ИНДИВИДУАЛЬНАЯ ДОЛГОВЕЧНОСТЬ И ФИЗИОЛОГИЧЕСКИЙ ВОЗРАСТ
Важной эпидемиологической особенностью иксодовых и аргасовых клещей служит их исключительная индивидуальная долговечность по сравнению с кровососущими двукрылыми. Е. Н. Павловским и А.
Н. Скрынник (1960) приводятся данные о выживании без пищи отдельных особей клещей Ornithodoros papillipes в течение И лет, Alveonasus lahorensis — 10 лет, а более мелких Ornithodoros nereensis, О. alactagalis и О. tartakovs- куі — 1.5—4 лет. В наибольшей степени эта способность развита у нимф старших возрастов крупных видов и в наименьшей у личинок. Последние выживают без пищи от нескольких недель (О. tartakovskyi) до нескольких месяцев {Alveonasus lahorensis, Argas persicus, Ornithodoros papillipes).Многие виды иксодовых клещей также отличаются значительной долговечностью в голодном состоянии. Например, в условиях лабораторного содержания взрослые Ixodes ricinus выживали до 27 месяцев (Nuttall а. Warburton, 1911), Hyalomma dromedarii — свыше 420 дней (Поспелова-
Штром, 1955), а Ixodes apronophorus — 454 дня (Голов, 1933). В природных условиях сроки выживания голодных клещей также исключительно велики. И. В. Разумова (1966а) установила, что из меченых имаго Dermacen- tor pictus, выпущенных весной в нормальных для этого вида природных биотопах, до осени того же сезона доживало 49 % клещей, а до весны следующего года — 37%. Таким образом, учитывая зимовку неактивизировав- пшхся клещей, перелинявших из нимф в конце лета и осенью, голодные имаго этого вида дважды зимуют и могут быть активны в течение двух сезонов. Клещи D. marginatus могут выживать до 2 лет (Покровская, 1953), a Ixodes texanus — несколько лет (Gregson, 1956). Выживание голодных клещей зависит как от их физиологических особенностей, так и от факторов внешней среды и в первую очередь температуры и влажности. Как правило, температура, близкая к 0°, и высокая относительная влажность благоприятствуют выживанию.
Так, в наших опытах голодные самки /. persulcatus выживали приО—3°и относительной влажности 98% — 1.5— 2 года, при 25° и той же влажности — 3—4 месяца, а при 25° и 80% относительной влажности — меньше месяца. Сходная картина наблюдается и в природных условиях, где голодные клещи этого вида, не сумевшие найти хозяина, погибают в основном ко второй половине лета.Непостоянство сроков развития даже в потомстве- одной самки или способность голодных клещей переживать в природе два и более сезонов активности обусловливают разновозрастность активных клещей. Между тем знание возрастного состава природных популяций эпидемиологически важных видов иксодид имеет большое практическое значение. По нему можно судить о фактической продолжительности жизненного цикла в данной местности и сроках активности клещей, а также учитывать эффективность истребительных мероприятий. К сожалению, объективные методики определения возрастного состава голодных клещей до последнего времени отсутствовали. Сведения об индивидуальной долговечности клещей в природе основывались в основном на косвенных данных из работ по определению продолжительности циклов развития или сроков выживания голодных клещей в лабораторных условиях. Эти сведения, однако, не могли быть достаточно точными, так как известно, что расходование пищевых резервов голодных клещей идет далеко не равномерно. Так, у Ixodes ricinus (Lees a. Milne, 1951) при пассивном пребывании в растительной подстилке пищевых запасов хватает на несколько месяцев, а во время активного подстерегания хозяев они расходуются за несколько дней или недель.
Методика определения возраста у кровососущих членистоногих достаточно подробно разработана в группе двукрылых главным образом благодаря работам В. Н. Беклемишева и его сотрудников. У самок кровососущих двукрылых различают календарный и физиологический возрасты. Первый характеризует продолжительность жизни насекомого в днях, а второй — число проделанных им яйцекладок. Зная среднюю продолжительность одного гонотрофического цикла, по физиологическому возрасту самок легко судить и об их календарном возрасте.
В качестве критериев для определения физиологического возраста используют изменения в половой системе, связанные с повторными яйцекладками (расширения на яйцевых трубочках, состояние их трахейной системы, количество желтых тел и др.; Детинова, 1962).С совершенно иной ситуацией мы сталкиваемся у иксодовых клещей. Анатомическое строение и физиологические особенности иксодид, относящихся к классу паукообразных (подтип хелицеровых), во многом принципиально отличаются от принадлежащих к другому подтипу насекомых. Весьма существенно также, что иксодовые клещи проходят лишь один гонотрофический цикл и затем погибают. Эти две особенности не позво-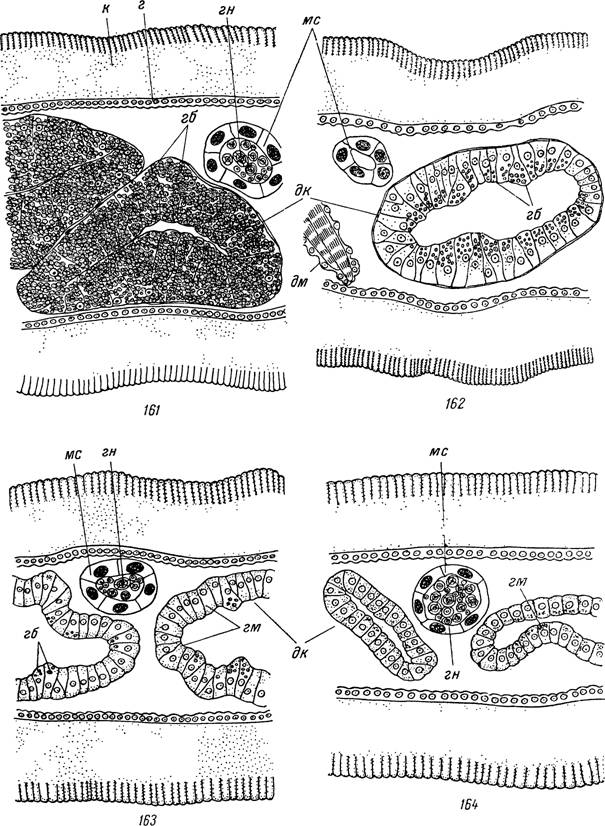
Рис. 161—164. Участки поперечных срезов через голодных самок Ixodes ricinus на 1-й (161), 2-й (162), 3-й (163) и 4-й (164) степенях истощения (схематизировано).
к — кутикула; г — гиподерма; дк — дивертикулы средней кишки; мс — мальпигиевы сосуды; дм — дорсо-вентральные мышцы; гб — пищевые включения из гемоглобина; гм — гематин; гп — гуанин.ляют применить к ним критерии физиологического возраста кровососущих двукрылых.
Изучение гистологических и гистохимических изменений в кишечнике и мальпигиевых сосудах голодных клещей позволило нам использовать их для определения возраста этой группы кровососов. Они характеризуют физиологический возраст клещей, который в отличие от кровососущих двукрылых отражает не число пройденных гонотрофических циклов, а состояние запасных питательных веществ в организме. Зная скорость их расходования в естественных условиях, можно довольно точно установить время, прошедшее после линьки (Балашов, 1961а, 1962а, 19626).
В качестве критериев для определения физиологического возраста клещей сем. Ixodidae мы предложили использовать следующие признаки: степень заполнения отростков кишечника, количество включений гемоглобина и гематина в пищеварительных клетках, степень загрузки мальпигиевых сосудов. Из этих трех показателей только для определения содержания гемоглобина и гематина в клетках требуется применение гистолоіи- ческих методов.
При этом, однако, нет необходимости проводить специальные гистохимические реакции. Кристаллы гематина обладают характерной собственной буро-черной окраской, а включения гемоглобина хорошо окрашиваются железным гематоксилином, эозином и азокармином, широко используемыми в гистологической технике. Для фиксации клещей пригодно большинство фиксаторов, удовлетворительно сохраняющих анатомические и цитологические структуры (смеси Буэна, Карнуа, Ценкера, Суза). Как дополнительные признаки может быть также использовано содержание гликогена и жиров в клетках средней кишки, выявляемых гистохимическими методиками. При определении возраста клещей необходимо одновременно учитывать все три основных признака, так как даже исключение одного из них может привести к ошибочным выводам.По состоянию пищевых резервов в организме голодных клещей мы различаем четыре степени истощенности.
1- я степень. Недавно перелинявшие особи, находящиеся на стадии послелиночного доразвития. Отростки кишечника слегка растянуты, и при разрыве из них вытекает буро-черная кашеобразная масса. Полость кишечника занята дистальными концами гипертрофированных пищеварительных клеток. Плазма последних полностью заполнена включениями гемоглобина, а между ними лежат мелкие кристаллики гематина. Мальпигиевы сосуды беловатой окраски и содержат в просвете много средних и крупных сферитов гуанина (рис. 161).
2- я степень. Особи недавно закончили послелиночное доразвитие. Отростки кишечника не имеют перетяжек. Внутри них — хорошо развитый просвет. Размеры пищеварительных клеток значительно уменьшаются. Внутри последних много включений гемоглобина, располагающихся преимущественно в их дистальных концах и обычно не соприкасающихся между собой. Кристаллов гематина мало и они лежат диффузно. Мальпигиевы сосуды прозрачные. В их просвете отсутствуют средние и крупные сфериты гуанина (рис. 162).
3- я степень. Клещи находились продолжительное время в состоянии активности. На отростках кишечника появляются многочисленные перетяжки, а диаметр их заметно уменьшается.
Внутри них обширный просвет. Пищеварительные клетки свободны от включений гемоглобина или содержат их в очень небольшом количестве. Число кристаллов гематина в клетках значительно увеличивается, но они по-прежнему лежат диффузно. Мальпигиевы сосуды часто имеют четковидную внешность. Их беловатые участки заполнены средними и крупными конкрециями гуа- /нина (рис. 163).4- я степень. Длительно голодавшие, сильно истощенные особи. Отростки кишечника крайне истонченные с многочисленными глубокими перетяжками. Стенки их местами спадаются и просвет исчезает. В пищеварительных клетках никогда не встречаются включения гемоглобина, а кристаллики гематина местами образуют значительные скопления. Мальпигиевы сосуды сильно растянуты заполняющими их сферитами гуанина и имеют на всем протяжении молочно-белую окраску (рис. 164).
Приведенная шкала оценки составлена на основе изучения половозрелых особей и нимф Hyalomma asiaticum и Ixodes ricinus. В основных чертах она подтвердилась и для остальных изученных нами видов иксодо- вых клещей. Видовые различия проявляются преимущественно в различной скорости расходования запасных питательных веществ, их внутриклеточной локализации, форме и размерах пищеварительных клеток и других особенностях. В связи с этим мы считаем возможным рекомендовать использованные нами признаки при изучении возрастного состава природных популяций иксодовых клещей. Запасные питательные вещества в организме личинок представлены эмбриональным желтком и его производными, в связи с чем они занимают особое положение и требуют специального дополнительного изучения.
Практическую применимость разработанной шкалы подтверждают наши исследования физиологического возраста голодных самок Ixodes ricinus и I. persulcatus в Ленинградской обл. Клещи первого вида собирались в Лужском районе с мая по сентябрь в 1958—1960 гг., а второго — в Киришском районе в мае—августе. Всего были собраны и подвергнуты гистологическому изучению 781 самка I. ricinus и 330 — I. persulcatus.
Полученные результаты сведены в табл. 11. Среди пойманных активныхТаблица 11
Физиологический возраст голодных самок Ixodes ricinus и I, persulcatus* собранных в Ленинградской обл.
| Месяц сбора | I. ricinus | I. persulcatus | ||||||
| процент клещей 2—4-й степеней истощенности | всего собрано | процент клещей 2—4-й степеней истощенности | всего собрано | |||||
| 2-я | 3-я | І 4-я | 2-я | 3-я | 4-я | |||
| Май | 60 | 34 | 6 | 283 | 69 | 31 | 0 | 165 |
| Июнь | 24 | 62 | 14 | 159 | 27 | 70 | 3 | 119 |
| Июль | 3 | 53 | 44 | 81 | 0 | 41 | 59 | 46 |
| Август | 33 | 32 | 35 | 146 | 0 | 0 | 100 | 5 |
| Сентябрь | 45 | 33 | 22 | 112 | 0 | 0 | 0 | 0 |
самок отсутствовали особи 1-й степени истощенности, что подтверждают данные о пассивности и неспособности к нападению на хозяев клещей, не закончивших стадии послелиночного доразвития.
У /. ricinus в Ленинградской обл. большинством исследователей отмечается два периода максимальной численности клещей на хозяевах: весенний в мае—июне и осенний в конце августа—сентябре, хотя они встречаются в течение всего теплого сезона (Алфеев, 1948; Померанцев, 1935).
Как видно из наших наблюдений, основную часть майских самок составляли особи 2-й (60%) и 3-й (34%) степеней истощенности. Они несомненно происходят от особей, перелинявших со второй половины лета прошлого года. Особи 2-й степени истощенности, вероятно, впервые активизировались после зимовки, так как подобное состояние, за исключе- ниєм периода зимнею оцепенения, обычно продолжается не более месяца. Из самок 3-й степени истощенности часть, по-видимому, была активна уже в прошлом сезоне и они просто возобновили активность, выйдя из зимнего оцепенения весной следующего года. Наконец, самки 4-й степени истощенности, вероятно, перелиняли из нимф, активизировались с начала второй половины лета и растратили к зимовке большую часть запасных питательных веществ.
В июне процент недавно активизировавшихся самок заметно уменьшается (24%). Последнее связано с уменьшением запаса взрослых клещей, перелинявших в прошлом сезоне. Основную массу в этот период составляют очень агрессивные самки 3-й степени истощенности (62%). Одновременно наблюдается увеличение числа сильно истощенных особей.
В июле наряду с общим снижением численности клещей почти полностью исчезают недавно активизировавшиеся самки и половину пойманных клещей составляют особи 4-й степени истощенности.
В конце августа заметно увеличивается количество активных голодных клещей, а среди них особей 2-й степени истощенности (33%). В сентябре последние составляют уже почти половину от общего числа пойманных самок (45%). Они несомненно перелиняли из нимф в конце июля— начале августа и закончили послелиночное доразвитие при высоких летних температурах менее чем за месяц. Одновременно в августе—сентябре наблюдается уменьшение числа истощенных особей (до 22%). Однако отдельные клещи 4-й степени истощенности попадаются даже в конце сентября, так что вполне возможен их уход на зимовку. Происходят ли они от нимф, перелинявших в прошлом году, или же перелиняли во второй половине лета и после активизации растратили большую часть пищевых резервов — решить трудно, хотя вероятнее последнее. Несомненно лишь, что очень небольшое число подобных особей способно перезимовать и возобновить активность в следующем сезоне.
Приведенные наблюдения вскрывают причины летнего снижения и осеннего подъема численности голодных клещей. Большинство авторов (Алфеев, 1948; Сердюкова, 1952; Хейсин, 19556) считает, что линька нимф происходит в основном во второй половине* лета и осенью. Значительное число клещей 2-й степени истощенности в конце лета и осенью связано с завершением послелиночного доразвития и активизацией части перелинявших особей в том же сезоне. За их счет происходит осенний подъем численности. На зимовку уходят как активные, так и не завершившие послелиночного доразвития клещи. Основная их масса становится активной сразу после зимовки. Однако в связи с тем, что часть клещей уходит на зимовку, не закончив послелиночного доразвития, эта стадия заканчивается уже в следующем весенне-летнем сезоне. Разновременность ее завершения в связи со сроками линьки нимф прошлой осенью, разницей в микроклимате мест развития и другими экологическими факторами обусловливает растянутость активизации клещей до конца июня. К началу июля численность голодных клещей в природе заметно уменьшается как за счет нахождения ими прокормителей, так и от естественной гибели от истощения. В июле обычно встречается максимальное количество особей 4-й степени истощенности. Достигнув этого состояния, клещи довольно быстро погибают. Тем не менее в связи с растянутостью весенне-летней активизации часть клещей, перелинявших в прошлом году, доживает до конца сезона активности следующего года. В результате осенняя популяция половозрелых Ixodes ricinus состоит из особей, принадлежащих к двум генерациям. В северной части ареала (Карельская АССР) летнего снижения численности клещей вообще не наблюдается (Хейсин, Бочкарева и Лаврененко, 1955), а осенняя активизация перелинявших в том же сезоне самцов и самок, вероятно, отсутствует. Напротив, в южной части ареала (Бабенко, 1958) взрослые клещи летом встречаются очень редко и во время осеннего подъема их численности, по-видимому, встречаются только особи, перелинявшие в том же сезоне.
Иную картину мы наблюдаем при анализе активности голодных взрослых /. persulcatus. На всем протяжении ареала они активны весной и в первой половине лета, а затем очень быстро исчезают. Линька нимф, как и у /. ricinus, происходит только во второй половине лета и осенью (Хейсин, Павловская, Малахова и Рыбак, 1955; Бабенко, 1956). Неясными остаются причины исчезновения клещей в середине лета.
Изучение физиологического возраста самок этого вида показывает, что в мае все клещи были 2-й (69%) и 3-й (31%) степеней истощенности. Они перелиняли из нимф во второй половине лета и осенью прошлого года. Часть из них закончила послелиночное доразвитие уже осенью того же года, но активизация произошла только после перезимовки. У некоторых клещей описываемая стадия завершается после зимовки, и активизация подобных особей растягивается на июнь. Расходование запасных питательных веществ у I. persulcatus идет быстрее, чем у /. ricinus. В июле 59% пойманных клещей составляли сильно истощенные особи, а к августу большинство из не сумевших найти хозяина клещей погибало от истощения. Переживание голодными клещами этого вида двух сезонов активности невозможно. Пользуясь предложенной нами методикой, были изучены изменения возрастного состава имаго /. persulcatus в Карельской АССР, Красноярском крае и в Пермской обл., и во всех случаях главной причиной смертности среди голодных клещей в конце сезона активности было израсходование запаса питательных веществ (Хижинский, 1963; Князева, 1964; Лыков, 1966).
Еще по теме ИНДИВИДУАЛЬНАЯ ДОЛГОВЕЧНОСТЬ И ФИЗИОЛОГИЧЕСКИЙ ВОЗРАСТ:
- Продолжительность жизни и физиологический возраст
- 23. Индивидуальный стиль трудовой деятельности (ИСД) и интегральная индивидуальность; устойчивость и изменчивость ИСД; ИСД и эффективность труда.
- 65. Развитие личности в возрасте поздней зрелости, пожилом и старческом возрасте.
- Медицинский контроль за физиологической полноценностью питания военнослужащих
- Физиологические реакции на стресс
- Билет 14. Физиологические основы психотерапии.
- Билет 14. Физиологические основы психотерапии.
- 26. Индивидуальное консультирование.
- Регенерационный остеогенез в организме взрослых (физиологический и репаративный).
- 36. Проведение группового и индивидуального психологического обследования.
- Индивидуальная психология.
- Определение физиологических потребностей ребенка
- Физиологические особенности детской кожи
- Физиологические особенности детской кожи
- Терминология индивидуально приобретенных рефлексов.
- Атрофические процессы в физиологической жизни
- Билет 42. Индивидуальная психотерапия.
- 10. Индивидуальный стиль деятельности.