Общие особенности
Иксодовые клещи принадлежат к экологической группе временных паразитов с длительным питанием (Балашов, 1984). В их жизненном цикле стадии личиночного, нимфального и имагинальноЬэ питания, когда они являются настоящими паразитами, чередуются с разделяющими их непаразитическими стадиями эмбрионального развития, метаморфоза напитавшихся личинок и нимф, яйцекладки самок и существованием голодающих особей до встречи с хозяевами (рис.
37). По числу сменяемых хозяев и месту линьки жизненные циклы клещей принято подразделять на трех-, двух- и однохозяинные.Суммарное время питания всех 3 фаз развития занимает не более 12—15 сут (по 3—6 сут у неполовозрелых особей и 6—12 сут у самок). У треххозяинных видов указанными сроками и ограничивается все их паразитическое существование, тогда как непаразитический период их жизни продолжается от нескольких месяцев до нескольких лет (табл. 3). У некоторых видов, как например D. reticulatus (Разумова, 1966; Cerny et al., 1982), наблюдается исключительная индивидуальная долговечность голодных имаго (до 2.5 лет), за счет чего доля непаразитического существования значительно увеличивается. Еще длиннее непаразитическая часть жизненного цикла у /. ricinus, /. persulcatus и других видов с многолетними циклами развития, включающими стадии диапаузы. Соответственно при 1-годичных циклах развития на паразитические стадии приходится 6—7 % от общего времени развития 1-го поколения, при 3-годичных — 2—2.5 % и при 5-летних — всего 1.4 %.
У двуххозяинных (табл. 4) и особенно однохозяинных (табл. 5) видов удельный вес времени паразитического существования несколько больший за счет добавления к периодам питания времени метаморфоза на теле хозяина личинок и нимф, а непаразитического существования — соответственно укорачивается. В результате соотношение между периодами паразитического и свободного существования в жизненном цикле существенно выравнивается.
Например, у однохозяинного В. microplus в тропическом климате развитие 1-го поколения занимает 2—3 мес (Nunez et al., 1985), из которых на паразитические стадии приходится около 23 сут или около 25—38 % от общего времени развития.Наибольшее распространение имеет треххозяинный цикл, свойственный всем видам в родах АтЫуотта, Аропотта, Ixodes, Haemaphysalis и большинству в родах Dermacentor, Rhipicephalus, Hyalomma. Примерами могут служить клещи /. persulcatus (рис. 38) или /. ricinus, питающиеся на неполовозрелых фазах на мелких млекопитающих, птицах, добывающих корм на поверхности почвы, или даже на рептилиях. Взрослые клещи этих видов паразитируют на крупных и средних млекопитающих, включая сельскохозяйственных животных. Некоторые виды при этом могут на всех фазах развития питаться на одном и том же виде хозяина, связанного с постоянной норой или гнездом (/. lividus, /. crenulatus, R. pussilus, H. hispanica и др.), или реже на не имеющих постоянного убежища бродячих животных (R. appendiculatus, A. hebraeum, A. variegatum). При подобном
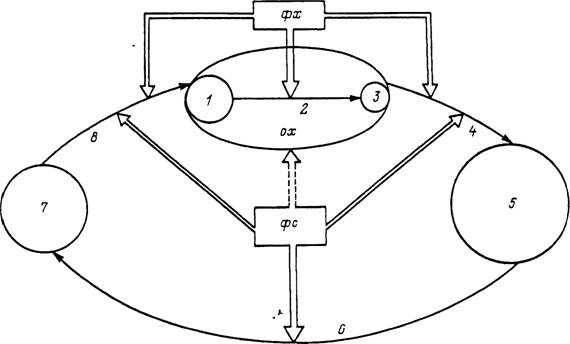
Рис. 37. Влияние факторов внешней среды и организма хозяина на жизненный цикл однохозяинного иксо-
дового клеща.
1 — число прикрепившихся личинок; 2 — питание и линька личинок, питание и линька нимф, питание самок; 3 — число напитавшихся самок; 4 — оогенез, яйцекладка и эмбриональное развитие; 5 — число вылупившихся личинок; б — свободное существование личинок до встречи с хозяином; 7 — число личинок, нападающих на хозяина; 8 — нападение на хозяина; ох — организм хозяина; фх — факторы организма хозяина (степень привлекательности для клеща, способность к самоочищению от клещей, уровень противоклещевой резистентности, пищевая ценность крови и др.); фс — факторы внешней среды, воздействующие на клеща; стрелки указывают направление действия факторов (сплошная линия — прямое и прерывистая линия — косвенное воздействие).
типе развития клещи, независимо от вида хозяина и особенностей местообитания, после завершения питания отпадают во внешнюю среду, линяют там на следующую фазу и подстерегают нового хозяина.
При двуххозяинном цикле (рис.
39) личинка, закончив кровососание, остается прикрепленной к хозяину, линяет здесь же на нимфу и клещ отпадает по окончании нимфального питания. Двуххозяинный тип развития чаще всего наблюдается у паразитов копытных животных (R. bursa, Н. detritum, Н. anatolicum), причем как неполовозрелые фазы, так и имаго паразитируют на одном или близких видах. Из этого правила наблюдаются и исключения, как например в случае питания личинок и нимф двуххозяинного Н. marginatum на птицах и грызунах, а имаго — на копытных. Наиболее приспособлены к паразитированию на бродячих животных клещи с однохозяинным типом развития (рис. 40), когда после прикрепления личинок на теле хозяина завершаются 3 питания, 2 линьки и отпадают только напитавшиеся самки. Подобный тип развития кроме всех видов Boophilus обнаружен у Н. scupense, A. nitens, D. albipictus.Следует отметить, что в роде Нуаіотта границы между трех-, двух- и однохозяинным развитием могут быть довольно условными. Значительное влияние на тип развития может оказать видовая принадлежность хозяина. В наших опытах при кормлении на кроликах большая часть клещей Н. anatolicum развивалась по треххозяинному типу, тогда как в природе при питании на скоте для них характерен двуххозяинный цикл. При питании на ежах треххозяинного Н. asiaticum часть особей развивалась по двуххозяинному циклу вследствие задержки напитавшихся личинок и линьки их на теле хозяина.
Значительное влияние на изменение цикла развития оказывают видовая принадлежность хозяев и окружающая температура. При питании личинок и нимф на крупном рогатом скоте Н. anatolicum имеет двуххозяинный цикл, а при питании на песчанках — треххозяинный. Сходным образом при питании на кроликах поПродолжительность развития (в сут) треххозяинных видов иксодид в лабораторных условиях
| Личинка | Нимфа | Самка | |||||||||
| Вид клеша | Хозяин | Температура (°С) | Питание | Линька | Питание | Линька | Питание | Подготовка к яйцекладке | Яйце кладка | Вылуп- ление личинок | Литературный источник |
| Лmblyomma атегісапит | Кролик | 21—26 | 3—7 | 9—27 | 3—6 | 14—19 | 10—16 | 7—10 | _ | 24—31 | Loomis, 1961 |
| А. hebraeum | Скот, кролик | 26 | 6—8 | 14—25 | 6—9 | 20—29 | 6—12 | 10—14 | 30—38 | 54—61 | Norval, 1974 |
| A. maculatum | То же | — | 2—10 | 7—121 | 4—11 | 17—71 | 5—18 | 39 | 13—75 | 21—142 | Diamant, Strickland, 1965 |
| А. та г то гейт | Черепаха | 26 | 30 | 2—3 | 51 | 21—28 | 60 | 12—15 | 30 | 37 | Norval, 1975b |
| A. variegatum | Скот, кролик | 27 | 6 | 24 | 6 | 18 | 12 | 12 | 18 | 34 | Borght-Elbl, 1977 |
| Dermacentor andersoni | Кролик | 21—26 | 3--6 | 16—27 | 6—8 | 18—34 | 7—12 | 8—22 | 24—28 | Loomis, 1961 | |
| D. marginatus | Кролик, мышь | 22 | 3—8 | 9—11 | 5—8 | 20—23 | 7—13 | 8—10 | 19—37 | Honzakova, 1971 | |
| D. nuttalli | Кролик, скот | — | 2—3 | 8—12 | 7—8 | 14—20 | 8—10 | 3—25 | ЗІ^Н | 15—36 | Teng Kuo-fan, 1962 |
| D. reticulatus | То же | 22 | 2—8 | 7—8 | 5—8 | 18—31 | 7—9 | 1—7 | — | 17—27 | Honzakova, 1971 |
| D. sinicus | » | — | 3—4 | 7—10 | 5—7 | 18—19 | 8—10 | 9—12 | 22—29 | 25—27 | Teng Kuo-fan, 1981 |
| Haemaphysalis concinna | Мышь, кролик | 22 | 3—8 | 9—11 | 5—8 | 20—23 | 8—12 | 10—15 | — | 53—67 | Honzakova, 1971 |
| H. longicornis | Кролик | 25 | 3—5 | 10—18 | 3-^6 | 12—20 | 4—9 | 4—7 | 16—19 | 20—23 | Fujisaki et al., 1976 |
| H. punctata | Мышь, кролик | 22 | 5—7 | 25—28 | 3—5 | 25—30 | 9—14 | 7—11 | — | 27—54 | Honzakova, 1971 |
| H. sulcata | Песчанки, кролик | 28 | 2—8 | 20 | 9 | 16—21 | 13—18 | 15 | — | 48—54 | Hadani et al., 1969 |
| Hyalomma anatolicum | Морская свинка | 28 | 3 | 7—8 | 4 | 19—29 | 6—8 | 10—11 | 15—19 | 2—8 | Snow, 1969 |
| H. asiaticum | Кролик, мышь | 22 | 4—6 | 11 — 19 | 7—12 | 22—23 | 10—13 | 4—8 | — | 26—39 | Honzakova, 1971 |
| Ixodes hexagonus | Еж, кролик | 22 | 5—18 | 7—31 | 6—14 | 32—44 | 8—13 | 1—50 | — | 17—27 | To же |
| I. laguri | Мышь | 15—17 | 3—16 | 28—180 | 4—7 | 33-42 | 11 | 5—13 | 11 | 28-40 | Honzakova et al., 1980 |
| I. lividus | Ласточка | 27—28 | 3—5 | 10 | 4 | 10—11 | 5—6 | 6—8 | 12—46 | 8—12 | Глащинская-Бабенко, 1956 |
| I. persulcatus | Кролик | 25 | 3—6 | 20—25 | 4—5 | 35—76 | 5—8 | 6—8 | 16—31 | 29—33 | Fujisaki et al., 1976 |
| /. ricinus | Мышь, кролик | 22 | 2—5 | 31—224 | 3—6 | 35—55 | 6—9 | 4—22 | — | 36—64 | Honzakova, 1971 |
| I. trianguliceps | Рыжая полевка | 23—25 | 4—7 | 25—30 | 4—7 | 25—30 | 7—8 | 13—17 | 10—12 | 30—42 | Бобровских, 1965 |
| Rhipicephalus sanguineus | Кролик | 27 | 3—5 | 7—14 | 3—5 | 14—18 | 6—14 | 2—7 | — | 20—32 | Pegram, 1984 |
| R. schulzei | Мышь, морская свинка | 25—28 | 3—5 | 12—33 | 3—6 | 14—33 | 5—8 | 6—15 | — | 33-42 | Кондрашкина, 1951 |
| R. turanicus | Кролик | 27 | 3-4 | 8—9 | 3—5 | 14—19 | 6—12 | 2—5 | — | 20—25 | Pegram, 1984 |
| Вид | Hyalomma del г it ит | Н. marginatum | Rhipicephalus hursa | R. evertsi |
| Литературный источник | Hadani et al., 1969 | Hadani et al., 1969; Knight et al., 1978 | Hadani et al., 1969 | Londt, Bijl, 1977 |
| Регион | Израиль | Израиль, Южная Африка | Израиль | Южная Африка |
| Температура (°С) | 28 | 26—28 | 28 | зо |
| Хозяин | Кролик | Кролик | Кролик | Кролик, крупный рогатый скот |
| Подготовка к яйцекладке | 8—23 | 5—11 | 6—10 | 4—6 |
| Яйцекладка | — | — | — | 14—27 |
| Развитие яиц | ЗО—*0 | 23—38 | 21—25 | 25—27 |
| Паразитирование личи | 16—34 | 16—28 | 15—35 | 13—29 |
| нок и нимф | ||||
| Линька напитавшихся | 20-40 | 18—32 | 14—29 | 13—20 |
| нимф в имаго | ||||
| Питание самок | 12—26 | 8—22 | 6—19 | 6—10 |
треххозяинному циклу развивается до 20 % особей Н. dromedarii (Ouhelli, 1994). Дельпи (Delpy, 1946) отмечает, что при высоких температурах воздуха Н. dromedarii имеет треххозяинный цикл развития, а при более низких — двуххозяинный. В условиях Туркмении (Бердыев, 1980), где этот вид обычен на верблюдах на всех фазах развития, в теплое время года цикл развития треххозяинный, а зимой наблюдается задержка с отпадением напитавшихся нимф. В наших опытах при питании личинок Н. asiaticum на кроликах при температуре 18—20 °С все напитавшиеся особи сразу же отпадали, а при 5—6 °С примерно 5—10 % личинок остались на кролике и перелиняли на нем в нимф.
Большинство исследователей считают исходным треххозяинный цикл развития. Переход к двух- и однохозяинности рассматривается как одна из важнейших адаптаций к паразитированию на бродячих копытных малоподвижных кровососов с подстерегающим типом нападения (Балашов, 1984). Наибольшая смертность у иксодид приходится на долю голодных клещей, не сумевших найти хозяина. С переходом клещей родов Boophilus, Hyalomma, Rhipicephalus к питанию на копытных животных в открытых степных и пустынных ландшафтах возможности их встречи с потенциальными хозяевами значительно ниже, чем у норовых и лесных
Таблица 5
Продолжительность развития (в сут) однохозяинных видов иксодид в природе
| Вид | Anocentor nitens | Boophilus microplus | Dermacentor alhipictus | Hyalomma scupense |
| Литературный источник | Abreu et al., 1986; Diaz, Vega, 1987 | Nunez et al., 1985 | Drew, Samuel, 1986; Addison, McLaughlin, 1988 | Галузо, 1947 |
| Регион | Куба | Аргентина | Канада | Казахстан |
| Хозяин | Крупный рогатый скот | Крупный рогатый скот | Лось | Крупный рогатый скот |
| Подготовка к яйцекладке | 3.1 | 3—6.6 | 36—41 | 15—20 |
| Яйцекладка | 10 | 9—32 | — | 41—66 |
| Развитие яиц | 19.9 | 23—75 | 79—106 | 32—68 |
| Непаразитическое существование личинок | — | 55—133 | Несколько месяцев | Несколько месяцев |
| Паразитическое существование | 16—34 | 20—41 | 175 (мин) | 120—180 |
-ЮС Балашов
97
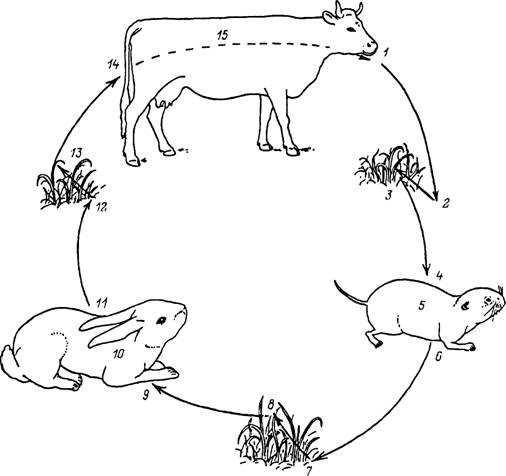
Рнс. 38. Треххозяинный цикл развития клеща Ixodes persulcatus.
I — отпадение напитавшихся самок; 2 — уход самок в растительную подстилку, яйцекладка, гибель самок и развитие личинок; 3 — активизация личинок и подстерегание хозяев на поверхности почвь/ или растительности; 4 — нападение личинок на хозяина; 5 — питание личинок; 6 — отпадение напитавшихся личинок; 7 — уход напитавшихся личинок в растительную подстилку и линька в нимф; 8 — подъем активных нимф на растительность и подстерегание хозяев; 9 — нападение иа хозяина; J0 — питание нимф; II — отпадение напитавшихся нимф; 12 — уход напитавшихся нимф в растительную подстилку и линька в имаго; 13 — подъем напитавшихся имаго иа растительность, спаривание и подстерегание хозяина; 14 — нападение иа хозяина; 15 — спаривание и питание самок.
видов. Приобретение в этих условиях способности к 1 или 2 линькам на теле хозяина увеличивает шансы выживания и успешного завершения развития. Мы также полагаем, что в условиях суровой зимы однохозяинность могла развиться как приспособление к зимовке в условиях теплого микроклимата поверхности тела хозяина. Известны случаи задержки в питании на протяжении всей зимы у отдельных особей D. silvarum и D. sinicus, прикрепившихся к хозяину осенью (Белозеров, 1973; Teng Kuo-fan, 1981), а такЗсе упоминавшаяся задержка напитавшихся нимф Н. dromedarii на теле верблюдов в период зимних холодов. У клещей D. albipictus и Н. scupense уже все развитие от прикрепления личинок до отпадения напитавшихся самок сместилось на холодное время года, и, по-видимому, именно благодаря однохозяинному циклу эти виды продвинулись на север дальше других представителей своих родов.
Глубокие различия в морфофизиологических и поведенческих особенностях иксодид, а также в их требованиях к условиям окружающей среды во время жизни голодных особей, питания, линьки или яйцекладки позволяют рассматривать эти периоды как самостоятельные стадии в жизненном цикле. Эти стадии повторяются на каждой из 3 активных морфологических фаз, а внутри их возможны более дробные подразделения, подробно рассматриваемые в следующих главах.
Иксодовые клещи принадлежат к членистоногим, которые сравнительно легко культивируются в лаборатории. Поэтому для многих видов известны сроки их1 — отпадение напитавшихся самок; 2 — уход клещей в растительную подстилку, яйцекладка, гибель самок и развитие личинок; 3 — активизация личинок, подъем иа растительность и подстерегание хозяев; 4 — нападение на хозяина; 5 — питание личинок, линька в нимф, питание иимф без смеиы хозяина; б — отпадение напитавшихся иимф; 7 — уход иимф в растительную подстилку или почву и линька в имаго; 8 — подъем имаго на растительность и подстерегание хозяев; 9 — нападение на хозяина; 10 — спаривание и питание самцов и самок.
развития в условиях контролируемой температуры, влажности, светового режима при кормлении на лабораторных животных (табл. 3, 4). Однако экстраполяция этих данных на возможные циклы развития клещей в природе во многих случаях некорректна. Более надежными оказались результаты изучения сроков развития
отдельных стадий в их естественных микробиотопах при нахождении клещей в разных типах садков из металлической или капроновой сетки, а также на огороженных участках местности. Суммирование сроков развития напитавшихся особей и продолжительности жизни голодных с определенными поправками позволяет установить реальное время одной генерации. У многих видов этот показатель может значительно варьировать не только в разных частях ареала и в разные годы, но даже в потомстве 1 самки.
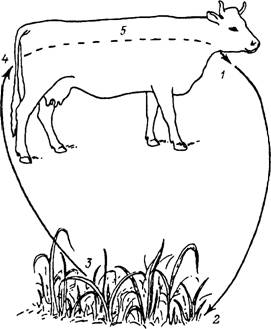
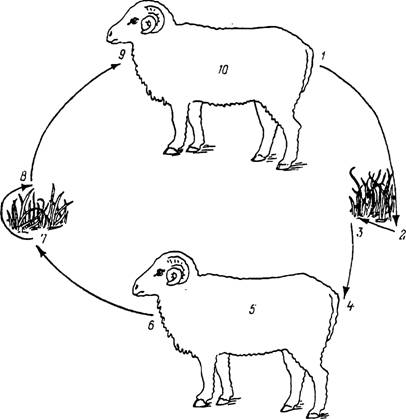
Рис. 39. Двуххозяинный цикл развития клеща Rhipicephalus bursa.
Рис. 40. Однохозяинный цикл развития клеща Boophi- lus annulatus.
1 — отпадение напитавшихся самок; 2 — уход самок в растительную подстилку или почву, яйцекладка, гибель самок и развитие личинок; 3 — подъем личинок на растительность и подстерегание хозяев; 4 — нападение личинок; 5 — питание личинок, линька в нимф, питание нимф, линька в имаго, спаривание и питание самцов и самок без смены хозяев.
Для иксодид характерно исключительное многообразие жизненных циклов по их общей продолжительности, сезонной активности голодных особей, распространению диапаузы и ее значению в переживании неблагоприятных условий. При отсутствии диапаузы, как например у В. microplus или A. nitens, наблюдаются непрерывные (гомодинамные) циклы, так что в течение года происходит смена нескольких налегающих друг на друга поколений. При наличии диапаузы жизненные циклы могут быть поливольтинными, когда за 1 год успевают развиваться 2 поколения, и моновольтинными с единственным поколением за 1 год. 1-годичные циклы широко распространены в тропиках и субтропиках, а у части видов и в умеренном климате. Однако чаще в умеренном поясе встречаются 2- или 3-летние, а в случаях с /. ricinus и /. persulcatus описаны даже 5-летние циклы развития. Жизненные циклы иксодид приспособлены к климатическим условиям их местообитаний, носят ярко выраженный адаптивный характер и их важнейшие особенности рассмотрены в следующем разделе на примере некоторых наиболее полно исследованных видов.
Еще по теме Общие особенности:
- ЗАНЯТИЕ №6 Особенности и общие принципы статистического анализа биомедицинских данных
- Общие результаты особенностей аэродинамических
- Общие особенности организации медицинской помощи:
- 2.1. Особенности и общие принципы статистического анализа биомедицинских данных
- п.8.3. Особенности эвакуационных мероприятий и общие принципы госпитализации больных или лиц с подозрением на COVID-19*
- 15.0. Общие биологические депрессанты (снотворные средства, общие анестетики, летучие инсектициды)
- Общие затраты
- I. Общие положения
- Общие мероприятия
- Общие принципы диагностики и лечения острых отравлений
- Общие положения
- 7.6.1. Общие закономерности межличностных отношений
- 1. Общие правила системы TNM.
- Общие вопросы
- Общие факторы психотерапевтических методов