Описания жизненных циклов
Boophilus
Всем 4 видам этого рода свойствен однохозяинный тип развития. Наиболее детально он изучен у В. microplus, первоначальной родиной которого была Юго- Восточная Азия и откуда он в исторический период распространился по всей тропической и частично влажной субтропической зонам.
У В. microplus в условиях постоянной высокой температуры и влажности, характерных для тропического климата Бразилии, Кубы или Северной Австралии, от отпадения напитавшихся самок до появления активных голодных личинок проходит всего около 4 нед. Если к этому добавить 3—4 нед питания и линек на теле хозяина, то время развития 1 генерации составляет около 8 нед. Фактические сроки жизни одного поколения увеличиваются в зависимости от сроков встречи с прокормителем голодных личинок (предельный срок жизни до 107 сут) еще на 1—4 нед. В среднем за год успевает развиваться 5—6 последовательных генераций клещей. Однако в случае с непрерывным циклом развития само понятие генерации представляется крайне неопределенным. Одновременно на теле хозяина находятся клещи разных фаз развития, на разных сроках питания. В связи с возможностью жизни голодных личинок до 1—2 мес питающиеся на одном хозяине и в одно время клещи могут принадлежать к родительскому и дочернему поколениям. Отпадение напитавшихся самок круглый год при отсутствии синхронизирующих факторов для сроков яйцекладки, эмбриогенеза и активности личинок в еще большей степени стирает всякие границы между поколениями. Реально в условиях тропического климата природные популяции В. microplus представлены совокупностями особей разных календарных возрастов без каких-либо генеративных разрывов.В субтропическом климате прохладная зима прерывает или сильно замедляет развитие непаразитических стадий, а весной и осенью для их завершения требуется 10—12 нед. В Аргентине в подобных условиях в течение 1 года успевают развиться 3—4 поколения.
Границы между ними обнаруживаются по нескольким последовательным пикам отпадения напитавшихся самок. Наиболее выражен первый весенний пик, который обусловлен одновременным нападением активизировавшихся с весенним потеплением личинок. Последующие пики становятся более пологими из-за увеличения различий в сроках развития непаразитических стадий в отличающихся по микроклимату местообитаниях. Десинхронизация развития связана также с разной продолжительностью жизни голодных личинок до встречи с прокормителем. В конце осени—начале зимы развитие напитавшихся самок и их кладок замедляется или прекращается, голодные личинки становятся неактивными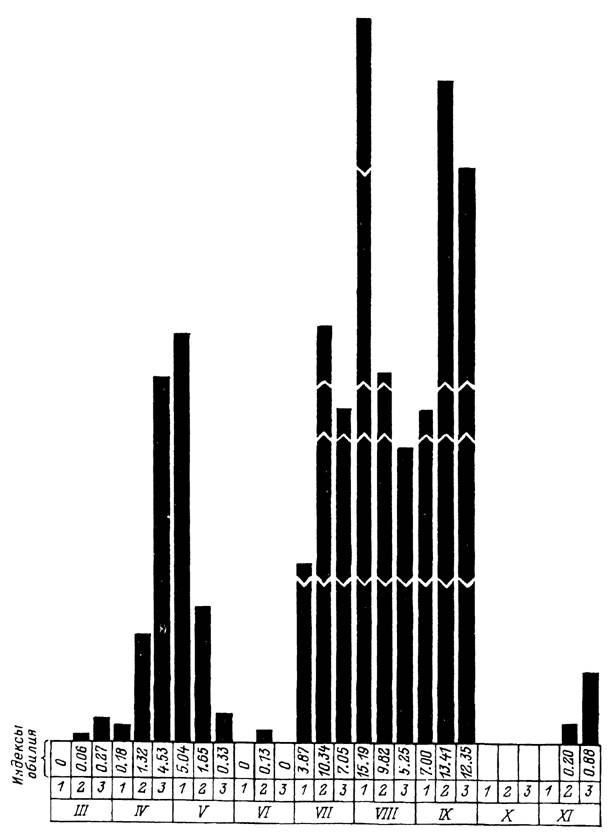
Рис. 41. Среднедекадные индексы обилия клеща Boophilus annulatus на крупном рогатом скоте в Гиссарс- кой долине Таджикистана (по: Потоцкий, 1945).
1—3 — декады; III—XI — месяцы наблюдений.и происходит частичное или полное освобождение скота от клещей (Nunez et al., 1985; Sutherst et al., 1988a).
Сходный тип развития свойствен имеющему средиземноморское происхождение В. annulatus. На юге Казахстана и в Средней Азии зимние температуры ниже 5—10 °С, полностью исключающие возможность развития этих клещей, сохраняются в течение 3—4 мес. В этот период на пастбищах перезимовывают голодные личинки, напитавшиеся самки и яйца. Состояние диапаузы, по-видимому, не возникает, так как при достаточно теплой погоде в отдельные зимние дни наблюдалось нападение на скот активизировавшихся личинок в Южном Таджикистане. Как правило, зимой скот свободен от клещей. Массовое нападение личинок начинается с марта, и в течение теплого сезона на скоте наблюдаются весенняя, летняя и осенняя волны заклещевления (рис. 41). Последние, по-видимому, отражают смену 3 частично перекрывающихся поколений В. annulatus, успевающих развиться за 1 год (Потоцкий, 1945; Галузо, 1950). Для В. decoloratus, являющегося эндемичным паразитом диких полорогих и крупного рогатого скота в Южной Африке, в условиях местного субтропического климата за 1 год успевает развиться от 3 до 4 генераций.
Им соответствуют 4 пика численности этих клещей на скоте в июле, сентябре—октябре, декабре—январе и марте—апреле (Rechav, Kostrezew- ski, 1991).Hyalomma
Для видов этого рода описаны в основном 1-годичные циклы, которые сдхра- няются даже у видов, обитающих в условиях пустынь Средней Азии и Казахстана с достаточно суровой и продолжительной зимой. В лабораторных культурах при температуре 25 °С средние сроки развития у 5 видов этого рода варьировали от 98 до 196 сут (Ouhelli, 1994).
Н. asiaticum широко распространен в пустынных и полупустынных ландшафтах от Сирии на западе до Монголии и Центрального Китая на востоке, от Казахстана на севере до побережья Персидского залива на юге. Это треххозяинный вид, все фазы развития которого в пустынях Туркмении встречаются круглогодично. Массовое паразитирование имаго на скоте и верблюдах наблюдается в апреле—мае и сентябре—октябре. Личинки паразитируют на больших песчанках и других мелких и средних млекопитающих с марта по сентябрь с максимальной численностью в июне—июле, нимфы — на тех же хозяевах с февраля по ноябрь с максимумом в августе—сентябре. Напитавшиеся весной самки в условиях высоких температур без задержки приступают к яйцекладке, а вылупившиеся из яиц личинки дают пик численности на хозяевах в июне—июле. С перелинявшими на них нимфами связан позднелетний пик численности на песчанках. На зимовку уходят голодные и напитавшиеся имаго и нимфы и голодные личинки от позднелетних кладок. Значительная индивидуальная долговечность голодных особей, достигающая в условиях микроклимата нор большой песчанки нескольких месяцев для личинок и нимф и более 1 года для имаго, приводит к частичному смешиванию потомства от самок весеннего и осеннего питания и даже клещей двух последовательных генераций. Таким образом, при преимущественно 1-годичном цикле часть популяции, представленная вылупившимися осенью личинками, уже принадлежит второй генерации клещей (рис. 42). Развитие ее завершается уже после зимовки в следующем сезоне.
У нимф же и особенно имаго, не сумевших в теплый сезон найти хозяина и ушедших на зимовку, развитие затягивается до 1.5—2 лет (Балашов, 1972; Бердыев, 1980).Н. dromedarii широко распространен в пустынных районах Африки к северу от экватора, в странах Ближнего и Среднего Востока, Средней Азии. Главными прокормителями для всех фаз служат верблюды и крупный рогатый скот. В южных частях ареала развитие идет преимущественно по треххозяинному типу, а на севере
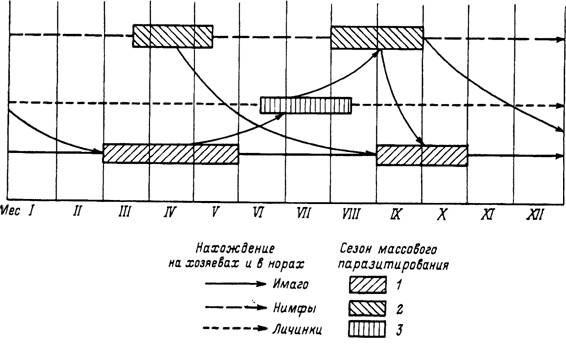
Рис. 42. Цикл развития клеща Hyalomma asiaticum иа юге Туркмении.
1 — имаго; 2 — иимфы; 3 — личинки.
ареала в Туркмении в летние месяцы — по двух- и треххозяинному типам, а в зимние — по однохозяинному. В Туркмении взрослые клещи встречаются на верблюдах круглый год, но в максимальном количестве — в июне—августе и в ноябре (рис. 43). Нимфы также паразитируют на верблюдах круглогодично с максимумами в августе и ноябре (рис. 44), причем напитавшиеся в конце осени особи остаются зимовать на теле хозяина и отпадают только весной. Пики численности личинок на хозяевах наблюдаются в июне—июле и осенью (рис. 44). Напитавшиеся осенью самки впадают в зимнюю диапаузу, зимуют в почве и откладывают яйца весной. При треххозяинном типе развития для завершения 1 поколения в теплое время года требуется 103—127 сут, а при двух- и однохозяинном развитии эти сроки еще короче. Соответственно, с учетом зимней остановки, в течение 1 года успевает развиться до 1.5 поколения и более, а границы между последовательными поколениями стерты (Бердыев, 1969, 1980).
Ареал Н. anatolicum охватывает пустынные и степные ландшафты Средиземноморья и Северной Африки почти до экватора, Аравийский полуостров, Иран, Пакистан, Индию и Среднюю Азию. Клещи при питании всех фаз на крупном
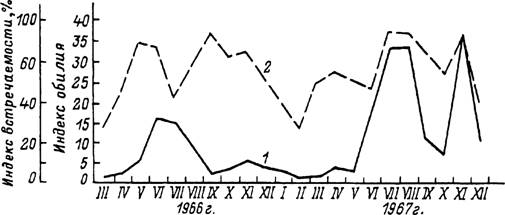
Рис. 43. Паразитирование имаго клеща Hyalomma dromedarii иа верблюдах в предгорных районах Туркмении в 1966—1967 гг.
(по: Бердыев, 1969).J — индекс обилия; 2 — индекс встречаемости.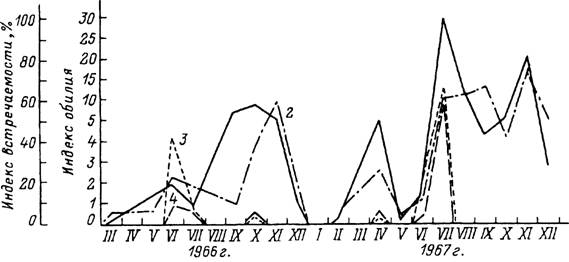
Рис. 44. Паразитирование личинок и нимф клеща Hyalomma dromedarii на верблюдах в предгорных районах Туркмении в 1966—1967 гг. (по: Бердыев, 1969).
J — индекс обилия иимф; 2 — индекс встречаемости иимф; 3 — индекс обилия личинок; 4 — индекс встречаемости личинок.
рогатом скоте имеют преимущественно двуххозяинный цикл, а при питании личинок на мелких млекопитающих — треххозяинный. На юге Туркмении имаго паразитируют на скоте круглогодично. Численность их быстро возрастает с середины апреля и достигает максимума в июне—августе, затем постепенно снижается, так что с ноября по март на животных встречаются единичные особи (преимущественно самцы). Личинки паразитируют с мая до конца сентября с максимумом в августе, нимфы — с мая по октябрь с максимумом в июле. Зимуют голодные и напитавшиеся имаго и нимфы и, возможно, в небольшом количестве личинки. Перезимовавшие и активизировавшиеся весной имаго сразу после питания приступаю^ к яйцекладке одновременно с перезимовавшими самками осеннего насыщения. Вылупившиеся из весенних кладок личинки успевают к концу осени развиться до стадии голодных или напитавшихся нимф, которые уходят на зимовку. Перезимовавшие сытые и голодные нимфы уже в конце мая—июне линяют в имаго, которые успевают напитаться или уходят на зимовку голодными. В результате продолжительность развития 1 поколения занимает от 1 до 1.5—2 лет (Бердыев, 1980).
Сходный 1-годичный цикл развития описан у этого вида и в Израиле (Rubina et al., 1982). В условиях эксперимента показано (Белозеров, 1981), что под влиянием длины дня может возникать состояние диапаузы. Она проявляется в задержке развития напитавшихся нимф, оогенеза самок и развития яиц, а также в снижении агрессивности и увеличении срока выживания непитавшихся особей. В условиях Туркмении диапауза возникает с начала сентября при длине дня в 13 ч и менее, а с середины сентября она наблюдается у 100 % напитавшихся в природе нимф.
В Египте формирование диапаузы этого вида происходит позднее, когда среднесуточные температуры опускаются ниже 25 °С. У напитавшихся самок диапауза возникает при длине дня в 14 ч и меньше, и соответственно в условиях Средней Азии задержка откладки яиц наблюдается у особей, напитавшихся в конце августа или начале сентября. В Египте же уход самок на зимовку зависит главным образом от температуры, и диапауза может возникнуть не ранее конца сентября—начала октября. Существование стадии морфогенетической диапаузы у нимф и самок Н. anatolicum обеспечивает сезонную упорядоченность цикла развития этого вида за счет совпадения сроков линьки нимф или яйцекладки самок независимо от осеннего или весеннего периодов их питания.Н. detritum имеет сходное с предыдущим видом распространение, но заселяет более мезофильные местообитания и тяготеет к орошаемой зоне, долинам рек.Цикл развития двуххозяинный, и все стадии питаются преимущественно на крупном рогатом скоте. При облигатном моновольтинном цикле развития за год развивается только 1 поколение клещей, даже если сумма эффективных температур достаточна для частичного развития второй генерации. Диапауза в этих случаях не только обеспечивает перезимовку клещей, но и синхронизирует их жизнедеятельность с определенными фенологическими сезонами. В условиях Южного Казахстана и в Средней Азии имаго паразитируют на скоте в июне—августе. Напитавшиеся самки через 1—2 нед приступают к яйцекладке, и из яиц без задержки вылупляются личинки. Последние остаются скоплениями в местах кладок в неактивном состоянии до конца сентября—октября. С начала октября начинается нападение личинок на скот, и напитавшиеся нимфы отпадают в октябре—ноябре. Часть нимф остается на теле хозяев и отпадает с началом весны в марте. Независимо от сроков отпадения линька нимф происходит в конце весны, и свеже- перелинявшие имаго начинают без задержки нападать на скот (Галузо, 1947; Бердыев, 1980).
Н. scupense сходен в своем распространении с Н. detritum, но заходит дальше на север, встречаясь на юге Украины и России. Развитие идет только по одно- хозяинному циклу. Личинки нападают на скот не ранее конца сентября. Напитавшиеся личинки остаются на теле хозяина и линяют в нимф, без задержки продолжающих питание. Имаго, перелинявшие из нимф в декабре—феврале, остаются прикрепившимися к хозяину, но не питаются, по-видимому находясь в состоянии диапаузы. Первые напитавшиеся самки в условиях Южного Казахстана появляются с конца марта, и отпадение их продолжается в апреле—начале мая. Они без задержки откладывают яйца, из которых в конце весны—начале лета вылупляются личинки. Последние весь летний период остаются в почве и растительных остатках в неактивном состоянии. Активные личинки поднимаются на растительность и нападают на скот только с конца сентября (Галузо, 1947).
Н. marginatum обладает исключительно обширным ареалом, включающим Средиземноморье, Среднюю Азию, большую часть Африки и Индостанский полуостров. Клещи встречаются в различного типа степях и саваннах, включая аридные горные редколесья. Цикл развития двуххозяинный. Неполовозрелые фазы паразитируют на питающихся на земле птицах, зайцах, ежах и других млекопитающих, а взрослые клещи — на крупном рогатом скоте, лошадях и других домашних и диких животных с крупными и средними размерами тела. В Средней Азии, Закавказье и на юге России цикл развития 1-годичный моновольтинный. Зимующими стадиями являются голодные имаго и напитавшиеся нимфы. Активация перезимовавших или перелинявших весной имаго и нападение их на млекопитающих происходят весной и в первой половине лета. Напитавшиеся самки без задержки откладывают яйца, из которых через 20—40 сут вылупляются и сразу же активизируются личинки. Сезон паразитирования их ограничен второй половиной лета и осенью (Галузо, 1947; Березин, 1971; Бердыев, 1980).
Н. aegyptium, паразитирующий на всех фазах преимущественно на наземных черепахах в Средиземноморье, Закавказье, Иране и Средней Азии, обладает трех- хозяинным 1-годичным циклом развития. Весной на активизировавшихся после зимней спячки черепахах последовательно сменяются питающиеся личинки, нимфы и в начале лета — имаго.
Rhipicephalus
Для большинства видов этого рода характерны треххозяинные циклы развития. Только у трех видов (R. evertsi, R. bursa, R. glabroscutatum), объединяемых в подроде Digineus, развитие идет по двуххозяинному типу.
R. bursa широко распространен в странах Средиземноморья и юга Средней Азии до Синьцзяна на востоке. Клещи обитают в аридных степных, кустарниковых и лесных стациях в условиях достаточно жаркого и продолжительного лета. Главными прокормителями для всех фаз служат овцы, козы, крупный рогатый скот и реже дикие копытные и зайцы. В условиях континентального климата Южного Дагестана с достаточно суровой зимой личинки появляются на хозяевах с середины лета, и паразитирование неполовозрелых особей продолжается до конца октября. Зимуют перелинявшие голодные имаго и напитавшиеся нимфы. С начала весны численность питающихся на скоте самок быстро увеличивается и достигает максимума в мае—июне (Ганиев, 1958). В условиях Израиля неполовозрелые фазы паразитируют с ноября по март, а имаго — с апреля по июль (Yeruham et al., 1989). По-видимому, этому виду свойствен моновольтинный 1-годичный цикл развития с осенне-зимне-весенним паразитированием личинок и нимф в условиях теплой зимы и летне-осенним паразитированием и прекращением зимней активности в условиях низких температур.
Ареал R. evertsi охватывает аридные ландшафты Африки к югу от Сахары. Все фазы развития паразитируют на скоте, овцах и диких копытных, а неполовозрелые фазы также часто встречается на зайцах. В ЮАР клещи встречаются на скоте круглогодично, но с четко выраженными сезонными пиками. Весной наблюдается быстрый подъем численности питающихся имаго, вероятно за счет большого числа перезимовавших напитавшихся нимф. Питающиеся самки многочисленны до конца осени, так что сезон их паразитирования продолжается с января до конца мая. Неполовозрелые фазы развиваются на хозяине от 13 до 29 сут (в среднем 19) и наиболее многочисленны с января по апрель. Предполагают, что за год может развиться 2 и более поколений (Londt, Bijl, 1977).
R. sanguineus наиболее известен среди треххозяинных видов этого рода благодаря всесветному распространению в странах с тропическим, субтропическим и даже умеренным климатом. Первоначально этот вид обитал в Африке, где паразитировал на местных видах диких хищников и копытных (Hoogstraal, 1978). Вместе с собаками, ставшими его главными и часто единственными хозяевами, он распространился по другим континентам, стал синантропом и приспособился к развитию внутри населенных пунктов и помещений для содержания собак. R. sanguineus обладает поливольтинным типом развития и в течение 1 года может развиться несколько поколений. В странах Средиземноморья за год успевают развиться 2 поколения, которым соответствуют 2 пика заклещевления собак. Зимуют главным образом напитавшиеся нимфы и голодные имаго (Feldman-Muhsam, 1981). В странах тропической Африки и Сингапуре цикл развития становится непрерывным и за 1 год развивается 3 и более поколений (Pegram, 1984). Об исключительной экологической пластичности этого вида свидетельствует появление его локальных популяций в Германии (Gothe, Hammel, 1973) и Польше (Szymanski, 1979), где эти клещи успешно размножаются в теплых помещениях для собак.
R. turanicus широко распространен в аридных ландшафтах Африки, Средиземноморья, Средней Азии и заходит на восток до Синьцзяна. Взрослые клещи паразитируют преимущественно на овцах, козах и на крупном рогатом скоте, на крупных и средних диких животных. Личинки и нимфы питаются на грызунах и зайцеобразных. В условиях Африки взрослые клещи паразитируют в конце дождливого и начале засушливого сезонов, а неполовозрелые фазы — в засушливый сезон (Pegram, 1984), имея 1-годичный цикл развития. В Туркмении имаго питаются на скоте и овцах с февраля по август с максимумом численности в апреле-июне (рис. 45). Напитавшиеся самки без задержки откладывают яйца, и из них уже в июне вылупляются и сразу же активизируются личинки. Неполовозрелые клещи встречаются на хозяевах в течение всего лета. Перелинявшие в конце лета и осенью имаго остаются в неактивном состоянии всю зиму до наступления весеннего потепления. В меньшем количестве зимуют напитавшиеся нимфы. Весь цикл развития укладывается в 1 год (Бердыев, 1980).
Отсутствие активации имаго осенью связано с развитием у них диапаузы при длине дня в 14 ч и менее. Подобная длиннодневная фотопериодическая реакция
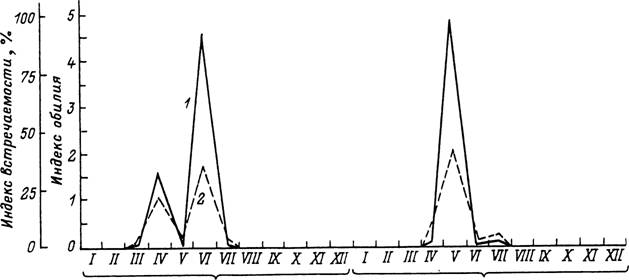
Рис. 45. Сезон паразитирования имаго клеїла Rhipicephalus turanicus в предгорных районах Туркмении в
1966—1977 гг. (по: Бердыев, 1980).
1 — индекс обилия: 2 — индекс встречаемости.
обеспечивает уход на зимовку всех голодных имаго от летне-осенней линьки текущего года и питание их вместе с имаго, перелинявшими от перезимовавших напитавшихся нимф (Белозеров, 1988).
R. ритіііо распространен на юго-западе России, в Казахстане, государствах Средней Азии до Синьцзяна и Западной Монголии. Клещи обитают в полупустынных и степных ландшафтах, где тяготеют к поймам рек и другим мезофильным стациям. Взрослые паразитируют на зайцах, ежах и сельскохозяйственных животных, а неполовозрелые — на зайцах, ежах, грызунах и реже — на птицах. Этому виду свойствен 2- и даже 3-годичный цикл развития. Питающиеся особи всех фаз развития встречаются на протяжении всего теплого сезона, а зимующими стадиями являются голодные имаго, нимфы и реже личинки (Галузо, 1948; Гребенюк, 1966). Активность голодных клещей регулируется фотопериодической реакцией длиннодневного типа и тормозится при укорочении светового периода до 15 ч и меньше, что обеспечивает перезимовку особей, перелинявших со второй половины лета (Белозеров, 1985).
R. schulzei относится к числу немногих видов этого рода, все развитие которых проходит внутри норы хозяина. Главными хозяевами являются несколько видов сусликов рода Citellus, а также степные хорьки (Mustela eversmanni) и в меньшей степени другие виды хищных и грызунов, использующие норы сусликов. Ареал вида связан с таковым сусликов и ограничен сухими степями, полупустынями и пустынями юга России, Казахстана и Средней Азии. Паразитирование клещей наблюдается с начала весны и до конца осени, причем одновременно на хозяевах встречаются личинки, нимфы и взрослые особи (рис. 46). Зимуют голодные неполовозрелые и взрослые особи, а также яйца. Весной на хозяевах преобладают перезимовавшие или свежевылупившиеся личинки и в меньшей степени нимфы и взрослые. Напитавшиеся весной личинки дают летний пик питающихся нимф. Имаго, перелинявшие из напитавшихся весной и летом нимф, также питаются в этом сезоне, причем часть из них также успевает напитаться и оставляет на зимовку яйца, а также успевших развиться от части из них личинок и реже нимф. От самок весеннего питания за 1 сезон могут успеть перелинять к осени голодные имаго следующего поколения, а немногие из них даже напитаться. В результате часть популяции развивается по 1-го- дичному циклу (Нельзина, Данилова, 1960), но ее ббльшая часть имеет 2-годичный цикл (Кондрашкина, 1951; Шатас, 1952; Левит, 1957).
R. appendiculatus принадлежит к числу наиболее полно изученных видов в связи с его исключительным значением как переносчика тейлериоза и лихорадки
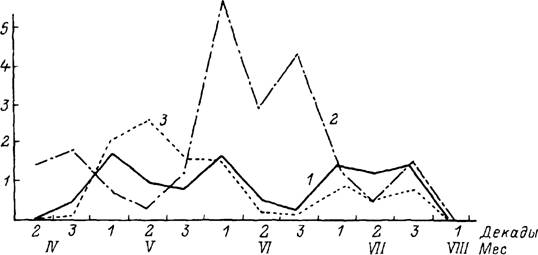
Рис. 46. Сезонная динамика паразитирования клещей Rhipicephalus schulzei на малых сусликах (по: Конд-
раішЛіна, 1951).
1 — имаго: 2 — нимфы; 3 — личинки. По вертикали — индекс обилия клещей.
Восточного Берега крупного рогатого скота. Вид широко распространен в Африке к югу от Сахары. Все фазы паразитируют на крупном рогатом скоте, а также на овцах, козах и антилопах. Сезонная активность значительно варьирует в зависимости от разных типов чередования дождливых и засушливых периодов в границах ареала. Активность взрослых клещей ограничена дождливыми сезонами и кроме влажности зависит от температуры и длины дня. В свою очередь сроки отпадения напитавшихся самок предопределяют активность личинок и нимф, приходящуюся на засушливый сезон. В зависимости от климатических условий за 1 год развивается от 1 поколения в ЮАР до 2 и, возможно, более поколений в странах Экваториальной Африки (Punyua, 1984; King et al., 1988).
Dermacentor
У клещей этого рода палеарктические виды характеризуются 1-годичными циклами развития, в сезонной регуляции которых главная роль принадлежит имаги- нальной диапаузе. Наиболее детально исследованы D. reticulatus (Олсуфьев, 1953; Daniel et al., 1980; Разумова, 1986) и D. marginatus (Покровская, 1951, 1953; Nosek, 1972; Gilot, Pautou, 1985), которые имеют во многом сходные циклы развития. D. reticulatus заселяет достаточно увлажненные луговые и кустарниковые биотопы в Европе и Западной Сибири, а ареал D. marginatus занимает зоны равнинных и горных степей. Повсеместно перезимовавшие голодные имаго этих видов активизируются со сходом снега ранней весной и нападают на скот, а также на средних и крупных диких млекопитающих до начала лета. В июне—июле клещи становятся неактивными и прекращают нападать на хозяев, хотя и М017Т оставаться на растительности. Нападение на скот возобновляется в августе—сентябре, но осенний пик активности значительно слабее весеннего (рис. 47, 48). У D. reticulatus в условиях Словакии и Польши весной в естественных местообитаниях этого вида самки приступали к откладке яиц в зависимости от температурных условий через 1—25 сут после насыщения, а личинки вылуплялись из яиц через 44—80 сут. Метаморфоз напитавшихся личинок занимал 12—22 сут и нимф — 25—60 сут. Голодные личинки живут не более 20—60 сут и нимфы — 35—70 сут (Daniel et al., 1980).
D. marginatus в условиях континентального и жаркого лета юга России обладает более сжатыми сроками развития. У самок весеннего поколения яйцекладка начинается через 9—12 сут, а вылупление личинок происходит через 12—15 сут.
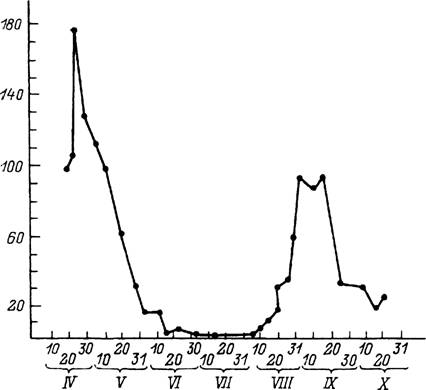
Рис. 47. Сезонный ход активности имаго Dermacentor reticulatus в Московской области (по: Разумова,
1986).
По вертикали — число клещей на 100 м; по горизонтали — даты сбора клещей с растительности.
Метаморфоз личинок занимает 3—7 сут и нимф — 22—26 сут. На все развитие от окончания питания самок до появления имаго следующего поколения требуется 90—ПО сут. Голодные личинки живут в природе не более 30—40 сут и нимфы — 40—50 сут (Покровская, 1951, 1953). Из-за эфемерности неполовозрелых форм зимовка обоих видов возможна только на стадиях голодных имаго и напитавшихся самок, а голодные или напитавшиеся, но не перелинявшие к зиме нимфы погибают. Напротив, голодные имаго отличаются исключительной долговечностью. У D.~reticulatus они могут жить в природе до 2.5 лет после линьки с 3 зимовками и 4 периодами активности, прерывающимися летом состоянием неактивности (рис. 49) (Разумова, 1966; Cerny et al., 1982).
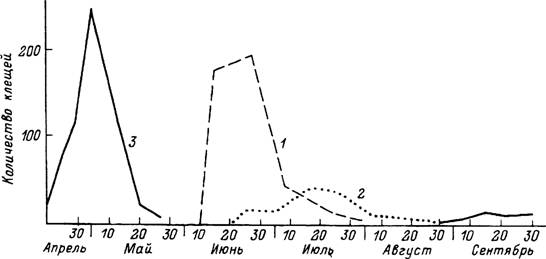
Рис. 48. Сезонность паразитирования клеща Dermacentor reticulatus в Московской области (по: Олсуфьев,
1953).
1 — средняя численность личинок на 5 обыкновенных полевках; 2 — то же для нимф; 3 — средняя численность имаго
на одной корове.
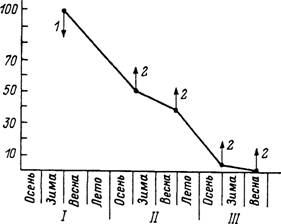
Рис. 49. Сроки жизни и повторная активность имаго Dermacentor reticulatus в природе в Московской области (по: Разумова, 1986).
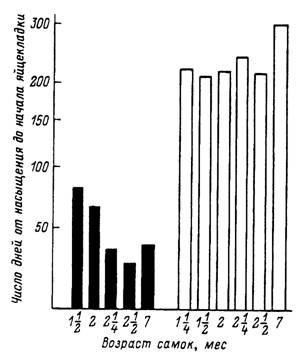
Рис. 50. Средние сроки от насыщения до начала яйцекладки (при 18 °С) у самок клеща Dermacentor marginatus из Волгоградской области в зависимости от возраста и фотопериодических условий содержания до кормления (по: Белозеров, 1965).
1 — выпуск окрашенных клещей; 2 — отлов клещей; 1—111 — последовательные годы жизни голодных клещей одного календарного возраста; по вертикали — число клещей (9Ь).
Черные столбики — 9 ч света; белые столбики — 18 ч света.
Самки, напитавшиеся с середины лета, и осенью находятся в состоянии репродуктивной диапаузы и приступают к яйцекладке только после зимовки вместе с особями весеннего насыщения. Поведенческая и репродуктивная диапаузы регулируются длиной светового дня. Для D. marginatus характерна короткодневная фотопериодическая реакция с критическим фотопериодом в 13 ч (рис. 50) (Белозеров, 1976). Экологическое значение обеих форм диапаузы однозначно и направлено на предотвращение откладки яиц в конце лета и осенью и гибели не достигнувших имагинальной фазы особей зимой. Во время зимовки низкие температуры обеспечивают к весне реактивацию всего запаса зимующих имаго.
У клещей D. silvarum (Беляева, Рябова, 1971), D. nuttalli (Teng Kuo-fan, 1962; Меринов, 1964) и D. sinicus (Teng Kuo-fan, 1981), обитающих в Восточной Палеар- ктике, включая Северный Китай и Монголию, в связи с жарким летом развитие протекает еще быстрее, чем у видов из Западной Палеарктики. Перезимовавшие голодные имаго питаются на скоте, крупных и средних диких млекопитающих в апреле—июне. Их потомство благодаря быстрому метаморфозу напитавшихся летом личинок и нимф уже к осени дает самцов и самок следующего поколения, так что развитие одной генерации без зимовки занимает не более 3—4 мес. Например, у D. nuttalli (Teng Kuo-fan, 1962) напитавшиеся самки в условиях, близких к естественным, приступают к яйцекладке через 3—25 сут по отпадении, и она продолжается 31—44 сут. Развитие яиц занимает 15—36 сут. Личинки способны к питанию уже через 3—4 сут после вылупления и линяют через 8—12 сут после питания, а нимфы линяют через 14—20 сут. Осенний период активности у имаго выражен слабо. Иногда немногие прикрепившиеся к хозяевам особи остаются ненапитавшимися на протяжении зимы и заканчивают питание следующей весной. Подобную задержку прикрепившихся к хозяину самок D. silvarum в связи с их неспособностью к насыщению В. Н. Белозеров (1973) считает особой формой диапаузы, регулируемой на основе короткодневной фотопериодической реакции. Для самок D. sinicus также было показано удлинение сроков питания после нахождения голодных особей в течение 2.5 мес в условиях длинного дня и укорочение их после "Пребывания в условиях короткого дня (Jiang Zai-jie et al., 1991).
У D. auratus, одного из немногих видов, обитающих в тропических лесах Юго-Восточной Азии и Индии, жизненный цикл по ряду черт сходен с таковым палеарктических видов (Jiang Zai-jie, 1987). Имаго этого вида паразитируют на циких свиньях в летне-осенние и зимние месяцы. Особи из Южного Китая, напитавшиеся в июне—августе, приступают к яйцекладке через 13—27 сут, а в ноябре — через 215—244 сут, так что этому тропическому виду также свойственна репродуктивная диапауза (Jiang Zai-jie et al., 1991). Развитие яиц у этого вида более медленное по сравнению с северными видами (вылупление личинок через 45—79 сут), а сроки жизни голодных нимф больше (до 124 сут). В связи с диапаузой за 1 год успевает развиться одна генерация, но сроки активности всех фаз развития более растянуты и налегают друг на друга.
Жизненные циклы американских видов значительно разнообразнее евроазиатских как по продолжительности, так и по числу хозяев, сменяемых в развитии. Наиболее детально изучен D. variabilis, широко распространенный в США. Клещи заселяют мезофильные листопадные леса, кустарниковые редколесья и залежные луга. Имаго паразитируют на средних и крупных диких млекопитающих, собаках и скоте, а личинки и нимфы — на мышевидных грызунах, кроликах и зайцах. На юге (Newhouse, 1983) цикл развития 1-годичный. Зимуют голодные имаго (весенняя когорта) и личинки. Клещи активизируются уже в марте. Пик паразитирования личинок в апреле, а перелинявших из них и сразу же активизировавшихся нимф — в конце мая—начале июня. Перелинявшие из этих нимф имаго уже в конце июня—начале июля дают летний пик активности (летняя когорта). Напитавшиеся самки без задержки откладывают яйца, из которых вылупляются личинки, перезимовывающие в состоянии диапаузы. Потомство напитавшихся самок весенней когорты развивается с отставанием от особей летней когорты и уходит на зимовку на стадии голодных имаго.
На севере США в Массачусетсе (McEnroe, 1982, 1983) D. variabilis не хватает 1 года для завершения цикла развития. Напитавшиеся весной самки весенней когорты откладывают яйца, из которых в том же сезоне успевают вылупиться и напитаться личинки и перелинять в зимующих нимф. Эти нимфы питаются следующей весной и линяют в начале лета в имаго летней когорты. Последние успевают в этом же сезоне напитаться, откладывают яйца и оставляют зимующих личинок. Напитавшиеся весной личинки успевают перелинять в нимф, которые в свою очередь питаются и линяют в имаго весенней когорты, оставаясь неактивными до следующей весны. Значительная индивидуальная долговечность голодных нимф и имаго обеспечивает регулярную панмиксию между клещами весенней и летней когорт. У северной границы ареала в Манитобе (Канада) цикл развития уже только 2-годичный (Burachynsky, Galloway, 1985).
Экспериментально установлено (Smith, Cole, 1941), что личинкам и нимфам этого вида свойственна длиннодневная фотопериодическая реакция. Клещи реагируют не на саму длину дня, но на его сезонные удлинения или укорочения. Короткий день способствует сохранению неактивного состояния у уходящих на зимовку голодных личинок и нимф, а удлинение дня вызывает не только активацию, но и поддержание состояния активного подстерегания хозяина весной и в начале лета.
Клещ D. andersoni широко распространен в горных лесах и прилегающих к ним прериях системы Кордильер от Канады до Мексики. В зависимости от географической широты местности, высоты над уровнем моря и микроклимата местообитаний развитие возможно по 1-, 2- и 3-годичному циклам (Wilkinson, 1968, 1979). Имаго повсеместно имеют единственный весенний период активности с марта—апреля до начала июня. При наиболее распространенном 2-годичном цикле перезимовавшие имаго питаются весной, и из отложенных ими яиц в этом же сезоне вылупляются личинки. Последние, по-видимому, находятся в состоянии диапаузы и, не активизировавшись, зимуют. Следующей весной они нападают на мелких млекопитающих и после питания вскоре линяют в нимф. Нимфы также успевают напитаться и перелинять в имаго, которые неактивны осенью и зимуют. В случае зимовки не успевших напитаться голодных нимф развитие занимает 3 года, а при вылуплении личинок, а также питании личинок и нимф в один и тот же сезон развитие завершается за 1 год. Перезимовавшие, но не нашедшие весной хозяина имаго становятся неактивными, зимуют и вновь активизируются следующей весной.
Состояние активности у голодных особей D. andersoni стимулируется предварительной реактивацией низкими температурами и угнетается при воздействии температур более 25 °С. У «прерийной» расы этого вида (Pound, George, 1991) в условиях сокращающейся длины дня сроки развития напитавшихся нимф значительно удлиняются по сравнению с таковыми в условиях постоянного или удлиняющегося фотопериода. У напитавшихся самок из сокращающегося или постоянного короткого дня масса тела была больше, чем у особей из удлиняющегося или постоянного длинного дня. Сроки развития напитавшихся личинок достоверно не менялись при разных режимах освещенности. Для особой «горной» расы влияния длины дня на развитие не выявлено.
D. albipictus с однохозяинным развитием имеет 1-годичный цикл. Ареал этого вида охватывает Северную Америку от Канады до Мексики. Клещи паразитируют на лосях, белохвостом олене и в меньшей степени на других диких копытных, а также на лошадях и скоте. Личинки нападают на хозяев осенью. В Канаде (Альберта) в северной части видового ареала активные личинки появляются на растительности с начала сентября, их численность быстро нарастает до начала октября и затем постепенно сокращается до полного исчезновения в декабре (Drew, Samuel,
1985) . Клещи встречаются на лосях осенью, зимой и весной, и минимальный период от прикрепления личинок до отпадения напитавшихся самок составляет 155 сут. Прикрепившиеся личинки сразу же начинают питаться и уже через 10— 22 сут появляются свежеперелинявшие нимфы. Последние остаются прикрепившимися, но не питаются в течение 3 мес до января, находясь в состоянии диапаузы. После этого они нормально питаются и линяют в имаго, которые также без задержки приступают к кровососанию. Количество имаго на лосях увеличивается с середины января до марта—апреля (Addison, McLaughlin, 1988).
Период отпадения с лосей напитавшихся самок приходится в Альберте на март—май. Яйцекладка начинается через 36—41 сут и занимает период с конца мая по конец июня. Личинки вылупляются через 76—106 сут (Drew, Samuel,
1986) . После вылупления личинки остаются в скоплении в неактивном состоянии (поведенческая диапауза), которое в зависимости от географической широты местности продолжается от 4—7 мес в Калифорнии до 2—3 мес в Британской Колумбии и всего около 2 нед в Альберте (Drew, Samuel, 1985). Диапауза контролируется фотопериодической реакцией короткодневного типа, которая снимается осенью при сокращении светового дня до 12 ч (Wright, 1971).
Anocentor
Единственный вид этого рода A. nitens ограничен в своем распространении влажными тропическими и субтропическими районами Южной и Центральной Америки. Однохозяинный вид, паразитирующий внутри ушных раковин и ноздрей лошадей, крупного рогатого скота и значительно реже диких животных. От прикрепления личинок до отпадения напитавшихся самок проходит 20—35 сут. Яйцекладка при 30 °С начинается через 3.1—3.4 сут по отпадении и продолжается 14.3—14.7, а при 20 °С — соответственно 8.5—9.8 и 28.9—30.2 сут. Минимальное время эмбрионального развития при 30 °С составляет 19.9 сут. Какие-либо формы задержки развития или диапаузы не обнаружены. Развитие 1 поколения продолжается 2—3 мес и за 1 год развивается несколько поколений (Rodriguez, Jimenez, 1989; Despins, 1992).
Amblyomma
Видам этого рода свойствен треххозяинный тип развития. Несмотря на преимущественное распространение в тропиках и субтропиках, поливольтинные циклы с развитием нескольких поколений за один год редки. Преобладают 1-годичные циклы, а у части видов даже 2- и 3-годичные. Невозможность завершения развития за 1 год ни в коей мере не связана с недостатком тепла, хотя развитие напитавшихся особей и особенно эмбриогенез протекают медленнее, чем у представителей других родов. В саваннах Африки, где многочисленны такие виды, как A. hebraeum и A. variegatum, перерывы в развитии связаны с сезонностью в выпадении осадков, чрезмерно высокими температурами и низкой относительной влажностью воздуха в жаркие и сухие сезоны.
Наиболее детально изучено развитие A. hebraeum. Этот вид широко распространен в Южной Африке в саваннах. Имаго и неполовозрелые фазы являются обычными паразитами крупного рогатого скота, овец, коз и других сельскохозяйственных животных. Из диких животных все фазы паразитируют на многих видах травоядных, а неполовозрелые клещи — также на птицах и рептилиях. Цикл развития в зависимости от климатических условий варьирует от 1- до 2-годичного. Взрослые активны преимущественно в летние месяцы, личинки — в конце лета, осенью и в начале весны, нимфы — зимой. Активность имаго приурочена к сезону выпадения дождей. В прибрежных районах с более мягким и влажным климатом все фазы развития встречаются одновременно и круглогодично, но также сохраняются сезонные пики активности. В эксперименте для этого вида выявлены положительная корреляция между пораженностью скота взрослыми клещами, длиной дня и температурой, а также отрицательная корреляция между указанными факторами окружающей среды и пораженностью скота нимфами. Предполагают, что активность имаго стимулируется в условиях длинного дня, а нимф — в условиях короткого дня (Petney et al., 1987).
Клещ A. variegatum широко распространен в Африке к югу от Сахары и вплоть до севера ЮАР и встречается в разнообразных ландшафтах от полуаридных саванн до влажных горных лесов. Круг хозяев тот же, что и у A. hebraeum. Имаго паразитируют в наибольшем количестве в дождливый сезон, а личинки и нимфы — с конца дождливого сезона и в засушливый период. В районах с 1 дождливым сезоном за год развивается 1 генерация, а при 2 сезонах дождей — возможно развитие 2—3 поколений (Petney et al., 1987). В Южной Африке обнаружена диапауза у напитавшихся самок. Она индуцируется при нахождении голодных самок в условиях короткого дня и заканчивается при понижении температуры с началом дождливого сезона. В результате у самок, напитавшихся в начале сезона активности, возникает задержка в развитии яиц, и они приступают к яйцекладке вместе с особями от более поздних сроков питания (Pegram et al., 1988).
A. marmoreum на имагинальной фазе паразитирует почти исключительно на черепахах и крупных ящерицах, а личинки и нимфы кроме рептилий питаются также на многих видах млекопитающих и птицах. Видовой ареал ограничен саванными редколесьями ЮАР, юга Мозамбика и Зимбабве. Сезон активности клещей тесно связан с таковым их главного хозяина — черепахи Geochelone pardalis. Для черепах характерен период зимней спячки с апреля—июня до сентября—октября, когда животные находятся в норах и не питаются. Клещи встречаются на черепахах на протяжении всего периода их активности с весны и до осени. Личинки появляются на черепахах с их выходом из спячки, и численность их достигает пика в апреле. Часть из них при этом остается прикрепившийся к телу хозяина, зимует на нем и отпадает следующей весной. Пик численности нимф приходится на весну, и затем количество их быстро снижается до минимума в конце лета—начале осени. Численность питающихся самцов и самок достигает максимума к середине лета и снижается к осени. Развитие 1 генерации завершается за 1 год только от нимф, напитавшихся весной. При этом перелинявшие из них самки должны успеть в это же лето напитаться, отложить яйца и дать активных личинок. Последние также должны успеть напитаться осенью и перелинять в нимф в осенне-зимний период. Часть популяции, развитие которой не укладывается в приведенные выше сроки, имеет 2-годичный цикл (Norval, 1975b).
А. атегісапит принадлежит к числу немногих видов, приспособившихся к условиям умеренного климата. Клещи распространены в юго-восточных и центральных штатах США и на севере Мексики, где обитают в разных типах лесов, в кустарниковых зарослях, залежных землях и на пастбищах. Имаго паразитируют на оленях и других крупных и средних диких млекопитающих, включая домашних животных, неполовозрелые — на тех же хозяевах, а также на птицах и грызунах. Сроки активности клещей варьируют в зависимости от климата, но их общей особенностью является прекращение паразитирования имаго с середины лета, а нимф — с начала осени. Имаго, не сумевшие найти хозяина к середине лета, или добавившиеся к ним свежеперелинявшие особи могут оставаться на поверхности почвы или на растительности, но у них отсутствует комплекс подстерегающего поведения и они не реагируют на присутствие хозяина. У нимф отмечены 2 пика численности на хозяевах, из которых весенний обеспечивают перезимовавшие особи, а позднелетний — нимфы, перелинявшие в этом же сезоне, и в меньшей степени сохранившиеся с весны. Зимуют голодные нимфы и имаго, отличающиеся большой долговечностью и устойчивостью к неблагоприятным внешним условиям (Sonenshine, Levy, 1971; Robertson et al., 1975; Barnard, 1988).
Развитие 1 генерации требует около 2 лет, но у части популяции может занимать и 3 года. Синхронизация в развитии клещей одного поколения обеспечивается диапаузой нимф и имаго (Haile, Mount, 1987). В эксперименте было показано, что при продолжительности фотопериода в 12 ч прикреплялось и напитывалось 98 % нимф и 100 % самок, а при длине в 8 ч — 62 % нимф и 51 % самок. Нападение голодных клещей стимулировалось не только общей продолжительностью, но и процессом увеличения длины дня и тормозилось при его укорачивании. Длина фотопериода и температура влияли не только на способность клещей к прикреплению, но также на массу поглощенной пищи и сроки питания, а у напитавшихся нимф также и на сроки их подготовки к линьке (Barnard et al., 1985; Pound, George, 1988).
Haemaphysalis
Несмотря на большое видовое разнообразие, жизненные циклы изучены только для немногих видов. Всем им свойствен треххозяинный тип развития продолжительностью от 1 до 4 лет.
Наиболее детально изучено развитие Н. longicornis, обитающего в Юго-Восточной Азии в странах с субтропическим и умеренным климатом, включая и юг Приморского края в России. В связи с обширным ареалом эти клещи встречаются в разных типах лесов и на травянистых пастбищах при условии их достаточной увлажненности. Взрослые паразитируют на многих видах травоядных и хищников, особенно на оленях и на скоте. Неполовозрелые питаются на мелких млекопитающих, козах и на птицах. В зависимости от климата цикл развития может быть 1-годичным на юге и 2—3-годичным на севере. В Австралии (Sutherst, Bourne, 1991) в более прохладных горных лесах зимуют диапаузирующие голодные взрослые и неполовозрелые клещи; жизненный цикл клеща хорошо синхронизирован со сменой сезона года и завершается за 1 год. В более жарких прибрежных местообитаниях у части популяции осенне-зимняя диапауза нарушается, и развитие может быть более коротким. В Японии и Южной Корее с более холодной зимой цикл развития 2-годичный с зимовкой голодных имаго и нимф (Ito et al., 1983). В Приморском крае у северной границы ареала (Беликова, 1969) цикл развития становится 2- и 3-годичным с зимовкой голодных личинок, нимф и взрослых.
У партеногенетической расы Н. longicornis голодные личинки и нимфы при 18 °С были активны -в условиях длинного дня (14—24 ч света). При освещенности менее 13 ч у личинок и 15 ч у нимф у клещей возникала и поддерживалась свыше 4—5 мес поведенческая диапауза в форме отсутствия агрессивности. Перенос клещей в условия длинного дня вызывал в течение 1—3 нед активацию, и клещи начинали нападать на прокормителей. У напитавшихся личинок фотопери- одические условия также регулируют продолжительность метаморфоза, так что в коротком дне развитие шло быстрее, чем в длинном (Белозеров, Лузев, 1974). Фотопериодическая индукция диапаузы голодных особей обнаружена в природе в Австралии, где в условиях укорачивающегося дня голодные нимфы и имаго становились неактивными (Sutherst, Bourne, 1991). В Японии нахождение голодных нимф и имаго этого вида в условиях короткого дня также вызывало развитие поведенческой диапаузы, а развивающиеся яйца, голодные и напитавшиеся личинки не реагировали на длину дня (Fujimoto, 1995а).
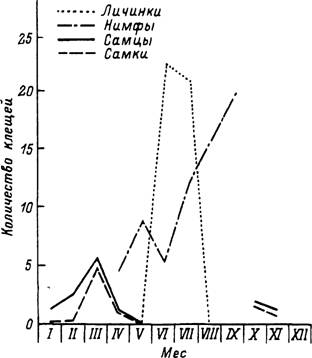
Н. sulcata распространен в степных и полупустынных ландшафтах Средиземноморья и Средней Азии. Взрослые паразитируют главным образом на козах и овцах. Неполовозрелые питаются на многих видах рептилий, реже на птицах и мелких млекопитающих. В Израиле (Feldman-Muhsam, Borut, 1983b) в условиях мягкой зимы взрослые паразитируют без перерыва с ноября по апрель. Самки, закончившие питание в начале зимы, приступают к яйцекладке в марте—апреле вместе с особями от более поздних сроков отпадения. Личинки появляются в мае и встречаются на рептилиях до августа, часто вместе с нимфами. Напитавшиеся личинки вскоре линяют в нимф, и последние питаются с конца июня до сентября. Напитавшиеся нимфы успевают перелинять в имаго осенью и сразу же начинают нападать на хозяев. В результате развитие 1 генерации укладывается в 1 год. В Киргизии (Гребенюк, 1966) с достаточно суровой зимой цикл развития занимает
1.5— 2 года. Взрослые паразитируют на скоте преимущественно осенью и ранней весной с зимним перерывом. Неполовозрелые встречаются на рептилиях весь летний период (рис. 51). Напитавшиеся личинки успевают в этом же теплом сезоне перелинять в нимф, но только часть из последних успевает напитаться до наступления осенних холодов. Значительная часть голодных и все напитавшиеся нимфы зимуют и завершают развитие следующим летом. Перелинявшие из них имаго зимуют напитавшимися или голодными и откладывают яйца только после зимовки.
Н. punctata также принадлежит к числу широко распространенных видов и ареал его сходен с таковым Н. sulcata, но заходит дальше на север, включая некоторые острова Северного и Балтийского морей. Вид обладает высокой экологической пластичностью и встречается в разных типах равнинных и горных ландшафтов со степной, кустарниковой и разреженной лесной растительностью. Список хозяев насчитывает несколько десятков видов млекопитающих, а для неполовозрелых фаз — также птиц и рептилий. Активность имаго в зависимости от географической широты местности имеет 2 пика, различающихся по продолжительности и обилию клещей, — весен-
Рис. 51. Сезонность паразитирования личинок и нимф клеща Haemaphysalis sulcata иа ящерицах и имаго иа скоте в Киргизии (по: Гребенюк, 1966).
ний и осенний. Неполовозрелые фазы активны с начала весны и до конца осени (Гребенюк, 1966; Nosek, 1973; Успенская, 1987). В Молдавии зимуют все фазы развития и продолжительность жизненного цикла сильно варьирует. В оптимальных условиях развитие 1 генерации завершается за 1.5 года. Потомство напитавшихся весной самок успевает в этом же сезоне достигнуть стадий голодных или напитавшихся нимф, которые после зимовки уже к середине следующего лета достигают имагинальной фазы. Преобладает 2-годичный цикл, когда от весенних кладок яиц на зимовку уходят напитавшиеся или голодные личинки, которые только с середины следующего лета линяют в нимф. Последние зимуют голодными или напитавшимися и на следующее лето линяют в самцов и самок, способных к осеннему питанию. У части популяции клещей, не сумевших напитаться ранее второй половины осени, развитие 1 поколения может затягиваться до 3 и даже 4 лет. Во всех случаях при прочих равных условиях отмечено сокращение сроков эмбрионального развития и линек личинок и нимф в весенне-летний период и удлинение их в осенние месяцы.
Н. сопсіппа в своем распространении связан с поясом широколиственных лесов Палеарктики. Кроме разных типов мезбфильных лесов клещи встречаются на влажных лугах, в зарослях кустарника и камыша. Хозяевами служат многие виды диких млекопитающих, скот и птицы. Повсеместно на протяжении теплого времени года одновременно активны на растительности и паразитируют все фазы развития (рис. 52). Зимуют также все фазы, кроме напитавшихся самок и яиц. Развитие 1 генерации продолжается 3 года и более (Беликова, 1969; Nosek, 1973; Lebedeva, Korenberg, 1981). Синхронизация жизненного цикла в условиях больших климатических различий в границах ареала достигается поведенческой диапаузой голодных взрослых и неполовозрелых особей, а также морфогенетической диапаузой у напитавшихся личинок и нимф. У личинок короткий день обеспечивает длительное, свыше 3 мес сохранение их скоплений в местах вылупления. В условиях длинного дня личинки активизируются и расползаются от места кладки уже через 10 сут. У нимф и взрослых длина дня непосредственно контролирует их агрессивность. В эксперименте в условиях длинного дня к мышам присасывалось 41—58 % выпущенных на них нимф, а в коротком дне только 5—7 % (Белозеров, 1974).
Для Н. bispinosa, обитающего в тропическом климате Индийского субконтинента от Шри-Ланки до Гималаев, характерен 1-годичный цикл развития. Питание взрослых, яйцекладка, развитие личинок и нимф завершаются в сезон летних муссонных дождей, а сухой сезон с декабря до мая переживают неактивные голодные имаго (Bhat, 1971).
Высокой экологической пластичностью отличается Н. leporispalustris. Этот вид распространен от Аляски до Аргентины, где приурочен к местообитаниям своих главных прокормителей — зайцеобразных. Кроме кроликов и зайцев, на которых питаются все фазы развития, неполовозрелые в большом количестве паразитируют на многих видах птиц. В южных прибрежных районах США с мягким климатом цикл развития 1-годичный. Личинки в наибольшем количестве встречаются на хозяевах с июля по декабрь, нимфы — в августе—сентябре, а взрослые — весь
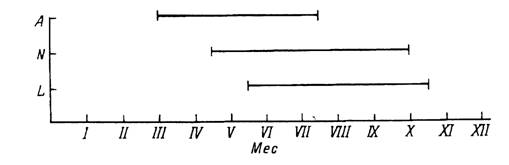
Рис. 52. Сезонность паразитирования клеща Haemaphysalis сопсіппа в Австрии (по: Sixl, 1973). L —личинки; N — нимфы; А — имаго.
год с максимумом в мае (Sonenshine, Stout, 1970). В районах с холодной зимой, как например в Миннесоте (Green et al., 1943), с декабря по март пораженность клещами их главного хозяина — зайца-беляка минимальна, так как зимующие голодные особи всех фаз развития находятся в неактивном состоянии. Количество питающихся клещей начинает увеличиваться только с апреля, и большое количество питающихся на всех фазах сохраняется с мая по июль. Быстрое снижение питающихся особей происходит в октябре—ноябре. В мае в наибольшем количестве паразитируют взрослые, а в августе — личинки. Как можно судить по сезонности активности неполовозрелых и взрослых особей, для развития 1 генерации требуется не менее 2 лет.
Ixodes
В наиболее богатом видами роде Ixodes при исключительно треххозяинном типе развития продолжительность существования каждой из фаз жизненного цикла и общая продолжительность лоследнего отличаются большим разнообразием.
В тропических и субтропических районах, по-видимому, достаточно широко распространены 1-годичные циклы. В частности, он свойствен обитателю лесов Восточной Австралии I. holocyclus. Активность личинок у него наблюдается летом- осенью, нимф — осенью-зимой и имаго — весной. У части популяции возможно удлинение развития до 2 лет (Doube, 1979). Сходный 1-годичный цикл существует и у /. rubicundus в Южной Африке (Fourie et al., 1989), причем у последнего вида активность ограничена достаточно влажными зимними месяцами. Как можно судить по последовательной смене периодов активности имаго, личинок и нимф в течение 1 года у I. matopi из Зимбабве, этот вид также должен иметь 1-годичный цикл развития, и он регулируется диапаузой у напитавшихся нимф (Colborne et al., 1981).
В умеренном климате 1-годичные циклы встречаются у обитателей нор и гнезд с достаточно теплым микроклиматом, но при этом часть популяций не успевает закончить развитие за теплый сезон и имеет 2-годичный цикл. /. lividus, паразит гнезд ласточек-береговушек (Riparia riparia), распространен по всей Палеарктике от Атлантического до Тихого океана. Быстрое развитие клеща в летние месяцы обеспечивается высокими температурами (30—32 °С) в гнезде хозяина в период насиживания яиц и развития птенцов. В Московской области (рис. 53) перезимовавшие в старых гнездах голодные личинки нападают во второй половине мая на прилетевших с юга взрослых птиц и питаются на них. Отпадение напитавшихся личинок происходит в конце мая—июне уже в новых гнездах. Нимфы появляются в конце июня—начале июля к моменту вылупления птенцов, сразу же нападают на них и отпадают внутри гнезда. Линька в имаго происходит в течение июля, и самки успевают напитаться на голых или уже оперившихся птенцах. Яйцекладка происходит с середины июля до начала августа. В конце августа—начале сентября вылупляются личинки, которые в состоянии диапаузы зимуют в оставленных птицами гнездах (Глащинская-Бабенко, 1956). В северной части ареала (Финляндия, Ленинградская область) небольшая часть популяции зимует на стадиях голодных нимф и имаго, и в этом случае цикл развития становится 2-годичным (Золотов, Букер, 1976; Ulmanen et al., 1977).
Сходным 1-годичным циклом, строго синхронизированным с гнездовым периодом хозяина, обладает клещ I. baergi — паразит скальной ласточки (Hirundo руг- rhonoto) в южных аридных районах США (George, 1987; Larimore, 1987). В середине мая после прилета и начала гнездования ласточек голодные перезимовавшие личинки перемещаются из трещин в скалах внутрь гнезд, где нападают и питаются на взрослых птицах. Напитавшиеся клещи отпадают с хозяина, линяют в нимф, и последние присасываются к уже вылупившимся из яиц птенцам. Напитавшиеся
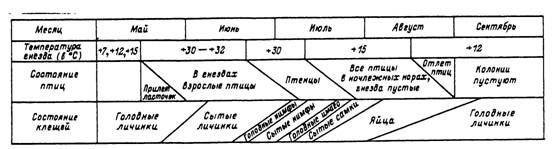
Рис. 53. Жизненные циклы Ixodes lividus и ласточек-береговушек в Московской области (по: Глащииская-
Бабеико, 1956).
нимфы выползают из гнезд, уходят в трещины скал и линяют в них во взрослых клещей. Голодные самки вновь перемещаются в гнезда и в первые две недели июля успевают напитаться на заканчивающих развитие птенцах. Отпавшие напитавшиеся самки выползают из гнезда, копулируют вблизи них с непитающимися самцами и уходят в трещины скал. В них происходит яйцекладка и вылупление личинок, которые остаются в неактивном состоянии до следующей весны. Как видно, в отличие от /. lividus у /. baergi развитие напитавшихся особей возможно вне гнезд ласточек благодаря высоким температурам юга США.
У клещей, обитающих в норах млекопитающих, благодаря возможностям контакта с хозяевами на протяжении всего года и более низким температурам внутри самой норы цикл развития менее жестко синхронизирован с таковым хозяина, и для его завершения может быть недостаточно 1 года. Так, у /. laguri — паразита сусликов рода Citellus, мелких хищников и других млекопитающих (свыше 30 видов), распространенного в степной зоне Евразии, все развитие протекает в норах хозяев. Зимуют голодные нимфы и имаго, которые активизируются весной и паразитируют на протяжении всего теплого сезона. Личинки от напитавшихся весной самок появляются в мае и тоже активны на протяжении лета—начала осени (Боженко, Шевченко, 1956). В условиях Чехословакии развитие одного поколения растянуто на 2 года (рис. 54), но часть популяции может развиваться и по 3-го- дичному циклу (Honzakova et al., 1980). В южных частях ареала возможен и 1-годичный цикл.
Сходным циклом развития обладает обитатель нор и гнезд околоводных млекопитающих /. apronophorusy распространенный по поймам рек и болот Европы и Южной Сибири. Главными прокормителями служат ондатра (Ondatra zibetica) и водяная полевка (Arvicola terrestris)y а всего более 40 видов млекопитающих. В Молдавии (Успенская, 1987) в плавнях реки Прут самки, нимфы и личинки паразитируют с марта до декабря с максимумами в апреле-мае и августе-сентябре. У личинок осенний пик активности значительно выше весеннего. Зимуют голодные
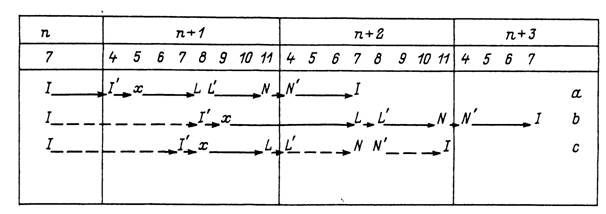
Рис. 54. Жизненный цикл Ixodes laguri в Чехословакии (по: Honzakova et al., 1980).
а — доказаный цикл; Ь, с — возможные циклы; L, N, I — голодные личинки, нимфы и имаго; L'% №, Г — напитавшиеся личинки, нимфы и самки; .г — кладка яиц; п ... л+і — календарные годы; 4, 5 ...11 — месяцы года; сплошная линия — доказанное развитие; прерывистая линия — возможное развитие.
особи всех фаз развития. Цикл развития завершается за 1 год, но чаще он 2-годичный, а при отсутствии прокормителей может быть и 3-годичным.
Клещ /. crenulatus распространен в степной зоне Евразии и обитает в постоянных норах более 30 видов животных, из которых главными хозяевами служат несколько видов сурков (Marmota), барсуки, лисы, хорьки. Клещи паразитируют даже на некоторых видах птиц, контактирующих с норами млекопитающих. В условиях холодного климата в горах Тянь-Шаня (Афанасьева, 1951) в норах сурка развитие одного поколения занимает не менее 3 лет. Одновременно летом в гнезде встречаются все фазы развития клеща с 2 пиками численности на хозяевах. Первый пик дают перезимовавшие нимфы и самки в марте, а второй — в июне — свежеперели- нявшие нимфы и личинки. В Молдавии (Успенская, 1987) в условиях более теплого климата клещи активны в норах барсуков, лис и других хищников на протяжении 10 мес. Пики численности питающихся самок приходятся на апрель, а нимфы многочисленны в течение всего теплого сезона с пиками в апреле и августе—октябре. Напитавшиеся весной личинки и нимфы успевают перелинять в этом же сезоне и напитаться в конце лета—начале осени. Личинки от летних кладок в основной массе остаются неактивными до следующей весны. По-видимому, для завершения развития 1 поколения в этих условиях требуется не менее 2 лет.
У видов с пастбищным типом паразитизма активность голодных особей и развитие напитавшихся протекает на растительности или на поверхности почвы. В результате эти клещи в большей степени зависят от местного климата и в первую очередь от гигротермических условий. В теплом умеренном климате для этих видов свойственны преимущественно 2-годичные циклы, но у части популяции наблюдается также 3-годичный цикл.
Преимущественно 2-годичным циклом обладает широко распространенный вдоль восточного побережья США /. scapularis (Yuval, Spielman, 1990). Личинки питаются на многих видах мелких млекопитающих и особенно на белоногом хомячке (Peromyscus leucopus) с июля по сентябрь. Перелинявшие из них нимфы активизируются после зимовки и также паразитируют на мелких млекопитающих с мая по июль. Перелинявшие в конце лета и осенью самки активизируются в этом же сезоне и большая часть популяции напитывается на белохвостом олене СOdocoileus virginianus) и других крупных и средних млекопитающих в осенние месяцы. При теплой погоде активность имаго продолжается зимой и заканчивается ранней весной следующего года. Существование осеннего и весеннего питания самок обусловливает разные сроки выплода и активации личинок и как следствие — наличие в популяции 2 частично обособленных когорт.
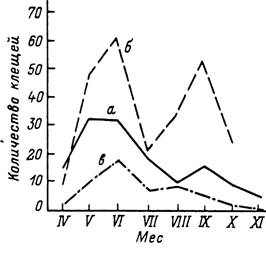
У некоторых видов, как например /. ricinus, жизненные циклы могут в зависимости от климата варьировать от 2- до 6-летних. В южных частях ареала, как например в горных лесах Алжира (Yousfi-Monod, Aeschlimann, 1986) или в Крыму (Мельникова, 1955), имаго активны в осенне-зимне-весенний сезоны непрерывно или с кратковременным прекращением во время сильных похолоданий. В умеренном климате Западной и Центральной Европы (Сегпу, 1957; Gigon, 1985; Gray, 1991; Craine et al., 1995), Белоруссии (Арзамасов, 1961) и средней полосы европейской части России (Бабенко, 1958) активность имаго приурочена к весенне-летне-осеннему сезону (рис. 55), причем весенний и осенний периоды обычно
Рис. 55. Сезонность паразитирования личинок и нимф клеша Ixodes ricinus иа мелких млекопитающих и имаго на скоте в Белоруссии (по: Арзамасов, 1961).
а — имаго; б — нимфы; в — личинки.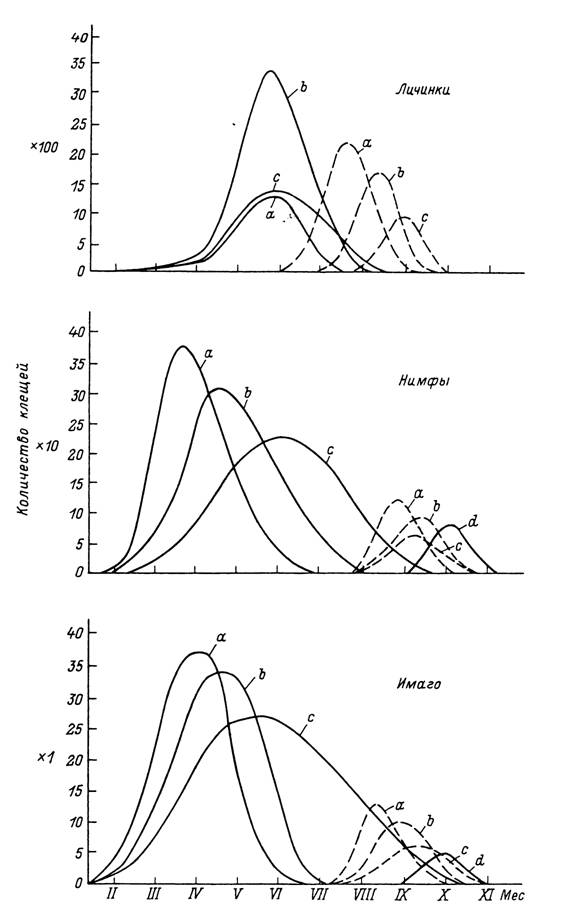
Рис. 56. Возможные типы сезонной активности клеща Ixodes ricinus в странах Западной Европы по данным сборов голодных клещей с растительности и питающихся хозяев (по: Gray, 1991).
а — луг; b — осветленный вторичный лиственный лес; с — сильно затененный лес; d — клещи, вылупившиеся весной, но активизировавшиеся осенью; сплошная линия — клеши весенней когорты; прерывистая линия — клещи осенней когорты.характеризуются максимумами численности, а летом может наступать снижение активности нападения клещей. В Карелии у северной границы ареала (Хейсин и др., 1955; Лутта, Шульман, 1958) при общем коротком периоде активности ее летний спад не выражен. У личинок и нимф в странах с умеренным климатом активность продолжается весь теплый сезон с разной степенью выраженности весеннего и осеннего пиков (Gray, 1991; Kurtenbach et al., 1995; Talleklint, Jaenson, 1996a).
Сезонность активности у этого вида обеспечивается существованием морфогенетической диапаузы на фазах яйца, напитавшихся личинок, нимф и самок, так что выплод голодных особей происходит в основной массе во второй половине лета и осенью (Хейсин, 1955; Бабенко,' 1958; Белозеров, 1988). Важную роль в синхронизации активности имеет также поведенческая диапауза у голодных особей. Она факультативна, контролируется фотопериодическими условиями и определяет соотношение между частями популяции, активизировавшимися в сезон вылупления или только после зимовки. Поэтому в южных частях ареала, как например в Молдавии (Успенская, 1987), осенний пик активности выражен значительно сильнее, чем на севере, в Карелии (Хейсин и др., 1955). Благодаря поведенческой диапаузе голодные особи на всех фазах могут переживать до 1 года и более, тогда как сроки жизни активизировавшихся клещей около 1 мес. Сроки активации перелинявших особей, продолжительность сезона их активности и индивидуальной жизни неактивных и активных особей определяются гигротермическими условиями в их местообитаниях и поэтому значительно варьируют в границах видового ареала (рис. 56).
Рассмотренные выше процессы в свою очередь определяют смену состава популяций, которые наряду с особями из разных когорт одной и той же генерации могут включать и особей, принадлежащих к 2 и более последовательным поколениям. Состав популяции /. ricinus, ее обилие и активность голодных особей обусловлены сезонностью развития яиц и напитавшихся личинок и нимф, сроками активации перелинявших особей и скоростью отмирания активных голодных клещей (Lees, Milne, 1951; Cerny, 1957; Бабенко, 1958; Балашов, 1962; Daniel et al., 1977; Gigon, 1985; Gray, 1991).
Для развития каждой из фаз жизненного цикла в большинстве случаев требуется не менее 1 года. Зимовка возможна на всех фазах развития в голодном и напитавшемся состоянии. Однако напитавшиеся клещи и яйца обязательно должны находиться в состоянии диапаузы, так как инициация осенью процессов эмбриогенеза или метаморфоза ведет к гибели клещей в зимний период. Благодаря факультативности диапаузы в южных частях ареала развитие при благоприятных условиях может завершиться за 2 года (Daniel et al., 1977). На севере, наоборот, для завершения 1 или всех фаз может потребоваться по 2 года, и у клещей наблюдался 5—6-летний цикл развития (Хейсин, 1955).
Сходным многолетним циклом развития обладает широко распространенный в таежных лесах Восточной Европы и Азии /. persulcatus. Активность голодных имаго наблюдается весной сразу со сходом снега и до середины лета (рис. 57). Они происходят от нимф, перелинявших осенью прошлого года, но остающихся в состоянии облигатной диапаузы в течение осенне-зимнего периода. Личинки и нимфы (рис. 58, 59) активны на протяжении всего теплого сезона и могут активизироваться сразу после вылупления из яиц или линьки во второй половине лета или же после зимовки как голодных, так и напитавшихся особей. У личинок и нимф диапауза факультативна, соответствует нормам длиннодневной фотоперио- дической реакции и предотвращает линьку особей, напитавшихся во второй половине лета. Напитавшиеся самки и отложенные осенью яйца неспособны к зимовке и погибают. Предполагают, что у голодных имаго поведенческая диапауза обли- гатна и предотвращает осеннюю активность клещей.
У японской популяции /. persulcatus (Fujimoto, 1993, 1995b) имаго, развивавшиеся в условиях длинного дня в первые 3 мес после линьки, находились в состоянии поведенческой диапаузы. В природе она снималась после зимовки к следующей весне. Длина дня в период от откладки яиц и до встречи с хозяином
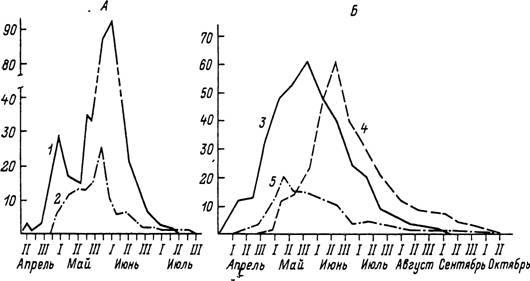
Рис. 57. Сезонный ход активности имаго Ixodes persulcatus (по: Таежный клещ..., 1985).
А — Московская область: У — с растительности в смешанном лесу; 2 — со скота. Б — Южное Приморье: 3 — с растительности в надпойменном кедровнике; 4 — с растительности в чернопнхтарннке; 5 — со скота. По вертикали — обилие
на 1 флаго-час (У, 3, 4) и на 1 животное (2, 5).
не вызывала поведенческой диапаузы у личинок. У голодных нимф, находившихся в условиях короткого дня, способность к питанию подавлялась. Режим короткого дня до, во время и после питания личинок тормозил линьку их в нимф, а в длинном дне развитие проходило без задержки. У напитавшихся нимф диапауза возникала при нахождении их в коротком дне до и во время питания. Условия содержания голодных нимф не влияли на их последующее развитие. Прочность поведенческой диапаузы голодных нимф и самок снижалась с увеличением времени, прошедшего после их линьки.
За 1 год обычно успевает закончить развитие только 1 фаза жизненного цикла, так что для завершения жизненного цикла требуется 3 года. В случае повторных зимовок на личиночной и нимфальной фазах, что нередко случается в условиях холодного лета и особенно у северных границ ареала, жизненный цикл 4- или 5-летний (рис. 60). В наиболее благоприятных условиях южных районов Дальнего Востока за теплый сезон успевают развиться яйца и вылупившиеся из них личинки
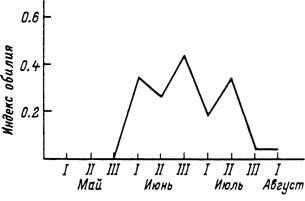
Рис. 58. Сезонные изменения обилия иимф Ixodes persulcatus на мелких млекопитающих в Красноярском крае (по: Таежиый клещ..., 1985).
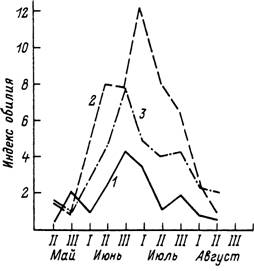
Рис. 59. Сезонные изменения обилия личинок Ixodes persulcatus иа мелких млекопитающих в Красноярском крае (по: Таежиый клещ..., 1985).
У — сосиово-листвеииый лес иа холмах; 2 — низинный темиохвойиый лес; 3 — равнинный лиственный лес.
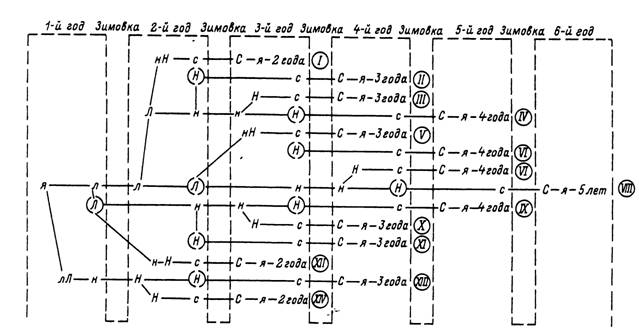
Рис. 60. Продолжительность жизиеииого цикла Ixodes persulcatus (по: Таежный клещ..., 1985).
л — яйца; л. н — голодные личинки и нимфы; Л, Н — напитавшиеся личинки и нимфы; (Л), (Я) — диапаузирующие напитавшиеся личинки и нимфы; с — неактивные самки и самцы; С — активные самки, самцы и напитавшиеся самки. Рил- сними цифрами в кружках обозначены варианты жизненных циклов.
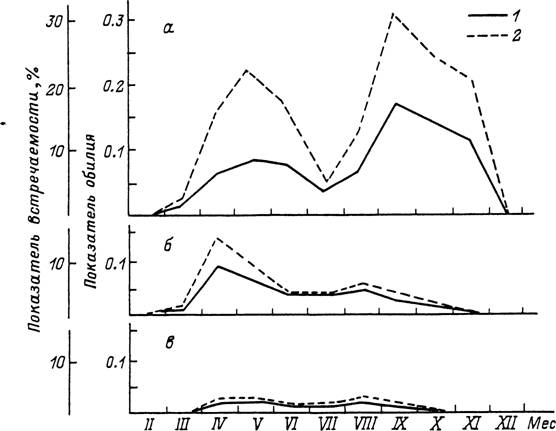
Рис. 61. Сезонность паразитирования клеща Ixodes trianguliceps на мелких млекопитающих в лесостеп ной зоне Среднего Поволжья (по: Бойко и др., 1982).
а — личинки; б — нимфы; в — самки; / — встречаемость: 2 — обилие.
и нимфы и развитие может уложиться в 2 года. Продолжительность жизненного цикла значительно варьирует не только в разных частях ареала, но и для особей одной и той же популяции (Таежный клещ..., 1985).
У /. trianguliceps, распространенного от Великобритании до реки Енисей на востоке в разных типах равнинных и горных лесов, продолжительность жизненного цикла может варьировать от 2 лет в Англии до 5 лет в Карелии. Активность всех фаз развития наблюдается круглогодично, включая и зимние месяцы (рис. 61). К зимовке способны напитавшиеся и голодные особи на всех фазах, а также яйца (Высоцкая, 1951; Бобровских, 1965; Randolph, 1975). В Швеции личинки паразитируют круглый год, кроме июля, с апрельским и сентябрьским пиками численности на прокормителях. Нимфы наиболее многочисленны на хозяевах весной, а летом и осенью их численность резко снижается. Самки также паразитируют круглый год с максимумом летом (Nilsson, 1974).
Дальше всех в приполярные области продвинулся паразит морских птиц /. игіае, и этому виду свойственны и наиболее продолжительные циклы развития. На побережье Баренцева моря развитие 1 поколения продолжается 5—7 лет (Карпович, 1973), в Ньюфаундленде — %—5 лет (Eveleigh, Threlfall, 1974) и в более теплом климате у побережья Шотландии — 2—4 года (Barton et al., 1996). На птичьих базарах контакт клещей с птицами возможен только в гнездовой период длиною в 2—4 мес. В северных частях ареала каждая из активных фаз питается лишь 1 раз в году и их дальнейшее развитие из-за низких температур местообитаний может занять 2 летних сезона. Зимуют голодные и напитавшиеся особи. В Шотландии большая часть напитавшихся личинок и нимф линяет в том же сезоне, а немногие личинки майских сроков насыщения успевают напитаться, перелинять в нимф, и последние также питаются в этом же сезоне. За их счет часть популяции имеет 2-годичный цикл, но основная масса 3-годичный и меньшая часть — 4-годичный. Зимуют в подавляющей массе голодные особи всех фаз развития.
Еще по теме Описания жизненных циклов:
- Синхронизация жизненных циклов с сезонными климатическими ритмами
- ПЕРИОДИЗАЦИЯ ЖИЗНЕННЫХ ЦИКЛОВ
- СЕЗОННОСТЬ И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНЕННЫХ циклов
- Экономический цикл и его фазы. Продолжительность циклов.
- 76. Жизненные перспективы личности.
- 45. Жизненная стратегия личности (К. А. Абульханова-Славская).
- 44. Жизненный путь личности как проблема
- 46. Методы исследования жизненного пути.
- Проблемы жизненного определения агрессивных старшеклассников
- Методика «Индекс жизненной удовлетворенности»
- Жизненный цикл организации
- 5.6. O возможности активизации жизненных сил ветеринарного врача
- Снижение содержания жизненно необходимых металлов,
- Последние стадии жизненного цикла семьи