УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
По мере роста экзоэритроцитного шизонта ядро его делится; затем разросшееся протоплазматическое тело паразита подразделяется на цитомеры, по периферии которых образуются многочисленные пальце-
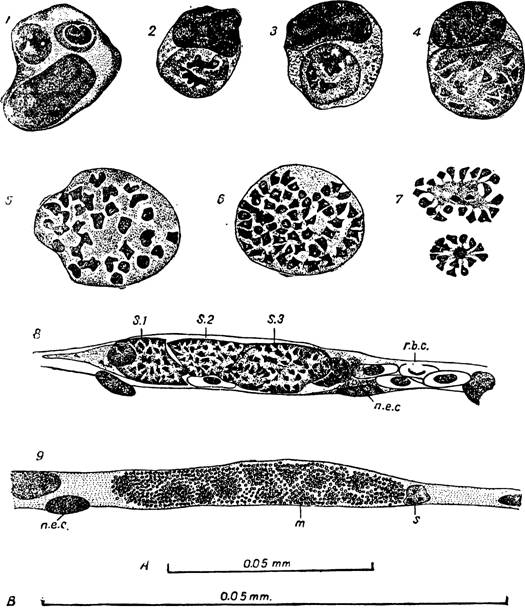
Рис.
88 — Plasmodium gallinaceum. Экзоэритроцитная шизогония./— моноц т из ма*ка легких с двумя молодыми плазмодиями; 2 — моноцит из мазка и чени; в іиизоіге видны две х юмии. оз .іе массы; 3 -4- даль е иие деление хроѵіатиновых млсс в шизанге; 5—6 бо іьлие созревши.* ш ізонты ч мазках легких;
7— груп’Ы мерозоигов из ра «рудіені-ого шизонта; 8— капилл » моіга, плокиро занный Тремя боль НИМИ ШИІО'ІТЗѴІИ (s„ S', Sx\ r.b.c. зритродиг, n.e.c. — я іро э нотелиаль- ной клетки); 9 — капилляр мозга, блокированный множествам мероз итов, окружающих остатки материнской осо >и (тп). Па S. James и P.Tate. .
видные выросты; в них вступают ядра, благодаря чему сформировываются мерозоиты. От клетки хозяина долгое время остается как бы наружная капсула, переполненная множеством мерозоитов. Мерозоиты состоят по внешности как бы из одного хроматина (James и Tate).
Все авторы говорят, что экзоэритроцитная шизогония существует наряду или добавочно к обычно протекающей эритроцитной шизогонии.
Эти новые вопросы требуют дальнейшего изучения, особенно в применении к малярийным паразитахМ человека; необходимо также решить вопрос: является ли экзоэритроцитная шизогония процессом, предшествующим обнаружению шизогонии в эритроцитах, или же эти процессы идут независимо друг от друга, или в какой-либо корреляции один за другим. Этот вопрос нуждается в дальнейшей разработке. Интересны
попытки обнаружения спорозоитов в s.2 тканях человека. Boyd и Kitchen (1939) на срезах биопсированных паховых лимфатических желез, ближайших к месту кормления спорозоитных анофелес на бедре человека, видели спорозоитов, которые лежали или в соединительной ткани трабекул узла или в околоузловой соединительной ткани (рис.
90). Такая участь постигает лишь часть спорозоитов, которые не попали прямо в кровь. Нели, нет каких-либо препятствующих обстоятельств (повышенная сопротивляемость организма хозяина, наличие в крови лекарственных веществ и др.), то через известный период у человека ..появляется первый приступ малярии. Длина такого срока инкубации вариирует для каждого вида малярийного плазмодия. При PL ѵіѵах (терциана) инкубационный период колеблется от 14 до 18 дней (более редкими крайними сроками являются,7— 20 дней); при PI. malariae (квартана)— 18 — 21 день (15—25 дней) и при PI falciparum (тропика) —9 — 12 — 19 дней.
Рис. 89 — Plasmodium gallinaceum.
Внизу — часть среза селезенки с большим шизонтом 5 в эндотелиальной клетке венозного синуса; т — скопление мерозоитов вокруг остатков материнской особи; nhc— дегенерирующее ядро эндотелия; р — малярийный пигмент; spc — клетка селезенки. По S. James и Р. Tate. Вверху — разрез легкого. s1% sit s3 — s$ — большие щизонты в клетках, прилежащих к альвеоле; nhc— ядро клетки, пораженной плазмодием. По S. Л a m е s и Р. Т a t е.
За инкубацией следует продромальный период, длящийся неопределенное время от нескольких часов до 3 дней с нетипичными болезненными проявлениями в форме чувства общей разбитости, головной боли и др.
За ним следуют периодические приступы малярии.
Затем наступают лихорадочные приступы. Длительность приступа лихорадки, включая и ее падение, колеблется от 8 до 16 часов.
При тропической малярии приступ лихорадки удлйнен, и температура падает не сразу и не равномерно. Поэтому настоящих безлихорадочных периодов может вовсе не быть.
Однако накопилось большое количество случаев, которые можно объяснить принятием длинного срока инкубации до нескольких месяцев и даже до одного года. О таком длительном латентном периоде говорят при случаях первичной малярии, падающих на раннюю весну или на зиму, когда исключается ближайшая возможность естественного заражения через анофелес. Подобный вывод можно сделать, только исключив раннее нетипичное (или забытое больным) заболевание малярией; в противном случае весеннее заболевание является уже не результатом первичной инвазии, а рецидивом.
Необходимо также иметь в виду возможность зимования спорозоитов малярийного плазмодия в организме анофелес, благодаря чему первичное заражение может происходить от таких комаров при благоприятной обстановке и зимой и ранней весной.
Рис. 90 — Спорозоиты малярийного плазмодия в соединительной ткани около лимфатического узла человека. По М. Boyd и S. Kitchen.
Длительную инкубацию называют иначе первичным латентным периодом, он сменяется затишьем или вторичным латентным периодом.
Размножение паразитов в, крови идет прогрессивно, но не все паразиты выживают, в противном случае кровь в короткий срок была бы заполнена плазмодиями. Известно^ количество паразитов фагоцитируется белыми кровяными клетками; другая часть находится под влиянием или погибает от гуморальных воздействий. Необходимо отметить, что механическая пропорция между количеством паразитов и интенсивностью заболевания не имеет общей силы потому, что вирулентность самих плазмодиев и состояние организма малярика подвержены колебаниям. Этим и объясняются такие парадоксальные случаи, когда малярия протекает очень тяжело при скудном количестве паразитов в периферическом русле крови и обратно: бывали примеры легкого течения болезни при большом количестве паразитов.
Повидимому, малярийным плазмодиям свойственны некоторые сезонные изменения их особенностей. При заражении людей PL ѵіѵах в процессе маляриотерапии через , уколы анофелес в зимнем квартале Boyd, Kitchen и Muench наблюдали меньшее количество заражений с более длительной инкубацией и более коротким клиническим течением; рецидивы появляются в первые восемь недель после прекращения первичных приступов. В (летнем же квартале было много успешных заражений при наименьшей инкубации и Han6c^eeJ длинном течении болезни; наибольшее количество рецидивов наблюдалось через 24 недели после первых приступов.
Неодинакова и индивидуальная восприимчивость людей к заражению- малярией.
Вторичный латентный период связан с прекращением шизогонального размножения, причем паразиты или вовсе исчезают из периферической крови, или находятся в ней в виде гаметоцитов.
Если впрыскивать людям кровь маляриков, содержащую в себе только одни гаметоциты, заражения малярией не произойдет; обратный эффект получится при впрыскивании крови с шизонтами.Отсюда делается заключение, что гаметоциты не заразительны для человека и поэтому не являются сами по себе болезнетворными для него. Если характеризовать значение всех стадий развития малярийного плазмодия в крови человека, то можно сказать, что возбудителями экспериментальной малярии являются шизонты и мерозоиты, но не гаметоциты.
Длительность вторичного латентного периода может быть весьма значительна и исчисляться годами (2—4—6—10 лет). Человек может забыть, что у него когда-то была малярия и вместе с тем может внезапно снова заболеть этой болезнью не за счет нового заражения (так называемой реинвазии), а в порядке возврата ранее бывшего заболевания, т. е. рецидива.
По данным Шаудинна, рецидивы объясняли партеногенозом макро- гаметоцитов, которые долгое время остаются в недеятельном состоянии, а затем под влиянием различных инсультов (переохлаждение организма, циэтетическое воздействие, травмы, кровотечения, воздействие ультрафиолетовых лучей солнца и мн. др.) начинают размножаться делением без предшествующего процесса слияния с микрога- метоцитами. Возникающие таким путем дочерние особи внедряются в эритроциты и в дальнейшем ведут себя как нормальные шизонты. вызывая приступы малярии.
Картина такого девственного размножения, когда макрогаметоци- ты будто бы дают некоторое количество мерозоитов и остаточное тело, морфологически не отличима от так называемых двойных инвазий. Последние характеризуются внедрением в один и тот же эритроцит двух паразитов на разных стадиях их жизни, причем каждый паразит ведет независимо от другого «свою линию жизни» — например, делится, когда другой находится на положении гаметоцита или растущего шизонта (табл. V, рис. 11; рис. 87, В, 8). Поэтому гипотеза девственного размножения макрогаметоцитов не имеет под собой базы, а многие авторы объясняют рецидивы тем, что в организме в скрытом состоянии сохранились шизонты, которые при подходящих к тому условиях начинают снова размножаться со всеми вытекающими отсюда последствиями.
Малярия может прекратиться у некоторых людей в естественном порядке, т.
е. без специального лечения. Такое выздоровление называется спонтанным. Абсолютным выздоровлением надо считать полное исчезновение малярийных паразитов из организма человека. Нормальными сроками этого процесса считают 3—5 лет.Рецидивы могут быть вызваны искусственно — рентгенизацией селезенки, впрыскиванием адреналина и др. (способ провокации малярии) для последующего уничтожения паразитов лечением.
От рецидивов следует отличать реинфекцию (точнее — р е и н- вазию), или повторное (т. е. новое) заражение человека, который^ в силу тех или других причин вовсе освободился от первичной инфекции или у которого она затихла; однако такое разделение на практике не всегда возможно провести в смысле установления диференциальной диагностики у больного.
Наконец, суперинфекцией (суперинвазией) называют добавочное или последовательное заражение малярийным плазмодием того же самого или другого вида паразита (например, суперинфекция РІ. ѵіѵах на PI. falciparum).
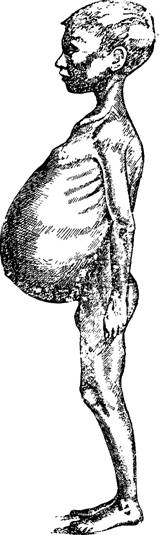
Рис. 91 —Увеличение селезенки у малярика. По Широкого- р о'в у.
Одним из характерных проявлений малярии является увеличение селезенки, край которой в норме не выходит за границы левых ребер. Увеличение селезенки при хронической малярии свидетельствует не только о зараженности человека малярией, но и о сопротивляемости организма этой инфекции. Показательны примеры тяжелых и смертельных случаев малярии, при которых селезенка сохраняла свои нормальные размеры.
Известны случаи увеличения селезенки до 6,4 кг весом.
При тропической малярии селезенка увеличивается не в такой степени, как при терциане.
При общем учете распространения малярии отмечается ее сезонность, проявляющаяся резким поднятием заболеваемости в определенные сезоны года. Случаи заболевания малярией могут время бт время наблюдаться круглый год, но все же для малярии типична сезонность, связанная с теплым периодом года. О ходе заболеваемости малярии судят по обработке материалов учета заболеваемости.
Обычно учитывается валовая заболеваемость. Статистические данные оформляют в виде месячной кривой, отражающей календарное течение заболеваемости. Рассмотрение больших материалов из разных в широтном расположении местностей СССР позволяет сгруппировать малярийные кривые в две категории. Для ряда местностей типична кривая, дающая максимальный подъем в первую половину лета (рис. 92); такой же ход заболеваемости характерен для северной половины территории. В южной половине кривая малярии дает второй — большой осенний подъем (рис. 93). Наблюдаются и различные отклонения этих кривых. Весенний подъем заболеваемости малярией обусловливается как рецидивами, так и первичной заболеваемостью с длительной инкубацией терцианой. Осенний подъем зависит от свежих случаев терцианы, от рецидивов и первичных заболеваний тро іической малярией. При раздельном учете первичных заболеваний и рецидивов терцианы в южных (местностях кривая заболеваемости малярией характеризуется также двувершинностью. При учете заболеваемости необходимо отмечать срок нахождения или время прибытия малярика в соответствующее место, где он подвергается учету. Валовые статистические данные имеет часто относительное значение для малярийной характеристики места учета, отражая лишь фактическое наличие маляриков, часть которых могла приехать в точку учета, заразившись малярией совсем в другом месте (подробнее см. стр. 180, 181).При регистрации маляриков необходимо точно различать первичное заболевание и первичное посещение. Первичным маляриком является человек, впервые^-заболевший малярией. Первично обратившимся считается всякий малярик,
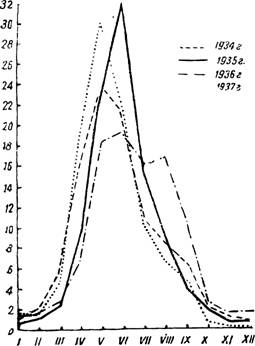
Рис. 92 — Северный тип кривой малярии (по заболеваемости в Ленинградской области в 1934—1937 гг., в процентах к годовой). По А. Якушевой.
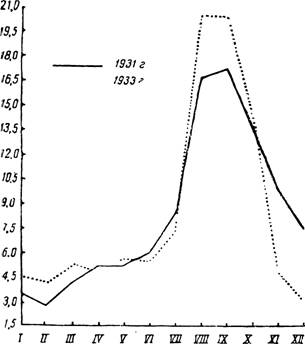
Рис. 93 — Южный тип кривой малярии (по заболеваемости в Дагестанской АССР в 1931, 1933 гг., в процентах к годовой). По А. Якушевой.
который впервые обращается в учетный год в лечебное учреждение из-за малярии. Первично обратившимися могут быть первичные малярики, малярики-рецидивисты и малярики с реинфекцией. Рецидивирующие малярики являются недолеченными хрониками; наличие их в данном месте является показателем необходимости противорецидивной профилактики (см. стр. 196). Точное установление первичных для данной местности заболеваний малярией говорит о том, что эпидемическая вспышка малярии не изжита, что диктует применение специальных мероприятий. Сравнение малярийных кривых по годам дает представление о росте или затухании эпидемичности малярии в месте ее учета (рис. 94, 95).
Кривые малярии строятся по абсолютному числу маляриков, или по расчету на 10 тыс. населения, или по индексам.
Для характеризования зараженности населения и малярийности местности определяют некоторые индексы, а' именно:
1. Паразитный индекс — процент паразитоносителей среди населения, обследованного микроскопированием окрашенных толстых капель; особое значение имеет определение процента гаметоносителей, ибо только эта категория маляриков заражает анофелес и, следовательно, является источником дальнейшего распространения малярии в районе.
2. Селезеночный индекс — процент лиц с увеличенной се-
лезенкой (рис. 96) по отношению к осмотренному населению; этот индекс является показателем распространения или экстенсивности малярии в обследуемом районе.
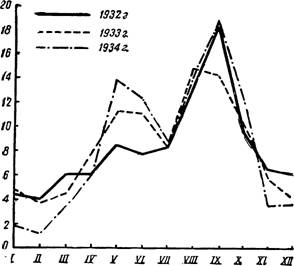
Рис. 94 — Двувершинная кривая заболеваемости малярией ^Куйбышевская обл. 1932—1934 гг., в процентах к годовой). По А. Якушевой.
Так как дети прежде всего заболевают малярией и реагируют на эту болезнь резче, чем взрослые, у которых может вырабатываться известная степень относительной невосприимчивости (стр. 180), то определение селезеночного индекса по возрастам имеет существенное значение; количественные показатели селезеночного индекса дзют более точную картину, если исходить из определения его у детского населения; маляриологи в Индии считают, что соответствующий индекс менее 10% характеризует местность как здоровую; 10 —
25% соответствуют умеренной малярийности; 25 — 50% свидетельствуют о сильной малярийности и, наконец, индекс выше 50% говорит о весьма тяжелой малярийности.
3. Индекс Росса иллюстрирует среднюю степень увеличения селезенки у обследованного населения, или интенсивность малярии. При осмотре всех обследуемых делят на четыре категории:
а — с нормальной по размерам селезенкой;
Ь — с селезенкой, выступающей из-под края ребер на 3 пальца;
с — с селезенкой, выступающей из-под края ребер на 4—5 пальцев;
d —с селезенкой, выступающей из-под края ребер более чем на 5 пальцев.
Индекс вычисляется по формуле:
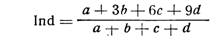
Под а, b, с, d подставляют численное обозначение соответствующей группы обследованных; при отсутствии лиц с увеличенной селезенкой индекс Росса равняется единице; повышение его до 5 и более свидетельствует о весьма тяжелом поражении населения малярией. Для определения увеличения селезенки удобно пользоваться способом С. Канделаки.
Индекс Росса, выраженный в механистической форме, не всегда точен, ибо селезенка при тропике увеличивается в меньшей степени, чем лри терциане, что в известной мере уменьшает значение этого вида.
Для наибольшего уточнения характеристики малярийности определяют эндемический индекс, под которым подразумевается общий процент лиц с объективными признаками малярии (наличие только паразитов в крови; наличие только увеличения селезенки и одновременное наличие обоих признаков).
Паразитный и селезеночный индексы, каждый в отдельности, не
гарантируют уловления всех маляриков среди обследуемых, так как могут быть паразитоносители без увеличения селезенки, и обратно:
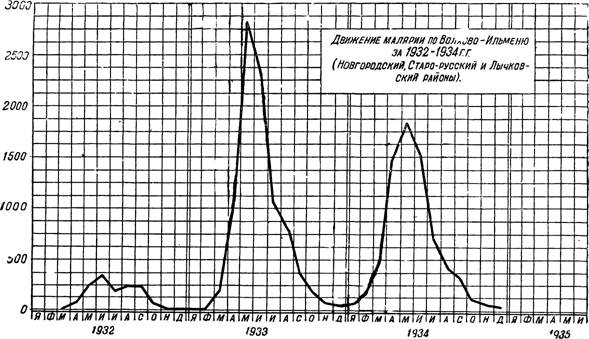
Рис. 95 — Движение малярии по Волхово-Ильменю за 1932 — 1934 гг. По 1’. Шубу и В. Федорову.
бывают люди с увеличенной селезенкой, но без паразитов в крови в момент исследования. Необходимо также помнить, что увеличение селезенки бывает и не у малярийных больных (например, кала- азар и др.).
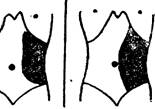
Рис. 96 — Колебания в величине селезенки. (Пастеровский институт в Алжире).
Необходимо отметить, что «одномоментное» обследование населения на малярийность не дает исчерпывающе точной картины, так как малярийные паразиты могут временами (или на более или менее долгий срок) исчезать из крови; селезенка также может значительно колебаться в размерах у одного и того же человека (влияние лечения и другие причины; рис. 96).
Наконец необходимо помнить, что зараженность обследованного населения малярией не всегда стоит в полном соответствии с малярийно- стью местности, так как могут быть приезжие малярики, которые заразились малярией не в месте их обследования. Поэтому важно дифе- рейдировать коренное население от пришлого и среди коренного населения обследовать особенно тщательно детей, как элемент наиболее оседлого характера, могущий, следовательно, наиболее точно характеризовать малярийность пункта работ.
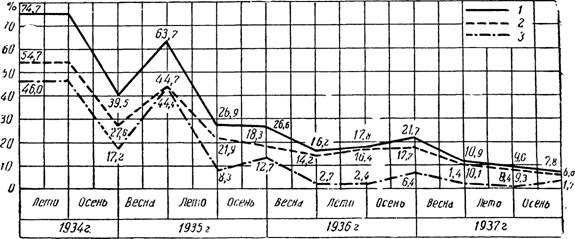
Рис* 97 — Падение индексов обследования на малярию, как показатель оздоровления малярийного очага (с. Хамидие Кабардинской АССР).
1 — эндемический индекс; 2 — селезеночный индекс; 3 — паразитарный индекс.
По М. Рашиной и А. Хованской.
Падение индексов при повторных обследованиях на малярию одной и той же точки или района является показателем оздоровления малярийного очага (рис. 97).
ИСТОЧНИКИ ЗАРАЖЕНИЯ МАЛЯРИЕЙ
Заражение от человека. В естественных условиях человек может заразиться малярией от человека в порядке внутриутробной инвазии: от матери, больной малярией, плазмодии могут пройти сквозь плаценту и через сосуды пупочного канатика попасть в плод; такое прохождение зараженных эритроцитов от матери в плод мыслимо при различных поражениях плаценты (например, сифилис), которая теряет вследствие этого свои барьерные свойства. Еще легче может произойти смешение зараженной крови матери с кровью плода при отторжении плаценты. Наконец, мыслимо заражение во время самих родов при попадании крови , матери в перерезанную пуповину или ссадины кожи плода.
Указанные способы заражения малярией называются к о н генитальными, а самая форма малярии врожденной (ложная наследственность). ^
Искусственное заражение людей малярией легко достигается впрыскиванием крови малярика с шизонтами человеку, причем для успеха дела бывает достаточно ничтожного ее количества (капля или даже меньше). Искусственная или экспериментальная малярия широко применяется для лечения прогрессивного паралича (Wagner ѵ. Jaurreg) (пиретотерапия). Бывают случайные заражения малярией при инъекциях (например, анестезия при экстракции зубок.
шприцем, которым ранее производилась инъекция в десны малярика и ‘без достаточной дезинфекции шприц был использован для здорового от малярии человека и т. п.) При лечебной практике накопился побочный биологический материал, говорящий в пользу некоторого различия течения и особенностей естественной и экспериментальной малярии. Последняя, в частности, дает гораздо меньше рецидивов и с чрезвычайной успешностью поддается лечению хинином. Инкубационный период экспериментальной малярии при прививке крови малярика значительно короче, чем при заражении крови через укол анофелес; для возбудителя терцианы он в среднем равняется 10—12 дням против 17 дней (при колебаниях от 3 до 31 дня); для паразита квартаны— 14 дней — против 21 -(при колебаниях от И до 50 дней) и для тропики — 5 дней против 10—12 (при колебаниях от 2 до 14 дней).
Для экспериментального заражения малярией необходима кровь с шизонтами плазмодия; кровь только с гаметоцитами вовсе не заразительна. Количество вводимой крови не имеет существенного значения; достаточно бывает даже следов крови в игле шприца, но практически инъицируют 1—2 см3 дефибринированной крови.
Возможны случаи заражения малярией при переливании крови даже тогда, когда у донора микроскопически паразиты не обнаруживаются и когда в его анамнезе не значится явно перенесенной малярии.
Заражение через переносчика. В настоящее время точно установленным положением является признание специфическим переносчиком возбудителей различных форм малярии комаров рода Anopheles, что экспериментально установлено Россом и Грасси. Замечательно, что дикари Восточной Африки интуитивно связали представление о малярии с комарами, ибо и малярия, и комары ими называются одним и тем же словом «мбу» (Кох).
Кроме анофелес никаких других переносчиков малярийных паразитов не известно.
Когда Anopheles напивается крови малярика, то всасывает в себя различные стадии развития паразита. Мерозоиты и другие стадии бесполого размножения в желудке комара погибают, тогда как гаметы приступают к созреванию; из макрогаметоцита выталкивается часть ядерного вещества с протоплазмой (редукционное тельце, рис. 84, 14 а), причем он созревает в макрогамету или женскую особь паразита. Микрогаметоциты подвергаются резко бросающимся в глазт изменениям. Ядра их распадаются на несколько частей, подходящих к поверхности плазмодия, который начинает выпускать как бы длинные и тонкие псевдоподии или бичи ,(рис. 84, 14 Ь; рис. 87, А, 8). Эти отростки протоплазмы с ядром внутри отделяются от остаточного тела микрогаметоцита в качестве зрелых мужских особей или микрогамет. Они имеют вид бичей длиной 20—25 ^ при толщине всего в 1 ,и. Из каждого микрогаметоцита образуется от 4 до 8 подвижных змееобразных микрогамет и остаточное тело.
Интересно отметить, что микрогаметы могут образовываться из микрогаметоцитов вне тела анофелес и вне русла крови человека; их образование можно видеть под микроскопом в капле свежей крови под влиянием ее охлаждения.
Их-то и заметил впервые Лаверан, при микроскопировании капли крови малярика; по их колебательным движениям Лаверан дал открытому им возбудителю малярии название OscUlaria таіагіае. Следует отметить, что в связи с указанными выше особенностями полового цикла кокцидий (стр. 144) в сходном порядке начинают толковать ход созревания гамет малярийного плазмодия, усматривая таковое в период после слияния мужских и женских особей плазмодия (зиготиче- ская редукция вместо предварительно протекающей гамети- ческой редукции).
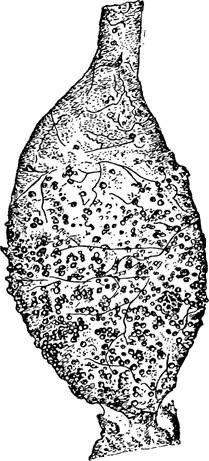
Рис. 98 — Желудок Anopheles maculipennis со множеством ооцист Plasmodium ѵіѵах. ПоСиманину.
Минут через 20 (и более — до 2 часов) после того, как кровь малярика попала -в желудок Anopheles, происходит оплодотворение, причем микрогамета, т. е. мужская особь, внедряется в макрогамету (в женскую особь — рис. 84, 16). Оплодотворенная макрогамета превращается в червеобразную зиготу или о о к и- н е т у (3—5 X 1°—24 Iй размером); последний термин в буквальном смысле значит «подвижное яйцо» или, в данном случае, подвижная женская оплодотворенная особь (рис. 84, 17). Мужское и женское ядро оокинеты сливают вместе.
Оокинета внедряется в стенку желудочного эпителия и проходит к его основанию, т. е. к его базальной перепонке.
Оокинета одевается оболочкой и начинает расти. По мере увеличения она подается наружу, выпячивает мышечную оболочку желудка и становится видной с его поверхности (рис. 98).
Эта стадия развитий называется ооцистой; внутри ооцисты ядра плазмодия энергично делятся. Вокруг ядра сгущаются участки протоплазмы, а сами ядра расходятся по периферии отдельных частей обособленной протоплазмы, называемых споро б ластам и, в которых развиваются веретеновидные спорозоиты.
В этом состоянии последние достигают 14 ^ длины при толщине на уровне ядра всего в 1 ja. Число их в каждой споро- цисте может превышать 10 тыс. В конце концов спорозоиты начинают плавать в полости ооцисты, которая к тому времени вздувается до максимальных пределов. Размеры ооцист колеблются, в зависимости от их зрелости, от 4—5 у до 60 и редко 70—90 у*. Количество ооцист может достигать 200—500.
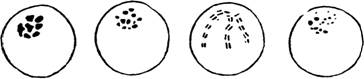
Ооцисты различных видов малярийного плазмодия отличаются по характеру отлагающегося в них пигмента (рис. 99). Для ооцист РІ. ѵіѵах характерно цепочечное расположение желтоватых или золотистых тонких и коротких палочек пигмента. В ооцистах PI. malariae пигмент имеет вид грубоватых зернышек, которые еще грубее у РІ. falciparum. Зато в ооцистах PI. ovale пигмент имеет вид нежных перекрещивающихся черточек.
Утончающаяся стенка ооцисты под давлением внутреннего содержимого лопается (рис. 84—24, 25). Спорозоиты выходят в полость тела комара и с током полостной жидкости .(гемолимфы) распространяются по всему его организму (рис. 84, 26). В груди они активно внедряются
в слюнные железы, вероятно, привлекаемые химиотаксически, проникают в слюнные клетки и сквозь них проходят в полость железы и в ее выводной проток (рис. 100, 101).
А В С п
Рис. 99—Различие пигмента в ооцистах малярийных плазмодиев.
А—тропической малярии; В — четирехднеяной малярии;
С — трехдневной малярии; D — птичьей малярии (Daniels).
Если теперь Anopheles со спорозоитами в слюне станет сосать кровь здорового человека, то он впрыскивает ему возбудителей малярии, т. е. спорозоиты. Последние, попав в кровь человека, внедряются в эритроциты, теряют в них веретеновидную форму и становятся
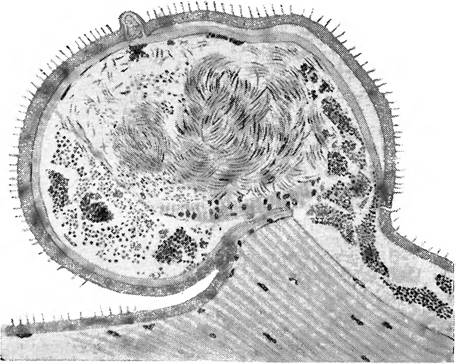
Рис. 100 — Продольный разрез спинки анофелес: в полости тела в гемолимфе видно огромное количество веретеновидных спорозоитов малярийного плазмодия. По Мюленсу.
молодыми шизонтами, которые растут и переходят к бесполому размножению. С этой частью их жизненного цикла мы уже ознакомились выше.
Человек явственно заболевает перемежающейся лихорадкой после инкубационного периода, , требующегося для превращения спорозоитов в шизонты и £ля размножения этих последних, результаты чего нам уже известны.
Таким образом замыкается полный жизненный цикл малярийного плазмодия — цикл, слагающийся из двух стадий: 1) жизнь в крови человека, характеризующаяся бесполым размножением плазмодия (шизогония) и подготовкой к дальнейшему половому размножению образованием гаметоцитов, и 2) пребывание в различных органах анофелес, связанное с половым размножением паразита и споруляцией его с конечным возникновением спорозоитов (спорогония).
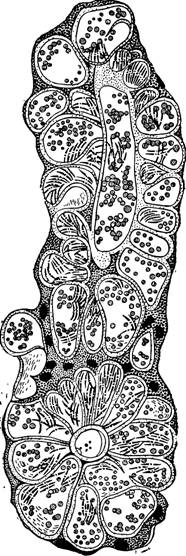
Рис. 101 — Спорозоиты в слюнной железе (на срезе). По М ю л е н с у.
Итак, оба хозяина малярийного паразита взаимно заражают друг друга только вполне определенными стадиями развития паразита: Anopheles инвазируется только гаметоцитами из крови человека, а этот последний — спорозоитами из слюны комара.
Низкая температура неодинаково действует на разные моменты жизни и на размножение малярийных паразитов в теле Anopheles. Наиболее уязвимы начальные стадии размножения плазмодия. Низкая температура уне позволяет оокинете проникнуть через стенку желудка и превратиться в ооцисту (Jancso). Если температура была достаточно высока, чтобы копуляция гамет плазмодия могла произойти, то дальнейшее развитие паразита протекает и при более низкой температуре (Qrassi); -f-5,5°C не вредят уже сформировавшимся ооцистам (We- nyon), что дает основание предполагать возможность зимовки незрелых цист в комаре и доразвитле их весной (Македония). J. James дал еще более убедительные данные в пользу такой возможности. В его многочисленных экспериментах (Англия) ооцисты на желудке и спорозоиты в слюнных железах не погибали, если комаров держали при -4-4—5,5°С три недели или при температуре около точки замерзания—
6 дней. Некоторые комары Ап. maculipennis жили со спорозоитами в слюне и сохраняли свою заразительность от 29 до 92 дней после появления в их слюнных железах спорозоитов.
Несомненно, что понижение температуры задерживает развитие ооцист, которое подвигается дальше, как только температура повышается до благоприятных норм. Возможно также, что плазмодий может переживать в организме анофелес легкие зимы.
Ооцисты PL falciparum выдерживают температуру 1,7° С не более суток, PL ovale заканчивает развитие ооцист в An. maculipennis при температуре 24—25° С в течение 16 дней против 10 дней, потребных в тех же условиях как для PL vivax (James, N'.col, Shute). А. Лисовой удалось заразить An. maculipennis и An. superpictus штаммом PL ovale из-под Уфы и наблюдать появление спорозоитов. На 22-й день при вскрытии были обнаружены крупные, непрозрачные, бедные мелкозернистым пигментом цисты без спорозоитов, мелкие прозрачные цисты и «множество спорозоитов в слюнных железах. Однако попытки заражения пяти человек кормлением на них Anopheles,* зараженных PI. ovale, дали отрицательный результат.
Сроки- развития малярийных плазмодиев в комаре Anopheles (в сутках)
1. Anopheles macuiipennis

Более детальные данные по длительности спорогонии у Anoph. macuiipennis (добыты Б. Николаевым)
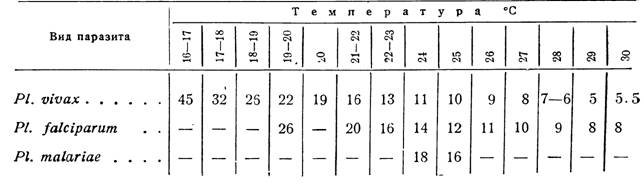
Ходукин с Лисовой и Симанин экспериментально доказали, что An. elutus (Ташкент) и An. macuiipennis (Барнаул), взятые с зимовок, могут в тепле зимой заражаться малярийными плазмодиями. Поэтому в известных условиях вполне возможно и зимнее первичное заражение человека малярией.
Присутствие в слюнных железах комара подвижных спорозоитов не обусловливает обязательности заражения малярией каждым таким комаром любого • человека .(Bruce Мауле), в то же время Mitzmain наблюдал заражение одним и тем же экземпляром Anopheles девяти человек из двенадцати в 14 дней. Для заражения малярией может быть достаточно только одного укола инфицированного анофелес. Другие авторы указывают, что в большинстве случаев необходимо повторное сосание крови человека анофелес и что для заражения человека требуется от 13 до 31 укола (Hanschell). *
Спорозоиты паразитов терцианы и тропики могут жить в слюнных железах анофелес до 4 месяцев, но заразительность комаров сохраняется лишь в течение 2—3 месяцев; снижаться же она начинает уже 1 2
[1] За это время при тех же условиях ооцисты PL falciparum еще недостаточно созрели.
[1] Эта температура, по Jancko, является нижним пределом развития плазмодия тропической малярии в комаре.
через месяц для Р1. ѵіѵах и, начиная с 20-го дня,— для PL falciparum, считая этот срок со времени проникновения спорозоитов в слюнные железы комара.
Влажность имеет меньшее значение для развития плазмодия, нежели температура. Из других факторов некоторые побочные или случайные обстоятельства также могут исключить возможность развития плазмодия в желудке комара. Такое действие оказывают различные кислые растительные соки, всасываемые самкой комара. Они создают неблагоприятную для плазмодия реакцию среды в желудке анофелес, и еще СеШ заметил, что в тех местностях Италии, где культивируются томаты, малярия относительно редка. Питание анофелес кровью животных и птиц может также делать их невосприимчивыми к заражению плазмодием.
James находил не восприимчивых к заражению комаров в природе. Их необходимо было выдерживать некоторый срок исключительно на питании человеческой кровью, чтобы потом можно было бы с успехом инвазировать плазмодием.
На заражаемость комара, кроме уже указанных моментов, влияют количество выпитой им крови, относительное богатство ее гаметоцита- ми, достаточное количество микрогаметоцитов, которых вообще развивается меньше, чем макрогаметоцитов, и факт предварительного лечения малярика хинином или синтетическими препаратами. Хинин губит гаметоциты PL ѵіѵах и не действует на PI. falciparum, поэтому анофелес могут быть заражены плазмодиями тропической малярии на больных, принимавших хинин, тогда как при аналогичных условиях PL ш~ ьах развивается в комаре редко (Wenyon).
Важным обстоятельством является факт потери микрогаметоцитами способности созревать, т. е. эксфлягеллировать после приема маляриком 0,03 синтетического хинолинового препарата плазмохина (М. Bahr), что подтверждено многочисленными наблюдениями и опытами. Такое гамостатическое действие препарата, пресекающее подготовку к половому размножению, ведет к тому, что анофелес, пьющие кровь таких маляриков, сами не заражаются малярийными паразитами; это же в конечном результате при достаточно экстенсивном применении плазмохина способствует значительному снижению заболеваемости малярией в данной точке.
Плазмоцид (советский аналог плазмохина) обладает аналогичными свойствами. Например, в Дагестане при лечении плазмоцидом 975 человек отмечено снижение индекса зараженности анофелес по сравнению с участком, где производилась хининйзация, и за весь сезон гаметоциты тропической малярии были найдены лишь у 6 человек (Пикуль* Сергиев и Тибурская).
Однако указанное гамотропное действие плазмоцида имеет предел. При дозе 0,12 в день в отношении РІ. ѵіѵах плазмоцид действует в течение 2Ѵг суток после приема последней дозы. При дозе 0,09 действие ее прекращается ко вторым суткам. При тропической малярии, леченной плазмоцидом (0,09 в течение одного дня) и акрихином (по 0,3 в течение пяти дней), больной остается незаразительным для комаров не менее шести дней после последнего приема плазмоцида (Тибурская).
Эти сроки важны для установления рациональных норм приема лечебных препаратов (см. ниже инструкцию по применению синтетических препаратов).
Возвращаясь к вопросу о связи богатства 'крови малярика гаметами и заражаемости анофелес, пьющего его кровь, отметим, что
An. maculipennis может быть заражен PL falciparum даже при наличии всего 10 гамет в 1 мм3 крови (Вещезеров). Число развивающихся ооцист не прямо пропорционально количеству проглатываемых гамет, а меньше. Mitzmain наблюдал, что большее число их (Р/. falciparum) проходит через кишечник и выбрасывается с испражнениями комара, и лишь малый процент развивается дальше в его желудке. Наряду с вышеприведенными результатами Вещезерову вовсе не удавалось заразить Anopheles кровью человека с 372 гаметами PL mala- гіае в 1 мм3. Имеет, повидимому, значение и возраст гаметоцитов; по Крэгу (Craig), они должны быть не моложе 12 дней. Повторность кормления данного анофелес на данном гаметоносителе увеличивает шансы заражения комара малярийными паразитами.
Паразиты квартаны труднее передаются через анофелес, что повело к неоправдавшемуся предположению Маршу, будто бы этот вид малярийного плазмодия имеет какого-то другого переносчика, а не анофелес. Опыты передачи Р/. таіагіае давали положительный результат с А. quadrimaculatus (США, М. Broyd и Stratman — Thomas) и A. maculipennis alroparvus (de Buck).
Плазмодиями четырехдневной малярии удачно заражала в Сухуми An. maculipennis, An. bifurcatus и An. nigripes Гендельман.
Попытки некоторых исследователей заразить Anopheles на больных, привитых кровью малярика, кончались неудачно. W. York и W. Wright были счастливее в этом отношении. Они показали что 53— 54 пассажа Р/. ѵіѵах через организм людей в течение ЗѴг лет нисколько не ослабили способности плазмодия к гаметообразованию и инфицирующей силы гаметоцитов. Анофелес заражался на последнем больном и своим уколом заражал людей, никогда не болевших малярией.
Заражение анофелес плазмодиями малярии, видимо, не влияет на среднюю продолжительность его жизни. Однако, по показаниям других авторов, Anopheles maculipennis, зараженные малярийными паразитами, летом быстро погибают; таким образом, до проявления потребности повторного сосания крови человека доживает лишь небольшое число самок, что уменьшает . шансы заражения людей малярией (Голландия, de-Buck и A. Swellengrebel). Очевидно, что длительность жизни зараженных анофелес в различных наблюдениях зависела не только от самой зараженности их плазмодиями малярии, но и от других факторов, совокупное действие которых ведет к разным результатам в отношении продолжительности жизни анофелес.
Не все виды анофелес являются в данном месте переносчиками любой формы' малярийного паразита. Возможно, что разные виды малярийных комаров переносят преимущественно разные виды плазмодия (Wenyon). Главным носителем Р/. ѵіѵах в Македонии Wenyon считает An. maculipennis, а для PI. falciparum — An. super- pictus. По Kinoshita,— PI. falciparum на Формозе переносится An. listoni и, в противоположность PI. ѵіѵах, вовсе не развивается в Ап. hyrcanus.
Эпидемиологическое значение разных видов анофелес устанавливается как вышеизложенными опытами с искусственным заражением их плазмодиями малярии, так и обследованием естественной зараженности плазмодиями в данной местности; для обнаружения последней вскрывают множество самок, ищут в них ооцисты и спорозоиты (последнее — дело более трудное, чем первое) и определяют процент зараженности комаров.
Таким способом определяется плазмодийный индекс анофелес, выводимый по отношению наличия ооцист на поверхности выпрепа- рированного желудка (ооцистный индекс) и применительно к присутствию спорозоитов в слюнных железах (спорозоитный индекс). Учитывая большую техническую легкость массовых вскрытий анофелес для определения зараженности их ооцистами, гораздо чаще плазмодийный индекс определяют только в его оодистной части. Спорозоитный индекс обычно меньше ооцистного, так как не из всех цист развиваются спорозоиты; но при несоответствии условий, необходимых для развития ооцист и для пребывания спорозоитов в слюнных железах, отношение обоих индексов может стать обратным (например, во второй половине сентября при понижении температуры. Лисова и Эскин, рис. 102).
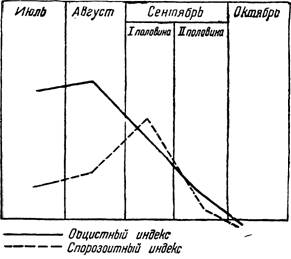
Рис. 102 — Кривая спорозоитного и ооци- стного [индекса по месяцам в Той-Тюбе (под Ташкентом). По А. Л и с о"в о й и В. Эскину.
Годовой ооцистный индекс для Сухуми у An. maculipenniSy взятых из жилых помещений, достигает 7,6о/о, из скотников—
1, 7, из подвалов и уборных —0. Наибольшее количество зараженных анофелес взято из жилищ маляриков. (Л. Рейнгард и Забудько- Рейнгард) при максимуме — на август.
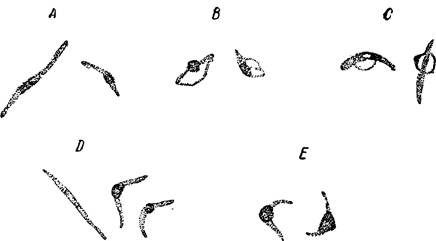
Рис. 103 — Дегенерация спорозоитов в слюнных железах.
А — An. superpictus (утолщение); В — С — (дегенерация); D — An. elutus (от нормы до утолщения и укорочения); Е— An. maculipennis (дегенерация). По М. Barber.
В некоторых спорозоитах наблюдается несколько глыбок хроматина, что считается делением ядра спорозоита. Наряду с нормальными спорозоитами наблюдаются спорозоиты, необычно утолщенные и дегенерирующие в различной степени (рис. 103). Этот процесс бывает у разных видов анофелес. В одной .и той же слюнной железе могут быть спорозоиты и нормальные и дегенерированные в разной степени. Явление дегенерации спорозоитов «в отдельных наблюдениях свя зывают с временем года (например, у A. mac. etutus летом дегенерация наблюдается реже, чем зимой; это явление весьма часто наблюдается
на Кипре), с возрастом спорозоитов и с другими причинами. Несомненно, что дегенерирующие спорозоиты теряют заражающую способность; это может вести при соответствующих условиях к ослаблению передача малярии в данной местности или к уменьшению •эпидемиологического' значения вида анофелес, которому в сильной степени присущ рассматриваемый феномен .(Barber).
Swellengrebel с сотрудниками наблюдал дегенерацию спорозоитов у А. maculipennis в Голландии во время зимования зараженных самок*, начиная с ноября; в январе уже не оставалось нормальных спорозоитов. Ооцисты также дегенерируют, что начинается раньше дегенерации спорозоитов.
Экспериментальное заражение разных видов анофелес плазмодиями малярии в СССР производилось рядом исследователей. Весьма удачны были опыты Вещезерова в районе Адлера с заражением Ап. maculipennis и An. bifurcatus. Оба вида были резко восприимчивы к PI. falciparum. Удавалось также заражение их и Р1. ѵіѵах. В первом случае развитие плазмодиев было прослежено до внедрения спорозоитов в слюнные железы анофелес. Ходукину в единичных случаях удалось найти цисты на желудке An. pulcherrimus (Голодная степь) после заражения комаров РІ. ѵіѵах и PL malariae. Авербух удалось заразить, до 40э/о этого вида комара, но естественное заражение его малярийными паразитами в Туркестане пока обнаружено только нахождением спорозоитов в слюнных железах комаров, взятых в августе из жилых помещений одного из поселков Голодной степи (Завадский).
Симанин не мог заразить среднеазиатских An. hyrcanus плазмодием тропической малярии. 136 подопытных комаров дали отрицательный результат, тогда как из 41 комара видов An. maculipennis ѵаг. sacharovi, An. pulcherrimus и An. superpictus заразились 34 комара,, соответственно в следующих процентах: 90, 84 и 77.
Лисовой также не удалось заразить An. hyrcanus паразитами тропической малярии; в другой серии опытов с PL ѵіѵах из 17 комаров заразились три (Ташкент). В Биробиджане Гуцевичу удалось заразить двух комаров из 26, также РІ. ѵіѵах, но на Дальнем Востоке и в Биробиджане, где имеется единственный вид А. hyrcanus, роль его в эпидемиологии малярии при настоящих условиях мала. Такое же мнение высказывают о соответствующей роли этого вида анофелес в эпидемиологии малярии в Приазовских плавнях (Данилова и Будышко).
An. nigripes в условиях эксперимента заражается всеми тремя видами малярийного плазмодия (Гендельман).
Недавно найденный в фауне СССР An. atgeriensis подвержен заражению плазмодиями тропической малярии (А. Лисова).
Естественная зараженность анофелес плазмодиями — плазмодийный индекс,— превышающий 10°/о, считается исключительной редкостью (Wenyon). Зараженность анофелес колеблется по месяцам года, что видно из нижеприводимых данных (см. табл, на стр. 179).
В Украинском протозойном институте из 560 вскрытых комаров зараженных было два. М. Парцванидзе в Мингрелии нашел только одного зараженного анофелес на тысячу. Топорков и Ефремов в Астрахани нашли ооцисты у 7 An. maculipennis из 187 вскрытых самок (август, сентябрь). Из 4490 An. maculipennis, вскрытых в течение четырех лет в Днепропетровске (В. Горицкая), малярийными плазмодиями были заражены 33 комара. Важно подчеркнуть, что в жилых строениях, хозяйственных постройках и в помещениях для скота наибольший процент зараженных анофелес был обнаружен в домах маля-
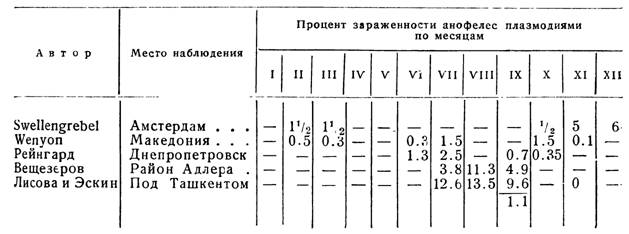
риков (например, 4 зараженных анофелес на 52, и далее: 4 из 40, 4: из 16). Первый зараженный анофелес был обнаружен 29 мая со зрелыми цистами; наиболее позднее нахождение комара с цистами и спорозоитами пришлось на 14 ноября. Отмечено также, что процент зараженных комаров, при прочих равных условиях, увеличивается в помещениях со скученным населением, живущим в антисанитарной обстановке и не охраняемом от нападения комаров (Лисова и Эскин).
В конечном счете количество инфицирующих анофелес зависит в каждом частном случае от количества комаров, пьющих кровь человека, количества гаметоносителей и количества анофелес, выживающих до проникновения спорозоитов в слюнные железы анофелес. Пример расчета количества зараженных анофелес возьмем из данных Данилова и Будышко (плавни Приазовья). В станице Тривенской анофелес, пьющих кровь человека, 22%, гаметоносителей — 3% и 16% зараженных анофелес доживают до появления спорозоитов. Общий процент зараженных комаров — 0,01 от всей комарийной популяции, т. е. 1 на 1000, что близко соответствует данным вскрытия анофелес.
Хотя зараженные плазмодиями комары и являются в общем редкостью, но каждый из них может повторно заразить несколько человек. Даже после сосания 30—40 человек в слюнных железах анофелес остаются многочисленные спорозоиты (James). Очевидно, эти органы время от времени пополняются спорозоитами, выходящими из последовательно созревающих, и лопающихся цист.
В общем эпидемиологическое значение различных анофелес в данной местности зависит, помимо наличия маляриков, от длинного ряда факторов: количества гаметоносителей, известной степени антропофилии комаров (см. главу о комарах), обстоятельств, благоприятствующих половому размножению плазмодиев (температура, влажность, реакция среды и химический состав содержимого пищеварительной трубки анофелес), склонности данного вида анофелес заражаться той или иной формой малярийного паразита, плотности анофелес, от длительности жизни насосавшегося малярийной кровью комара (обстоятельства, имеющего особо важное значение), от восприимчивости людей* к заражению малярией. Под индексом плотности анофелес понимается среднее количество анофелес на сбор в той или другой постройке (хлев, дом, погреб и др.). Передача малярии анофелес до известной степени пропорциональна плотности анофелес, конечно, при осуществлении основного условия — литания анофелес кровью человека. Имеет также значение вид анофелес. Например, в Биробиджане,, в Дальневосточном крае и в. Приморье единственным известным пока: видом является An. hyrcanus. 'Местная заболеваемость малярией невелика, что зависит по линии переносчика от комбинации следующих обстоятельств: 1) слабая восприимчивость An. hyrcanus к заражению плазмодиями малярии, 2) малое количество- перезимовывающих особей и позднее начало их размножения, 3) краткость периода активной деятельности большинства комаров, 4) относительно слабый контакт с человеком (А. Гуцевич).
Трудно сказать, существуют ли люди, абсолютно не восприимчивые к заражению малярией, но несомненно, что некоторые весьма стойки в указанном отношении. Несомненно также, что с возрастом коренное население малярийной местности становится более выносливым к малярии, и хроническая инфекция протекает гораздо легче, чем у недавно прибывших людей. "Повидимому, вырабатывается относительный нестерильный (т. е. не связанный с полным исчезновением паразитов из организма) иммунитет. Однако среди, казалось бы, невосприимчивого коренного населения бывают сильные обострения малярии, когда в данную местность вливаются незараженные пришельцы. Последние заражаются малярией; вирулентность местных штаммов паразитов, вероятно, усиливается при циркуляции их через свежих людей; благодаря этому паразиты оказывают повышенное влияние' на коренное население, которое добавочно заражается через комаров анофелес. Аналогичное значение имеет приезд гаметоносителей из других местностей; при этом происходит внесение нового штамма малярийных плазмодиев с большей вирулентностью для коренного населения, у которого уже выработалась известная невосприимчивость к местной малярии.
Наибольшее значение в распространении малярии в СССР играет An. maculipennis; в Средней Азии и на Кавказе такую же роль играет An. superpictus. Эпидемиологическое значение An. hyreanus и Ап. риі- cherrimusy повидимому, гораздо меньше. An. bifurcatus и An. plumbens, в силу своих экологических особенностей, едва ли играют особо заметную роль в распространении малярии, но в районном отношении значение их может быть вполне актуальным (санатории, стройки и др.). В настоящее время изучается эпидемиологическое значение недавно обнаруженного в СССР An. algeriensis (Средняя Азия, Кавказ), который, по Sergent, является переносчиком малярии.
Еще по теме УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ:
- Паразитические черви, проникающие в организм человека активно \
- Передача паразитических червей через посредство насекомых-переносчиков.
- Паразитические самцы рабдитовидного типа
- Б. Механическая (не специфическая передача яиц и личинок паразитических червей насекомым и-пере носчиками
- А. Специфическая передача личинок паразитических червей насекомы ми-переносчикам и [30] [31]
- Тип Protozoa — простейшие
- Простое окрашивание
- Простой герпес
- Простейшие (общая характеристика)
- Культивирование простейших кишечника
- Учение о фагоцитозе
- Просто гигиена.
- Учение макробиотиков
- Глава 1. Общее учение о болезни
- 2. Учение о стрессе
- Просто пришло Время ...
- 11.3.1.6. Энцефалит, вызванный вирусом простого герпеса