аллоскутум; гп — гипостом; жпо — женское половое отверстие; j — зубцы гипостома; кс — кокса; нп — неподвижный палец хелицеры; огн — основание гнатосомы; п — пальпа; пл— плащ; поп — поровое поле; пп — подвижный палец хелицеры; пр — перитрема; ск — скутум; ств — ствол хелицеры.
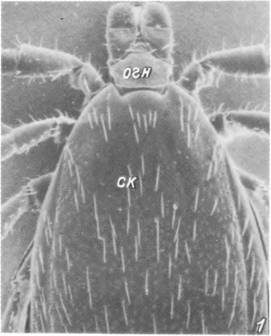
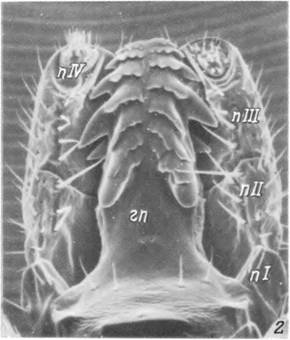
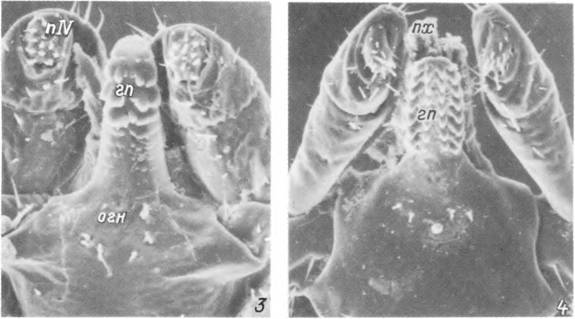
Табл. III. Строение гнатосомы у видов Ixodes.
1,2 — самец 1. persulcatus: 1 — передняя половина тела с дорсальной стороны, 2 — гнатосома с вентральной стороны; 3, 4 — гнатосома самца (і) и самки (4) 1. trianguliceps с вентральной стороны, гп — гипостом; огн — основание гнатосомы; пі—пГѴ— I—IV членики пальп; пх — палец хелицеры; ск — скутум.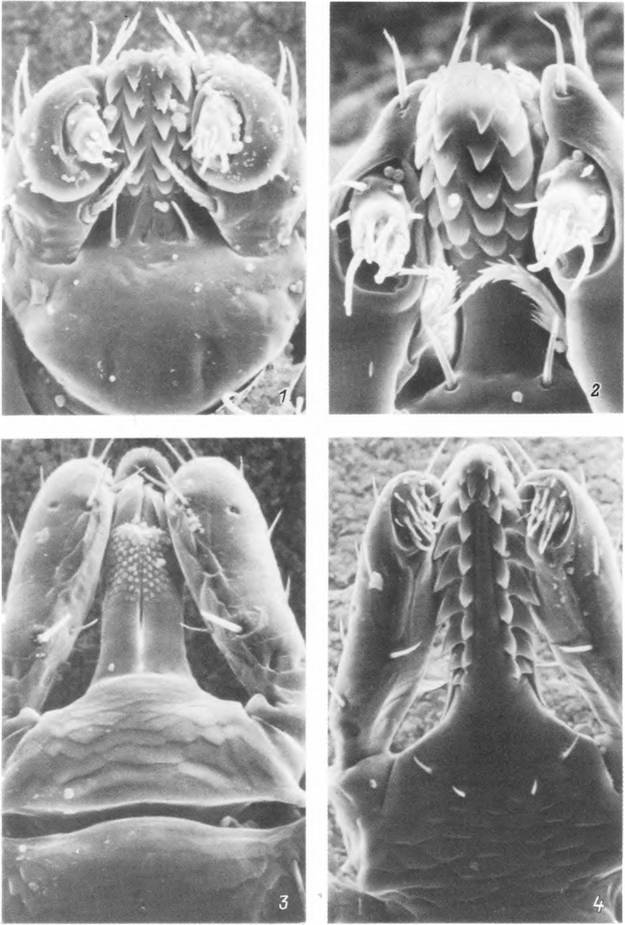
Табл. IV. Гнатосомы личинок.
1.2 — Boophilus annulatus (У) и Rhipicephalus turanicus (2) с вентральной стороны; 3, 4 — Ixodes ricinus с дорсальной (J)
и вентральной (4) стороны.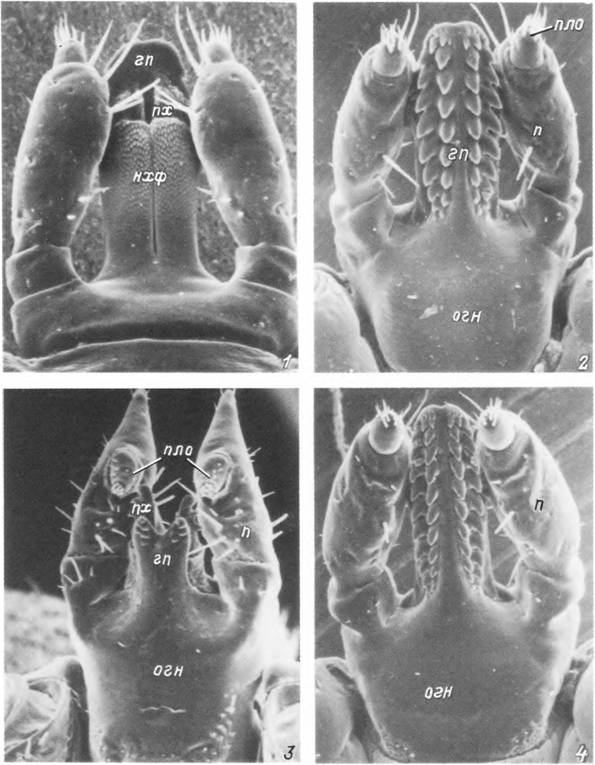
Табл. V. Строение гнатосомы у клеща Ixodes игіае.
1,2 — гнатосома нимфы с дорсальной (/) и вентральной (2) стороны; 3, 4 — гнатосома с вентральной стороны самца (і) и самки (4). гп — гипостом; нхф — наружные хелицеральные футляры; огн — основание гнатосомы; п — пальпа; пло —
пальпальный орган; пх— палец хелицеры.
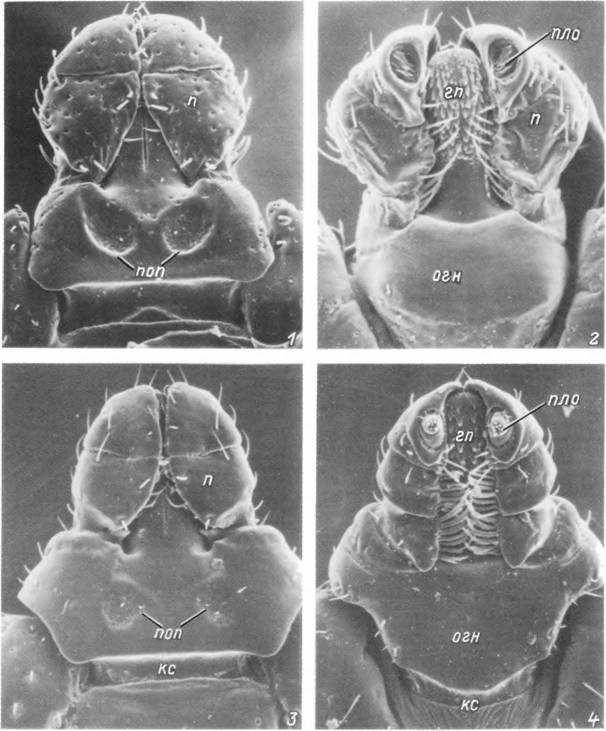
Табл. VI. Гнатосомы самцов и самок.
У. 2 — самка Dermacentor reticulatus с дорсально» (У) и вентральной (2) стороны; 3, 4 — самец Rhtpicephalns t»ran>cus с дорсальной (3) и вентральной (4) стороны, гп — гипостом; кс — камеростомальная складка; огн — основание гнатосомы; п — пальпа; пяо — пальпальный орган; поп — поровое поле.
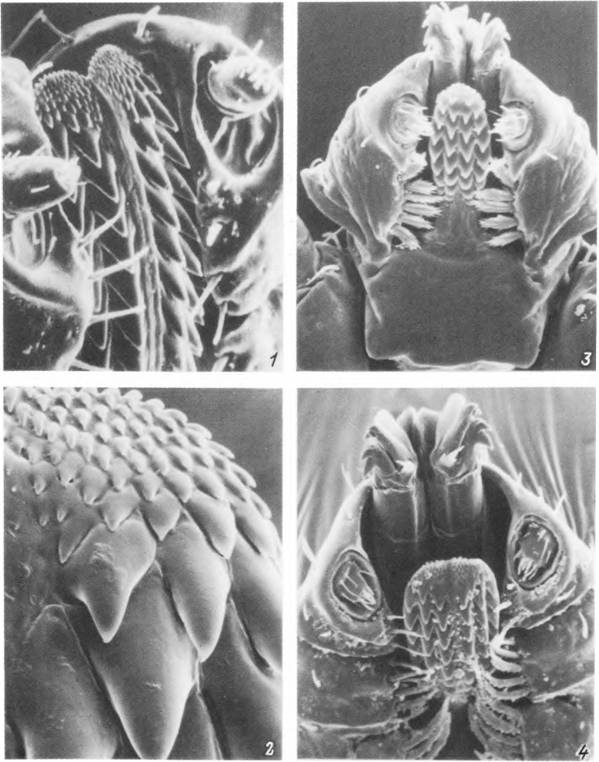
Табл. VII. Гнатосомы иксодовых клешей с вентральной стороны.
1,2 — самка Нуаіотта asiuticum: 1 — внешний вид гнатосомы, 2 — дистальный конец гипостома; 3 — нимфа Haemaphy- salis punctata с выдвинутыми хелицерами; 4 — то же, самец Rhipicephalus turanicus.
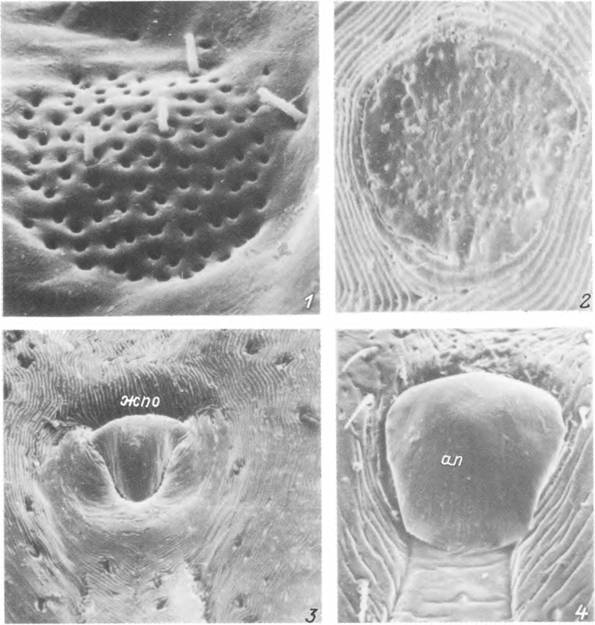
Табл. VIII. Детали поверхности тела Hyalomma asiaticum.
/—і — самка: I — поровое поле, 2 — фовеа, 3 — генитальное отверстие; 4 — генитальное отверстие самца, ап — апрон;
жпо — женское половое отверстие.
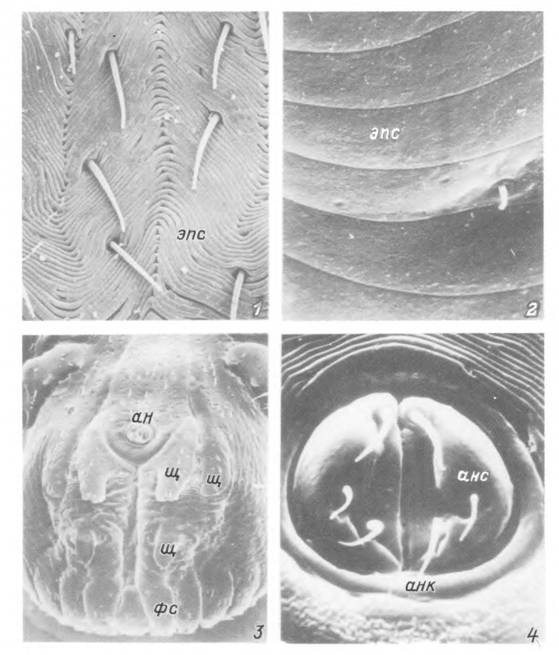
Табл. IX. Детали поверхности тела Hyalomma asiaticum.
1,2 — участок растяжимой кутикулы голодной (7) и напитавшейся (2) нимфы; 3 — задняя половина тела питающегося самца с вентральной стороны; 4 — анальное отверстие нимфы, ан — анальное отверстие; анк — анальное кольцо; анс — анальная створка; фс — фестоны; эпс — эпнкутнкулярные складки; щ — вентральные кутикулярные щитки.

Табл. X. Поверхность кутикулы, щетинки и отверстия дермальных желез на поверхности идиосомы Ixodes
persulcatus.
1,2 — участок поверхности аллоскутума со складками эпикутикулы, отверстиями дермальных желез и механорецепторнымн щетинками при малом и большом увеличениях; 3 — твердая кутикула в области конскутума и краевого валика самца; 4 — кончик щетинки краевого валика самца; 5 — складчатость эпикутикулы, выводное отверстие дермальной железы и щетинки аллоскутума личинки; 6 — кончик щетинки аллоскутума личинки при большом увеличении.
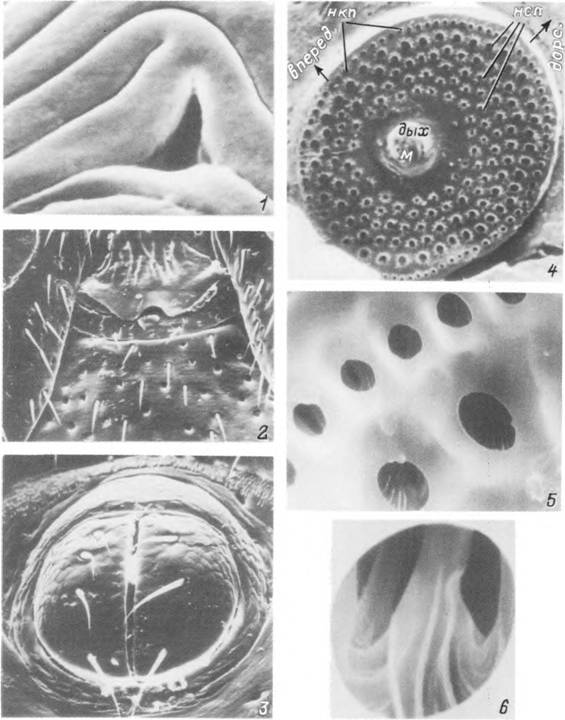
Табл. XI. Детали поверхности идиосомы Ixodes persulcatus.
1 — отверстие дермальной железы II типа на аллоскутуме самки; 2 — генитальное отверстие самца; 3 — анальное отверстие самца; 4 — правая перитрема самки; 5 — наружные краевые п срединные поры перитремы самки: 6 — перегородчатые структуры в глубине срединной поры, дых — дыхательное отверстие; м — макула; нкп и неп — наружные краевые и срединные поры.
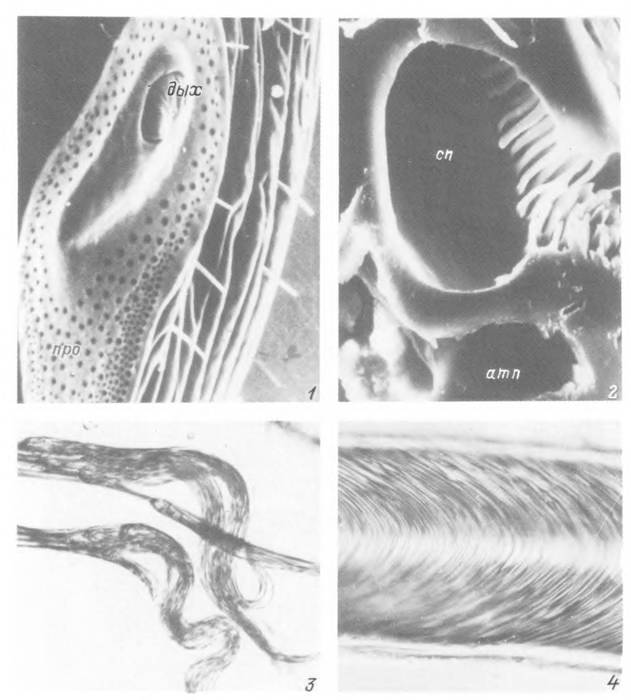
Табл. XII. Органы дыхания самца Hyalomma asiciticum.
1 — внешни» вид перитремы; 2 — срез через участок перитремы; 3 — трахеолы у окончания трахейного ствола; 4 — участок трахейного ствола, дых — дыхальце; про — поровые отверстия перитремы; атп — атриальная полость; сп — стигмальная
полость.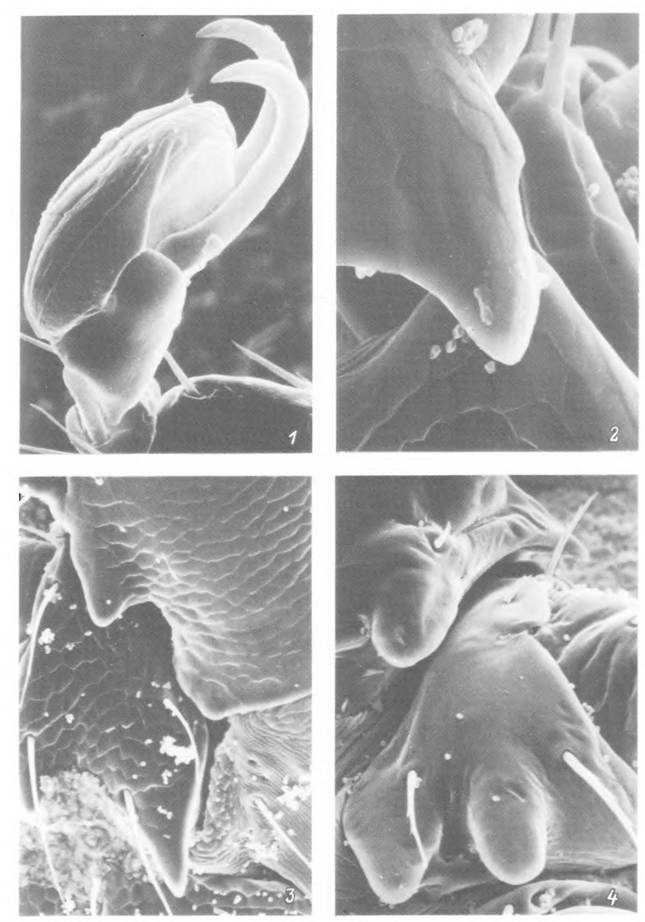
Табл. XIII. Кутикулярные структуры, участвующие в удержании клеща в шерсти хозяина.
1 — лапка I пары ног личинки Ixodes ricinus] 2 — дорсальная корнуа нимфы Ixodes ricinus; 3 — вентральная корнуа и кок- сальный шип нимфы Ixodes ricinus-, 4 — корнуа и коксальный шип нимфы Dermacentor reticulatus.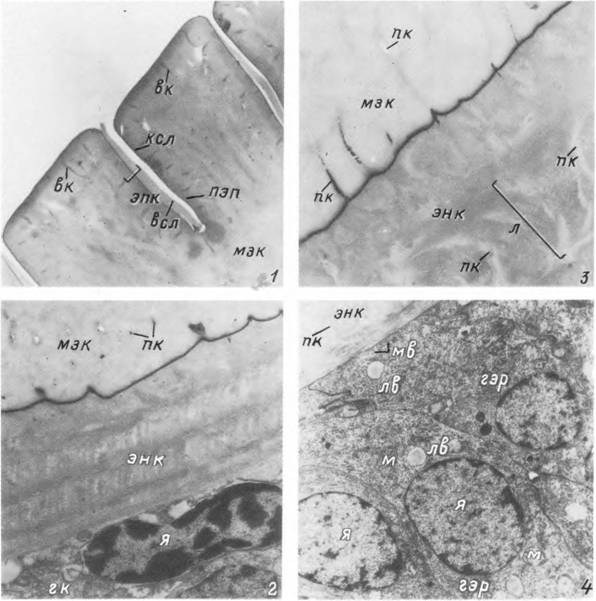
Табл. XIV. Участок растяжимых покровов нимфы Ixodes persulcatus.
1 — поверхность и апикальная часть кутикулы голодной особи: 2 — гиподерма в период голодания; 3 — базальная часть кутикулы голодной особи: 4 — гиподерма в период питания, вк — восковой канал: всл — восковой слой; гк — гиподермальная клетка; гэр — гранулярный эндоплазматический ретикулум: ксл — кутикулиновый слой; л — ламелла прокутикулы: лв — липидное включение; м — митохондрия; мв — микроворсинки; мж — мезокутикула: пк — поровые каналы; пэп — протеиновая эпикутикула; энк — эндокутикула; эпк — эпикутикула: я — ядро.
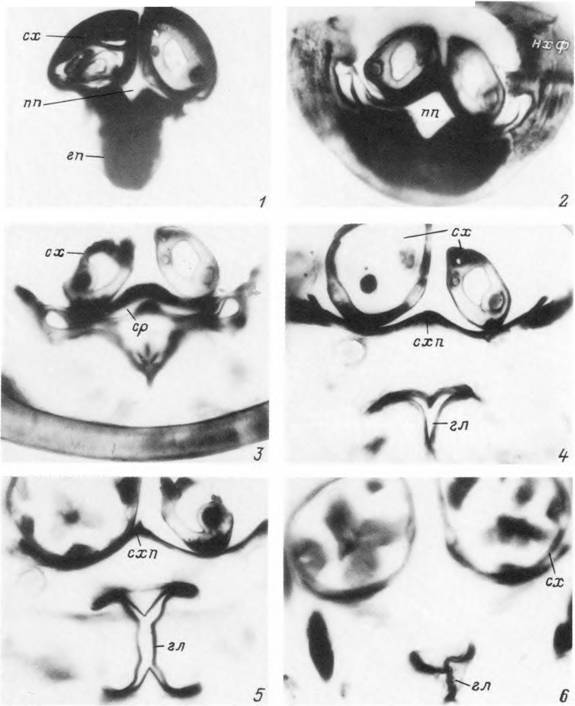
Табл. XV. Ротовые органы и глотка нимфы Ixodes persulcatus на поперечном срезе.
I — дистальный конец хоботка; 2 — средняя часть хоботка на уровне слияния слюнного резервуара п предротовоп полости; 3 — проксимальная часть предротовой полости п слюнной резервуар; 4 — начало глотки; 5 — средняя часть глотки; 6 — задний конец глотки, гл — глотка; гл — піпостом; ихф — наружные хелнцерапьные футляры; пп — предротовая полость; ср — слюнной резервуар; сх — ствол хелицеры; схп — субхелицеральная пластинка.
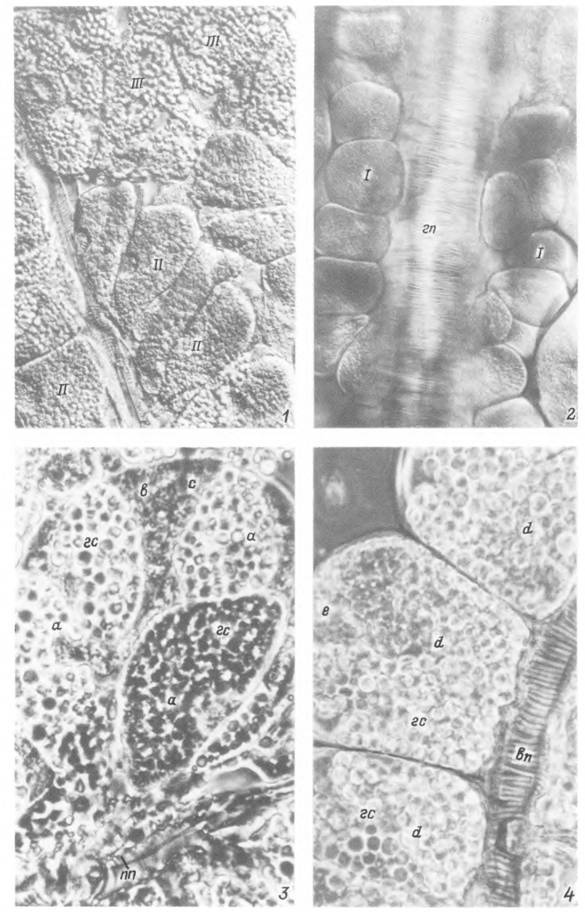
Табл. XVI. Прижизненные фотографии альвеол слюнной железы голодной самки Hyalomma asiaticum.
1 — группа альвеол II и III типов; 2 — альвеолы I типа на главном выводном протоке; 3 — альвеола II типа; 4 — альвеолы III типа (7, 2 — интерференционный контраст; 3,4 — фазовый контраст). 1 — альвеолы I типа; II— альвеолы II типа; 111 — альвеолы III типа, вп — вторичный выводной проток слюнной железы; гп — главный выводной проток; а — клетки типа а; b — клетки типа Ь\ с — клетки типа с; d — клетки типа d\ е — клетки типа е\ гс — гранулы секрета; пп — полость протока.
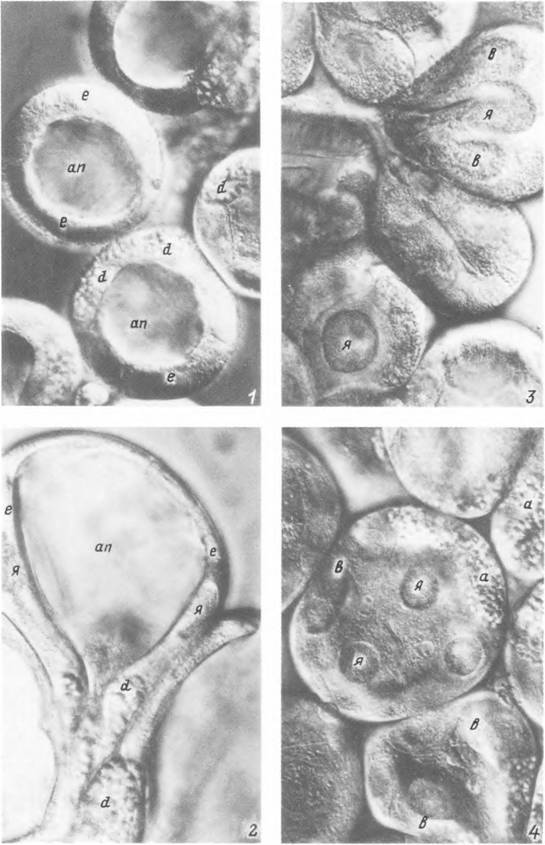
Табл. XVII. Прижизненные фотографии альвеол слюнной железы питающейся самки Hyalamma asiaticum
(интерференционный контраст).
I — альвеолы 111 типа в начале питання; 2 — то же, в конце питания; 3 — альвеолы II типа в начале питания; 4 — то же,
в конце питания. Обозначения как на табл. XVI.
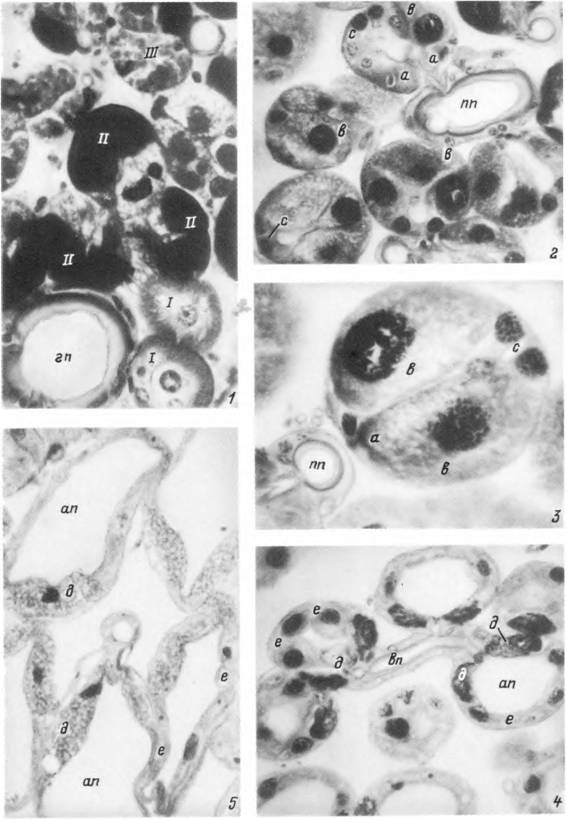
Табл. XVIII. Срезы через альвеолы слюнной железы самки Hyalomma asiaticum (окраска железным гематоксилином-эозином).
У — участок железы вокруг главного выводного протока голодной особи: 2 — альвеолы II типа в начале питания: і — то же, в конце питания: 4 — альвеолы III типа в начале питания: 5 — то же, в конце питания. Обозначения как на табл. XVI.
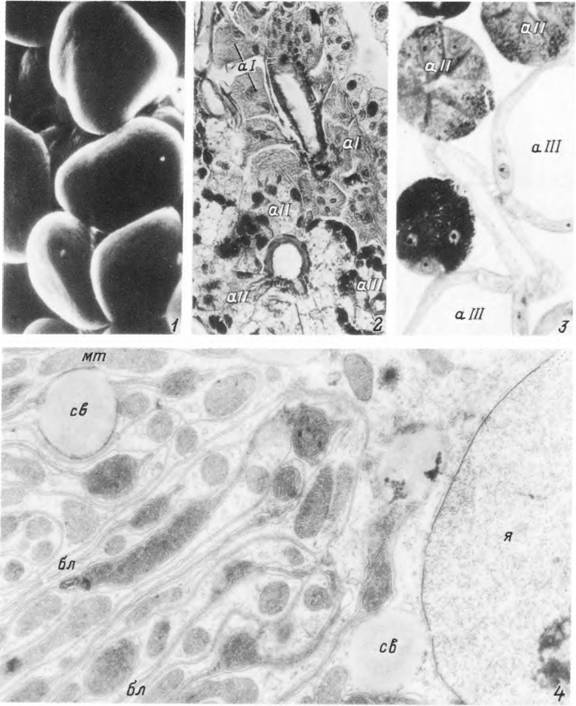
Табл. XIX. Слюнные железы самки Ixodes persulcatus.
1 — альвеолы III типа питающейся особи в растровом электронном микроскопе; 2, 3 — гистологический срез через участок железы голодной (2) и питающейся (2) особи; 4 — электронограмма участка альвеолы I типа, аі— альвеолы I типа; all— альвеолы II типа; аШ— альвеолы III типа; бл — базальный лабиринт; мт — митохондрии; са — секреторная вакуоль; я — ядро.
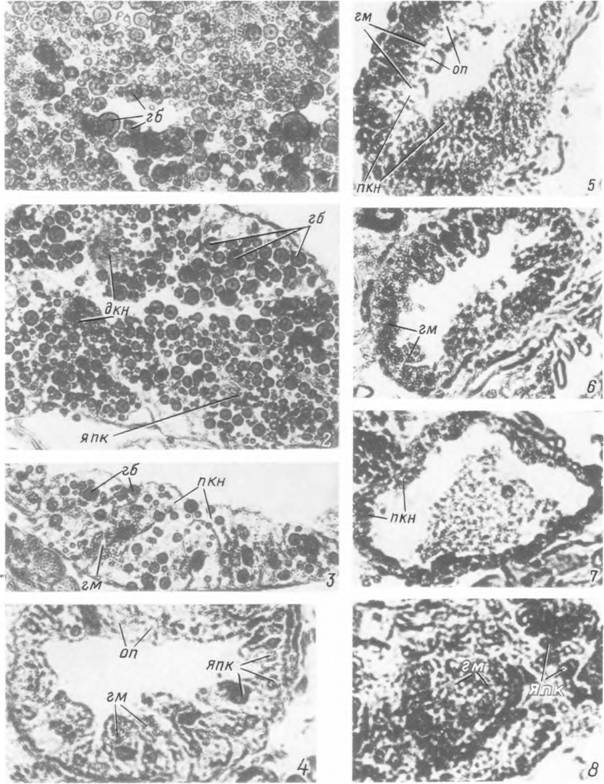
Табл. XX. Изменения в клетках дивертикулов средней кишки самки Ixodes persulcatus при старении
клешей.
7—3 — особи I физиологического возраста на стадии послелиночного доразвития; 4, 5 — особи II возраста вскоре после активации; 6 — особи III возраста в середине периода активности; 7, 8 — особи IV физиологического возраста в конце жизни, гб — внутриклеточные включения гемоглобина; гм — кристаллики гематина; дкн — дегенерирующие кишечные клетки нимфальной фазы; on — оптические пустоты на месте растворившихся липоидных включений; пкн — пищеварительные клетки нимфальной фазы: япк — ядра пищеварительных клеток.
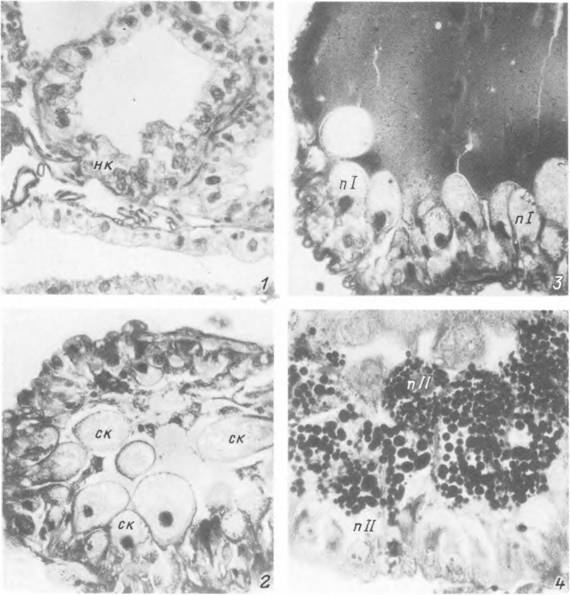
Табл. XXI. Поперечные срезы через дивертикулы средней кишки самок Ixodes persulcatus на разных сроках питания.
1 — голодная особь; 2 — 1-е сутки питания; 3 — 5-е сутки питания; 4 — полностью напитавшаяся особь перед началом яйцекладки, нк — недифференцированные кишечные клетки; пі и nil— пищеварительные клетки I и II типов; ск — секреторные клетки.
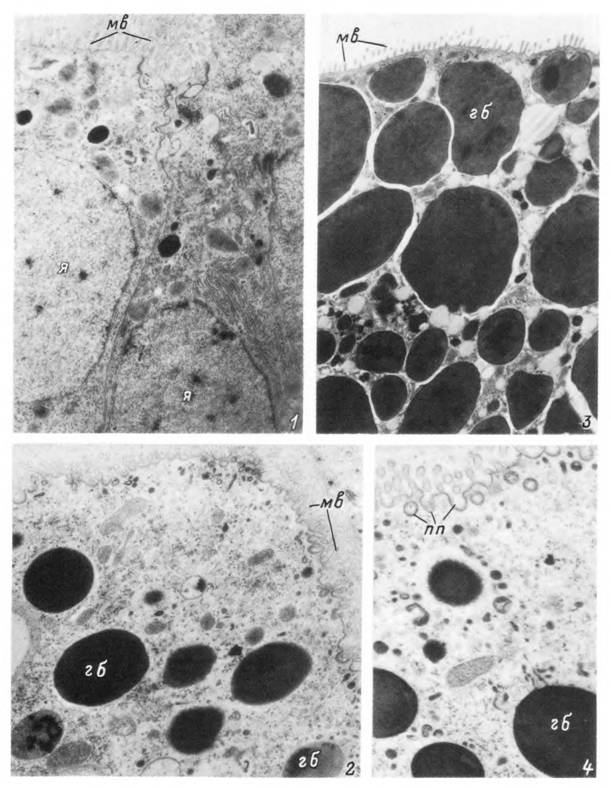
Табл. XXII. Ультраструктура кишечных клеток питающейся самки Ixodes persulcatus.
I — апикальная часть секреторной клетки; 2 — апикальная часть пищеварительной клетки I типа; 3 — апикальная часть пищеварительной клетки II типа; 4 — пиноцитоз на поверхности пищеварительных клеток II типа, гб — включения гемоглобина; мв — микроворсинки; пп — формирующийся пнноцитознын пузырек.
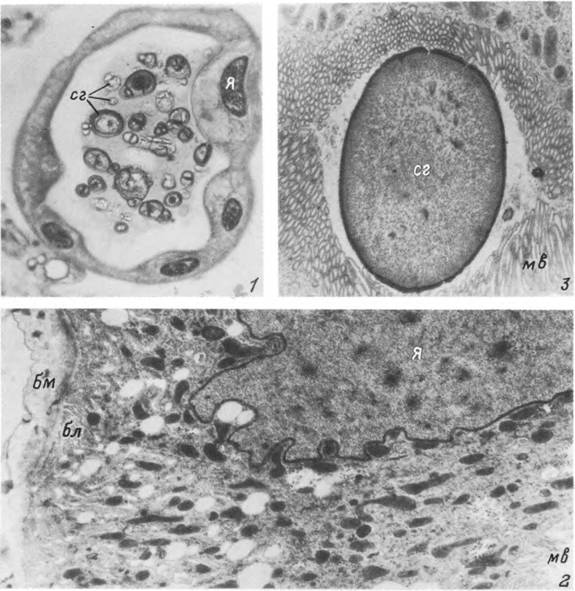
Табл. XXIII. Мальпигиев сосуд питающейся самки Ixodes persulcatus.
/ — поперечный срез в средней части сосуда; 2 — участок стенки сосуда; 3 — сферит гуанина в просвете сосуда, бл — базальный лабиринт; бм — базальная мембрана; мв — микроворсинки; сг — сфериты гуанина; я — ядро.
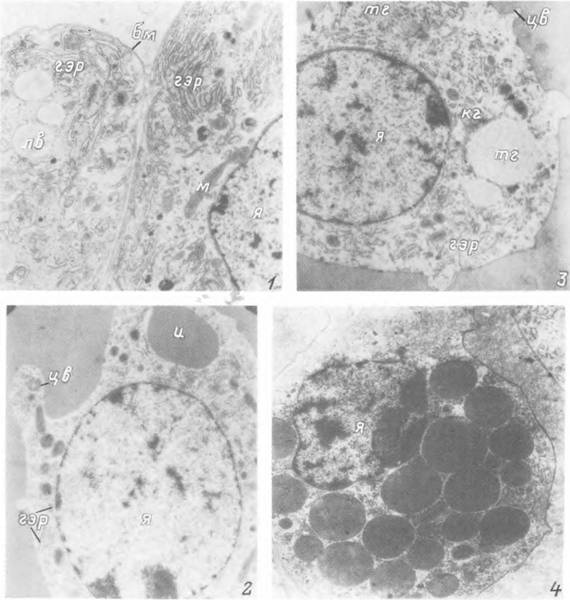
Табл. XXIV. Ткани внутренней среды нимфы Ixodes persulcatus.
1 — клетки жирового тела питающейся особи; 2 — плазматоцит; 3 — гранулоцит I типа; 4 — гранулоцит II типа, бм — базальная мембрана; гэр — гранулярный эндоплазматический ретикулум; и — инвагинация поверхности клетки; кг — комплекс Гольджи; лв — липидное включение; м — митохондрии; тг — тубулярная гранула; цв — цитоплазматический вырост; я — ядро.
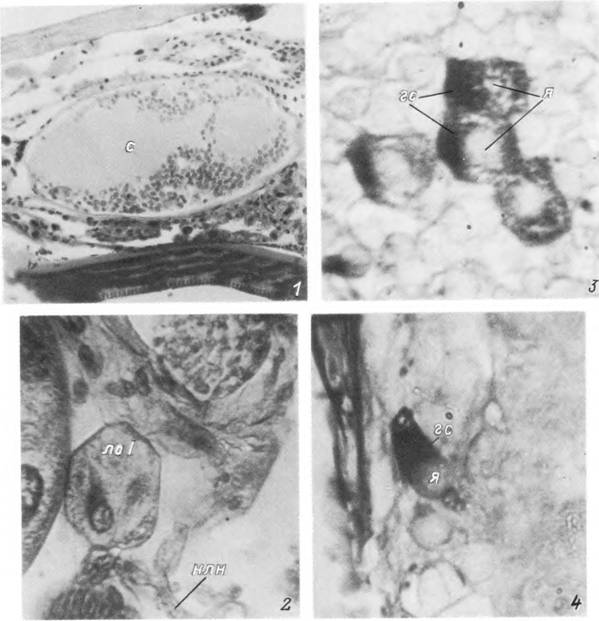
Табл. XXV. Нейроэндокринная система самки Ixodes persulcatus.
1 — синганглий; 2 — латеральный орган I пары; 3, 4 — секреторные клетки I и II типов, гс — гранулы секрета; лоі — латеральный орган; нлн — наружный латеральный нерв; с — синганглий; я — ядро.
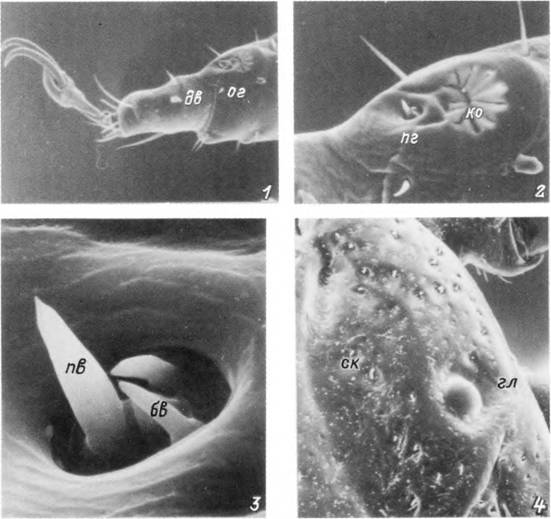
Табл. XXVI. Органы чувств самки Hyalomma asiaticum.
1 — тарзальный членик I пары ног; 2 — внешний вид органа Галлера; 3 — передняя группа сенсилл органа Галлера; 4 — глаз на спинном щитке, бв — бороздчатые волоски; гл — глаз; дв — дистальные волоски; ко — капсула органа Галлера; ог — орган Галлера; пв — пористый волосок; пг — передняя группа волосков; ск — скутум.
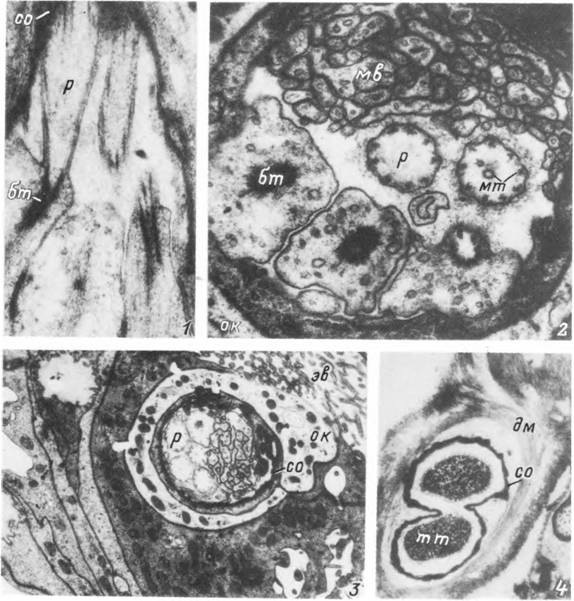
Табл. XXVII. Строение клеточных элементов хемо- и механорецепторных сенсилл Ixodes persulcatus.
1 — продольный срез периферических отростков в районе отхождения ресничек; 2 — поперечный срез в районе начала ресничек; 3 — общий вид клеточных элементов сенсиллы; 4 — трубчатые тельца механорецепторов, бт — базальное тельце; дм — дополнительная волокнистая мембрана; мв — микроворсинки; мт — микротрубочки; ок — оберточная клетка; р — ресничка; со — сколопоидная оболочка; тт — трубчатое тельце; эв — экстраклеточная вакуоль.
п
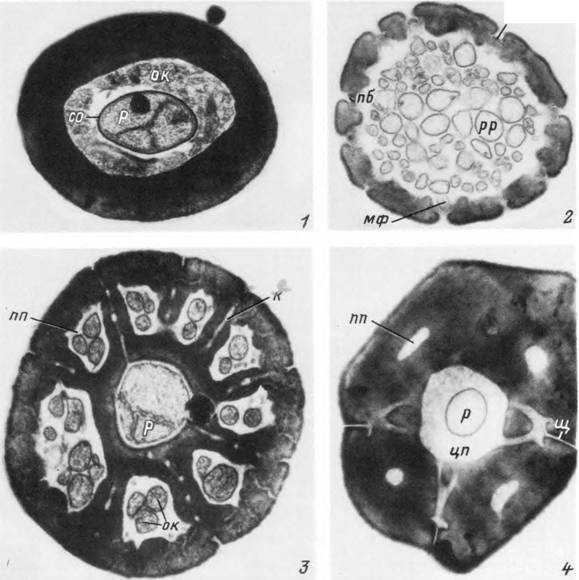
Табл. XXVIII. Поперечные срезы волосков сенсилл Ixodes persulcatus.
I — вкусовой волосок; 2 — пористый волосок; 3 — бороздчатый волосок; 4 — посткапсулярный волосок, к— каналец; мф — микрофнламенты; ок — отростки оберточной клетки; п — пора; пб — поровая бляшка; пп — периферическая полость;/;— ресничка;/;/; — разветвленная ресничка; со — сколопондная оболочка; цп — центральная полость; щ — щелевидная полость.
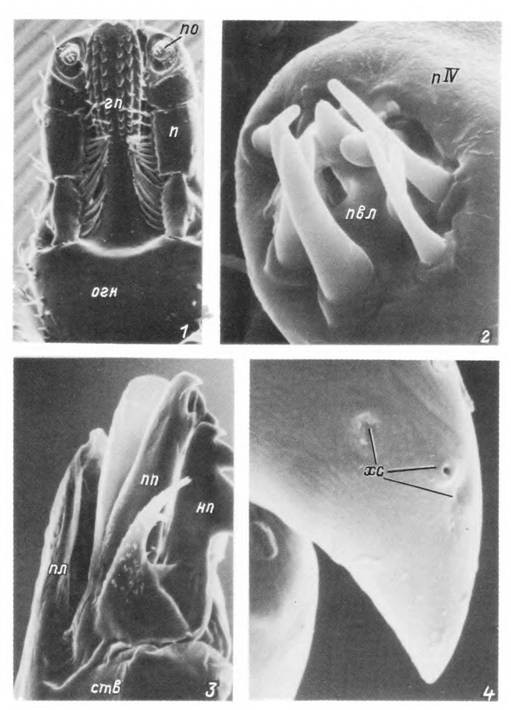
Табл. XXIX. Органы чувств самки клеща Hyalomma osiaticum.
1 — гнатосома с вентральной стороны; 2 — пальпальный орган: 3 — хелицера; 4 — хелнцеральные сенсиллы, гп — гипос- том; огн — основание гнатосомы; п — пальпа; пІѴ — дистальный членик пальпы с пальпальным органом (я«); пел — пористые волоски пальпального органа: пп — подвижный палец хелнцеры; пл — плащ хелнцеры; нп — неподвижный палец хелицеры; ств — ствол хелнцеры; хс — отверстия хелнцеральных сенсилл.
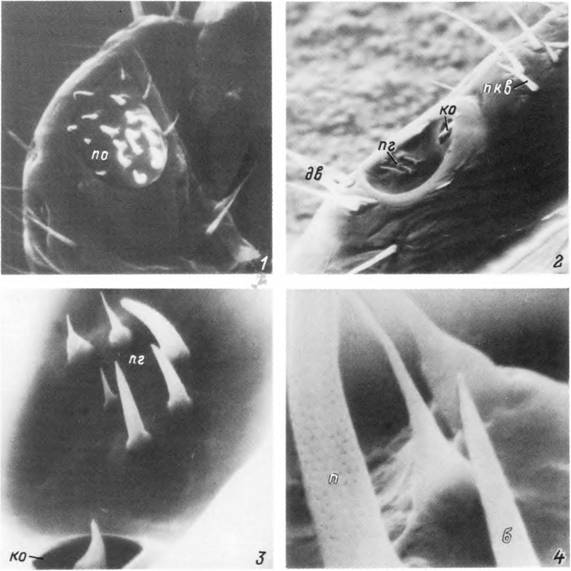
Табл. XXX. Органы чувств самки Ixodes persulcatus.
1 — внешний вид рецепторного пальпального органа; 2 — внешний вид органа Галлера; 3 — внешний вид передней группы сенсилл и капсулярного отверстия органа Галлера; 4 — рецепторные волоски передней группы при большом увеличении. 6 — бороздка; дв — дистальные волоски: ко— капсулярное отверстие; п — пора; пг — передняя группа сенсилл; пкв — посткапсулярные волоски; по — пальпальный орган.
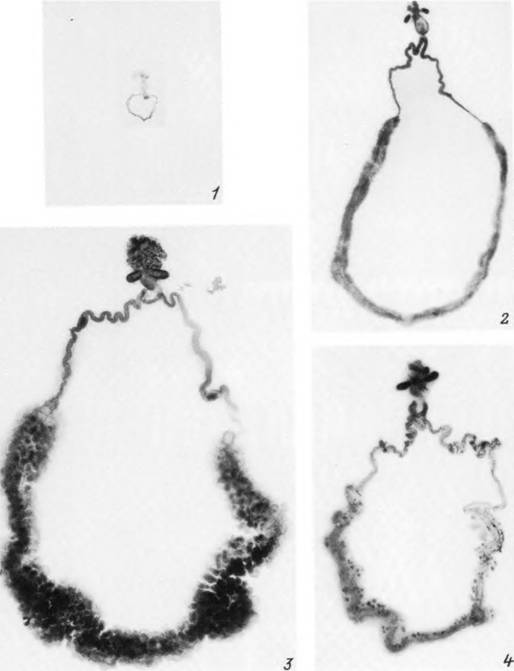
Табл. XXXI. Половая система самки Ixodes persulcatus.
голодная самка; 2 — напитавшаяся перед отпадением; 3 — перед началом яйцекладки; 4 — после окончания яйцекладки.
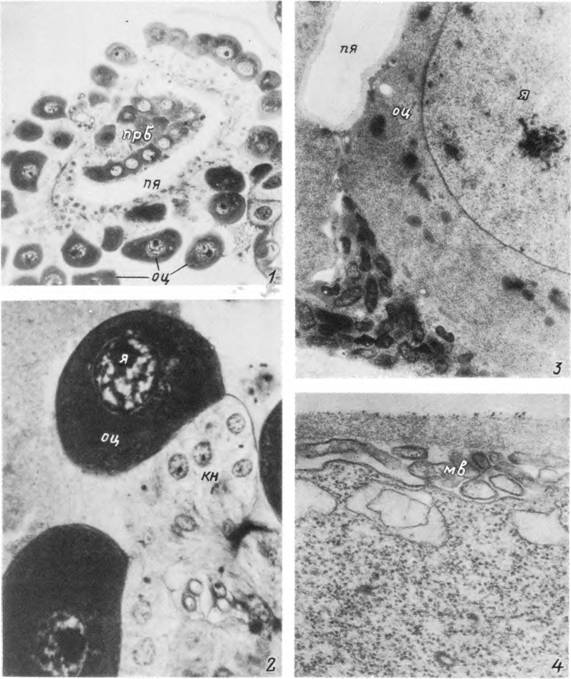
Табл. XXXII. Развитие ооцитов у питающейся самки Ixodes persulcatus.
t — поперечный срез через яичник на 3-й сутки питания; 2 — отдельный растущий ооцит с ножкой на той же стадии; 3 — участок стенки яичника в начале питания; 4 — формирование наружной оболочки ооцита («хориона»), кн — клетки ножки; мв — мнкроворсинки; оц — ооцит; прб — продольная борозда; пя — полость яичника; я — ядро.
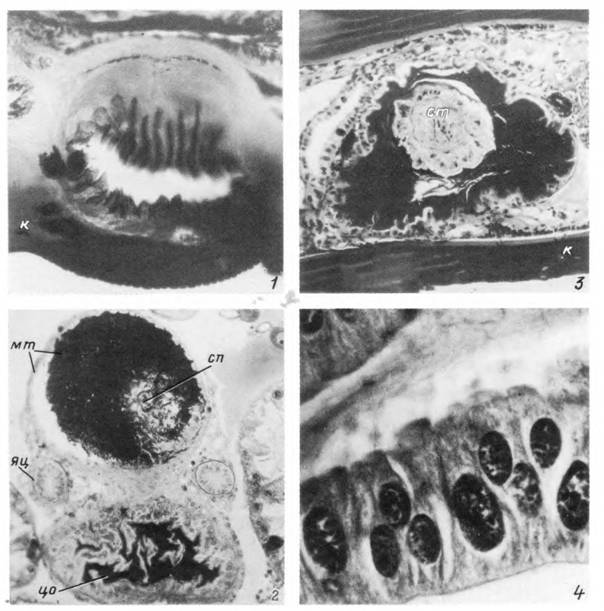
Табл. XXXIII. Выводные пути половой системы самки Ixodes persulccitus.
1 — вестибулярный и 2 — цервикальный отделы влагалища; 3 — место соединения вестибулярного и цервикального отделов; 4 — участок стенки трубчатой придаточной железы питающейся особи, к — кутикула стенки тела; мт — матка; сп — спермин; cm — соединительная трубка; цо — цервикальный отдел; яц — яйцевод.
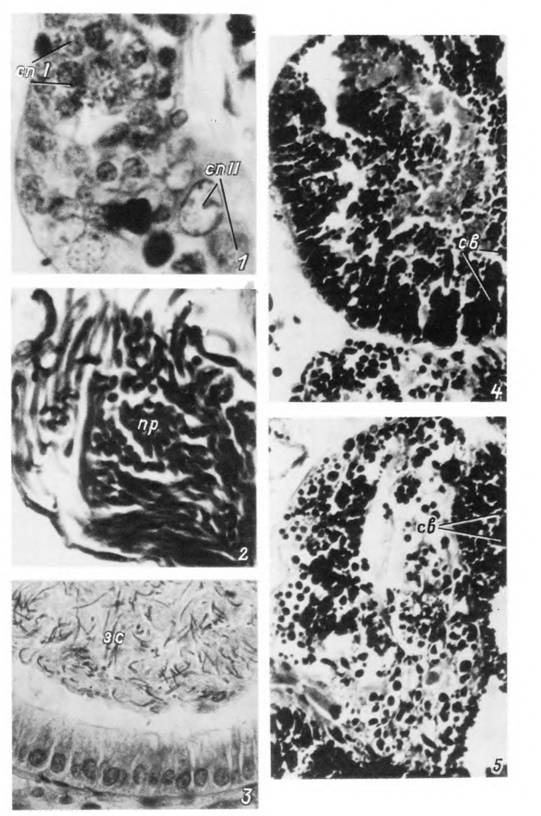
Табл. XXXIV. Мужская половая система и сперматогенез Ixodes persulcatus.
1 — участок семенника с семенными фолликулами; 2 — участок семяпровода; 3 — цервикальный отдел влагалища самки со зрелыми спермиями; 4, 5 — срезы через лопасти придаточной железы с секреторными включениями, зс — зрелые спермин; пр — проспермнн; св — секреторные включения; спі, cnll — сперматоциты I и II порядка.
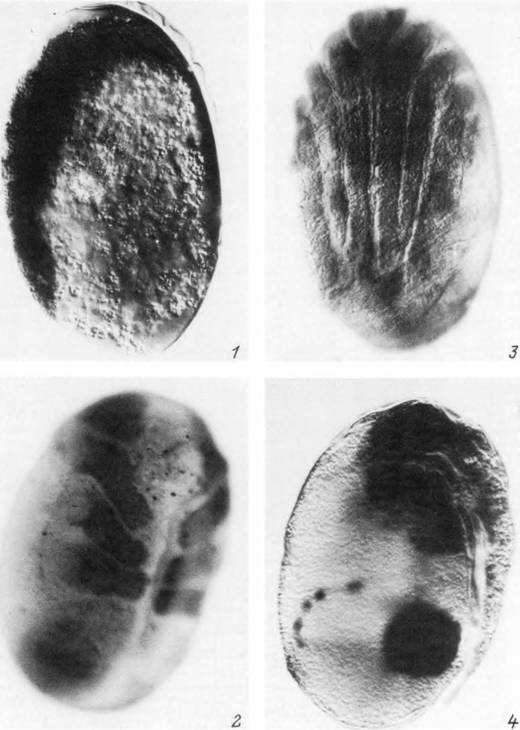
Табл. XXXV. Эмбриональное развитие Ixodes persulcatus.
I — стадия зародышевой полоски; 2, 3 — стадии закладки и дифференцировки конечностей; 4 — стадия формирования мальпигиевых сосудов, ректального пузыря и средней кишки.
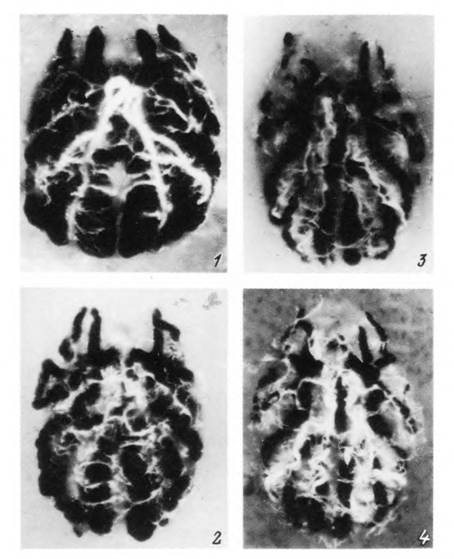
Табл. XXXVI. Голодные самки Dermacentor reticulaius разных физиологических возрастов, вскрытые со спинной стороны. Видна разная степень заполненности дивертикулов средней кишки (черные) и мальпиги- * евых сосудов (белые) (по: Разумова, 1977).
I — особь I физиологического возраста, 2 — II возраста, 3 — III возраста, 4 — IV возраста.
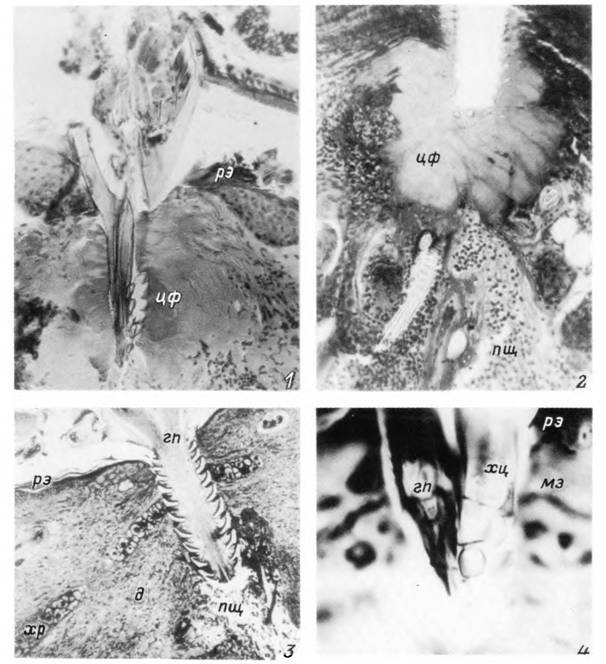
Табл. XXXVII. Погруженные в кожу млекопитающих ротовые органы иксодовых клещей.
1 — хоботок нимфы Hyalomma asiaticum в коже белой мыши; 2 — то же, после удаления хоботка клеша; 3 — хоботок самки Ixodes ricinus в коже кролика; 4 — гипостом и хелицера нимфы Ixodes ricinus в коже рыжей полевки, гп — гипостом; пщ — пищевая полость; мэ — мальпигиев слой эпидермиса; д — дерма; рэ — роговой слой эпидермиса кожи; цф — цементный
футляр; хр — хрящ; хц — ствол и палец хелицеры.
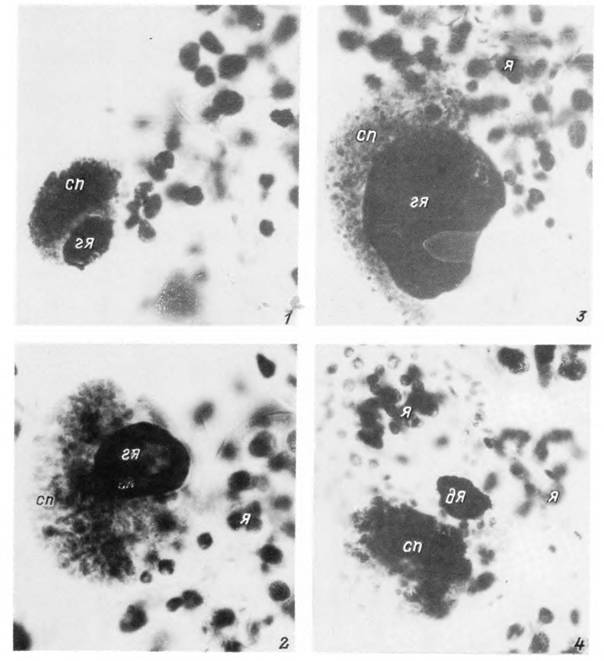
Табл. XXXVIII. Тейлерии Theileria annulata в клетках альвеол III типа слюнной железы самки Нуаіотта anatolicum на 3—4-е сутки от начала питания.
1 — начальная стадия роста споронта; 2 — стадия многоядерных цитомеров; 3, 4 — начало (5) и завершение (4) дифференциации спорозоитов, сп — споронт; гя — гипертрофированное ядро инвазированной клетки; я — нормальные ядра секреторных клеток; дя — дегенерирующее ядро гипертрофированной клетки на стадии освобождения спорозоитов.
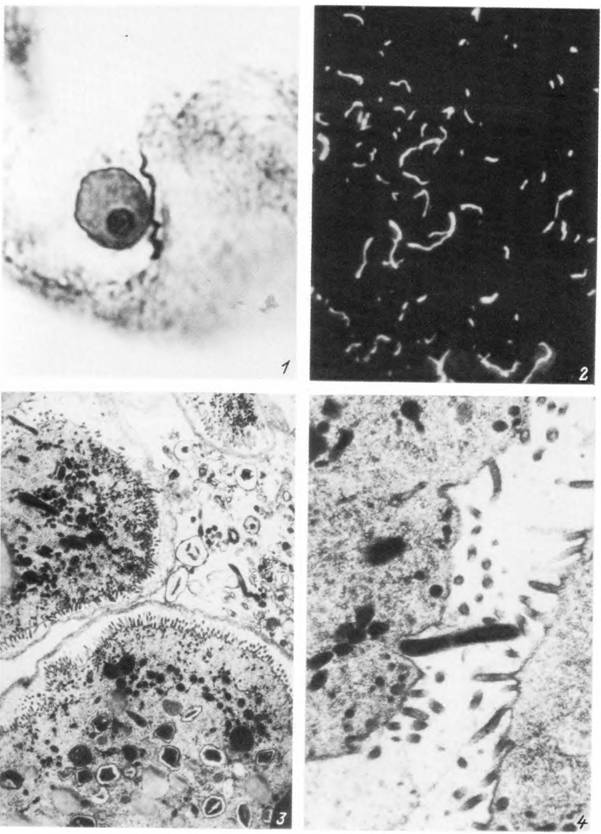
Табл. XXXIX. Borrelia burgdorferi s. lato в организме самки Ixodes persulcatus.
1 — одиночная боррелия в секреторной клетке слюнной железы питающейся особи (серебрение гистологических срезов по Левадитти); 2 — боррелии в давленом препарате участка дивертикула средней кишки голодной особи (темное поле); 3 — электронограм.ма участка дивертикула средней кишки питающейся особи (видны боррелии в полости кишки и внутри кишечных клеток); 4 — электронограмма одиночной боррелии, внедряющемся в кишечную клетку.
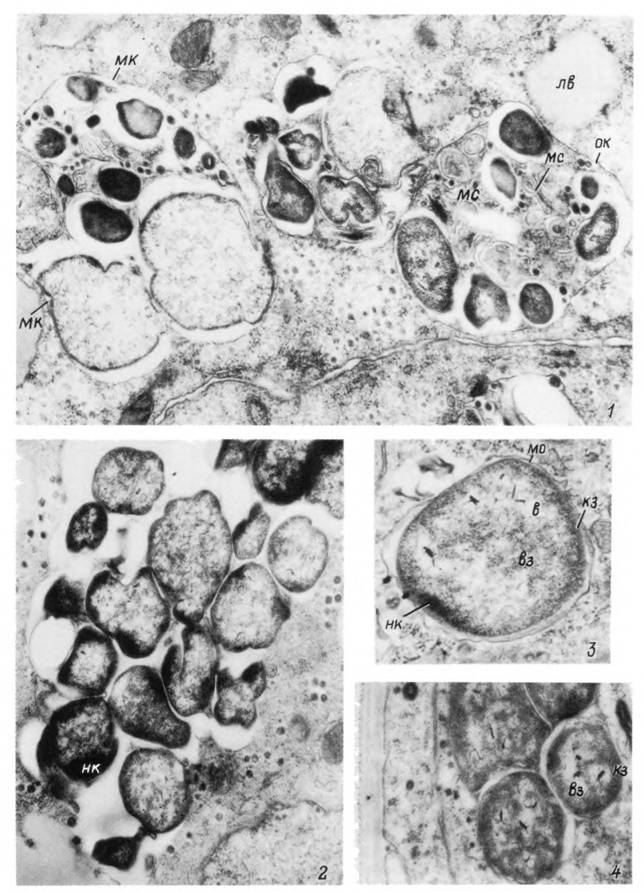
Табл. XL. Риккетсии рода Wolbachia в организме самки клеща Hyalomma asiaticurn.
1 — участок стенки мальпигиева сосуда с колониями риккетсии; 2—4 — риккетсии в развивающихся ооцитах на 2—4-е сутки от начала питания, а — риккетсия; вз— внутренняя цитоплазматическая зона; да— дегенерирующие риккетсии; кі — кортикальная цитоплазматическая зона; лв — лизосомные включения; мк — микроколонии риккетсий; мо — мембрановидная оболочка колоний: мс — мембрановпдная структура риккетсии: нк — нуклеоидные тела; ок — общая оболочка
колонии риккетсий.
Еще по теме Синхронизация жизненных циклов с сезонными климатическими ритмами:
- Описания жизненных циклов
- СЕЗОННОСТЬ И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНЕННЫХ циклов
- ПЕРИОДИЗАЦИЯ ЖИЗНЕННЫХ ЦИКЛОВ
- Происхождение синхронизации медленных колебаний.
- 38. Сезонная спячка как реакция адаптации к неблагоприятным условиям существования.
- Умови оплати праці сезонних робітників протичумних закладів
- Сезонность вспышек холеры
- Климатическое лечение
- Экономический цикл и его фазы. Продолжительность циклов.
- Значение природно-климатических и санитарно-гигиенических условий при холере
- Природно-климатические особенности республики
- ВРЕМЕННОЕ УКОРЕНЕНИЕ ХОЛЕРЫ ЭЛЬ-ТОР И СОХРАНЕНИЕ ЕЕ ВОЗБУДИТЕЛЯ В МЕЖЭПИДЕМИЧЕСКИЙ СЕЗОН
- 4.2 Оценка влияния климатических факторов на клиническое течение бронхиальной астмы
- Природно-климатические и демографические особенности Кыргызстана