СИМБИОНТНОЕ ПИТАНИЕ И ПИЩЕВАРЕНИЕ
Под симбнонтным питанием и пищеварением понимают участие животных или растительных организмов в обеспечении витания (или расщепления отдельных пищевых веществ) животного-хозяина.
Хотя эти симбионт- ные отношения лучше всего выражены в организме некоторых насекомых (термитов) и млекопитающих (жвачных), их можно наблюдать и у животных, стоящих на более низких ступенях эволюции.Довольно распространено у низших организмов — кораллов, турбсллярип, некоторых моллюсков симбионт-
нос питание при помощи зооксантелы и зоохлореллы. Одноклеточные зоохлореллы образуют благодаря фотосинтезу крахмал, который затем поглощается подвижными клетками — амебоцитами. Зоохлореллы поглощаются полипами.
Особенно большое значение приобретает симбионтное питание при усвоении целлюлозы. У большинства высших организмов нет фермента, расщепляющего целлюлозу— целлюлазы. Только некоторые организмы, в том числе дождевые черви, способны самостоятельно усваивать целлюлозу. Большинство же улиток (Helix) переваривает целлюлозу при помощи бактерий. Из насекомых жуки-усачи (Cerambycidae), златки (Buprestidae) и жуки-точильщики (Anobiidae) имеют в кишечнике целлю- лазу и могут питаться древесиной. Другая группа жуков— рогачи (Lamellicornia), личинки которых тоже питаются древесиной, содержат в кишечнике бактерии, расщепляющие целлюлозу.
Однако симбионтное питание не ограничивается использованием целлюлазы. У жвачных огромная масса инфузорий и бактерий, населяющая рубец, переваривается в сычуге и нижележащих отделах кишечного канала; так удовлетворяется около 1/3 общей потребности животного в белке.
Микроорганизмы рубца способны синтезировать белок из таких простых азотистых соединений, как мочевина, аммонийные соли и даже аммиак. Это служит основанием для использования синтетической мочевины при кормлении сельскохозяйственных животных. Есть основания полагать, что в слепой кишке грызунов происходят аналогичные процессы, и поедание цекотрофов кала служит источником белка, так как в основном це- котрофы СОСТОЯТ ИЗ тел микробов 'И инфузорий.
При содержании животных в стерильных условиях для них требуется большее количество белка. То же наблюдается и при исключении цекотрофии.Пребывание пищи в рубце жвачных характеризуется интереснейшим среди высших организмов примером симбиоза бродильных бактерий, инфузорий с макроорганизмом жвачного. Рубцовое «пищеварение» связано с деятельностью микроорганизмов, расщепляющих клетчатку, и с образованием глюкозы, молочной кислоты и летучих жирных кислот (уксусной, пропноновой, масляной, мура-
вышой и др.). При этом выделяется и значительное количество тепла. Если бактериальная флора рубца обеспечивает химическое расщепление клетчатки путем брожения, то инфузории механически расщепляют клетчатку, содействуя тем самым более полному ее расщеплению. Брожение происходит с образованием большого количества нелетучих жирных кислот и СО2. И те, и другие всасываются в кровь, а углекислота частично отрыгивается. Непрерывная секреция паротидных слюнных желез обеспечивает поддержание в рубце слабощелочной реакции (pH около 8,0), что является оптимальным для протекания бродильных процессов. Фактически размножение микробов и инфузорий приводит к тому, что перевариванию в сычуге и кишечнике подвергается огромное количество микробных тел и простейших, составляющих основной источник усвояемости жвачными азота (Lously, 1949; Renaux, 1955; Ennison, a. Lewis, 1962). Кроме того, некоторые микробы синтезируют аминокислоты из мочевины, а также витамины.
Около 40% энергетического обмена восполняется в организме жвачных за счет микрофлоры и простейших рубца. Кормление корнеплодами, содержащими много углеводов, усиливает процесс перехода растительных белков в животные белки тел микроорганизмов в рубце. Переход на грубые корма у новорожденных жвачных сопровождается появлением инфузорий в рубце. Так, по старым данным, появление инфузорий в рубце ягнят и телят можно наблюдать только с началом кормления сеном. Закономерная динамика нарастания количества инфузорий обнаруживается к 6-месячному возрасту животного с последующим падением числа простейших к 7—8 месяцам, когда их количество достигает величин, характерных для взрослого.
Корм задерживается в рубце в течение 5—6 ч (овцы). Реакция содержимого рубца при скармливании углеводов становится кислой, при скармливании грубого корма — щелочной. У овец бродильные процессы приобретают наибольшее развитие через 4 ч после начала пастьбы. Кормовая свекла увеличивает степень брожения и вызывает увеличение количества инфузорий (Ташенов, 1956). По данным той же работы, наименьшее брожение вызывает скармливание животным силоса. Это может объясняться тем, что в силосе уже есть большое количество перебродивших продуктов расщепления углеводов. Процессы нитрификации в рубце протекают крайне энергично. При этом усваивается как неорганический азот (например, азот аммиака), так и азот мочевины (Виноградова-Федорова, Ми- хина, Павлов, Солдатенков и Трофимова, 1935; Имше- нецкий, 1953; Ташенов, 1956).Таким образом, в преджелудках жвачных, поскольку у млекопитающих нет ферментов, расщепляющих клетчатку, осуществляется расщепление клетчатки вместе с населяющими преджелудки бактериями и отчасти инфузориями. Здесь обеспечивается переваривание до 45% всей клетчатки, поступившей с пищей. Сетка и книжка служат, по существу, фильтрами, обеспечивающими более полное разложение грубой пищи в рубце и препятствующими переходу ее 'В сычуг.
Как было сказано выше, переваривание клетчатки обеспечивается не только процессами, происходящими в рубце у жвачных (расщепление клетчатки микроорганизмами), но и расщеплением клетчатки в слепой кишке непарнокопытных и специальным механизмом цекотро- фии у некоторых видов грызунов. Было установлено расщепление целлюлозы в слепой кишке лошади и доказана роль бактерий в этом процессе (Scheunert, 1906).
Переваривание клетчатки у непарнокопытных (лошади) обеспечивается интенсивно развитой слепой кишкой, вмещающей до 37 л содержимого (вдвое больше желудка). По характеру процессов слепая кишка может быть уподоблена рубцу с той разницей, что интенсивность процессов брожения и нейтрализации образующихся при этом жирных кислот происходят за счет выделения кишечной слизи.
Частично процессы брожения протекают в толстой (ободочной) кишке. Пища задерживается в слепой кишке лошади до 72 ч, причем переваривается около 40—50% всей клетчатки, содержащейся в химусе. Здесь происходит переваривание химуса ферментами частично растительного, частично животного происхождения, поступающими из вышележащих отделов кишечного канала (Никулин, 1954). Однако и у жвачных слепая кишка является местом интенсивного брожения. Мигрирующие из лимфатических желез в просвет кишечника лимфоциты регулируют развитие флоры кишечника и тем самым процессы брожения (Синельников и Ивкина, 1954).По-видимому, сходные явления длительного задерживания пищевой массы (химуса) в слепой кишке происходят и у некоторых видов грызунов, питающихся зелеными частями растений (полевок, песчанок). Толстая и прямая кишка составляет у домовой мыши 18,6% общей длины кишечника, а у обыкновенной полевки — 38,3% (Рубин, 1953); в заднем отделе кишечника и слепой кишке идут процессы брожения и усвоения клетчатки.
Первые исследования значения слепой кишки для пищеварения у кролика принадлежат Цунтцу (Zuntz, 1905). Морфологические особенности слепой кишки указывают на чрезвычайное развитие этого органа у ряда видов грызунов (травоядные). Наибольшего развития слепая кишка достигает у травоядных форм — полевок. Удаление слепой кишки в асептических условиях приводило к быстрой гибели полевок, снижению веса тела у черной крысы, но не оказывало практически никакого влияния на состояние крыс-пасюков (В. К. Шепелева). Как известно, пасюк в значительной мере питается животной пищей и усвоение клетчатки имеет для этого вида наименьшее значение в ряду грызунов. Отсутствует слепая кишка у знмоспящих видов — сонь, однако для этих животных характерно исключительно зерноядное питание, главным образом орехами и семенами хвойных. У домашних кроликов многие исследователи (Morot, 1882) уже давно обнаружили поедание собственного кала (копрофагию) и скопление кала в области дна желудка.
При лишении возможности поедания части собственного кала кролики погибали в течение нескольких месяцев. Копрофагия играет важную роль в пищеварении многих грызунов: крыс, морских свинок и хомяков (Krzywanek, 1927). Кал кроликов состоит из двух частей: более плотной части — мелких шариков (pellets), которые поедаются животными, и более крупных, составляющих обычный кал (Briiggeman, 1937). Было высказано предположение (Morot, 1882), что два вида кала связаны в своем происхождении с однократным или двукратным прохождением его по кишечнику. Исследованиями Мадсена (Madsen, 1939) было установлено, что в кишечнике кролика формируются особые слизистые комки, которые заглатываются животными и откладываются в области дна желудка. На рисунке 75 показано распределение очищенных зернышек винных ягод (точки) и слизистых клубочков-катышков (pellets) в желудке кроликов, убитых в разные сроки введения этих зернышек в опорожненный желудок. Катышки заглатываются животным сразу же после выхода их из анального отверстия. Интересно, что в этих опытах «желудочные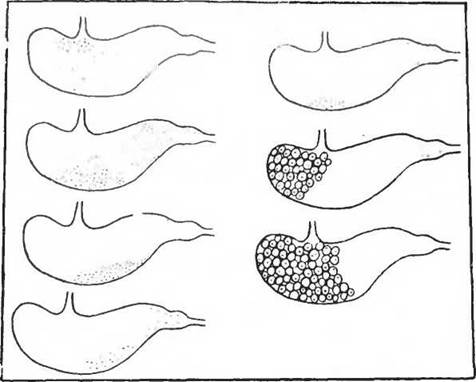
Рис. 75. Контуры желудка кролика (по Olsen a. Aladsen, 1944). Точки — распределение семян винных ягод через разные промежутки времени после введения в желудок; кружки — желудочные шарики-катышки; кружки с точками — то же с включением семян
шарики» были обнаружены в прямой кишке через 7 ч после кормления, а в желудке только через 12 ч после кормления. Некоторые из «желудочных шариков» представляли просто скопление слизи, в центре других находилось непереваренное зернышко.
Многократное прохождение пищи через кишечник кроликов доказывается и тем, что введенная в пищу соль меди (CuSO4) циркулирует в кишечнике кролика до 5 недель (Eden, 1941). Такая циркуляция обеспечивает как наиболее полное насыщение содержимого кишечника расщепляющими целлюлозу микроорганизмами, так и циркуляцию и использование образуемого микроорганизмами витамина Віг-
Вторичное пережевывание выделяемых шариков-катышков кроликами и зайцами рассматривалось некоторыми исследователями, как так называемая псевдожвачка, а роль всех этих явлений в усвоении целлюлозы начисто отрицалась.
Так как шарики-катышки поедаются животными прямо из анального отверстия, в нормальных условиях видеть эти образования невозможно. Специальные формы эксперимента — деревянные воротники, позволили некоторым исследователям (Olsen a. Aladsen, 1944; Harder, 1950 и др.) не только отделить эти образования от кала, но и установить значение поедания и двукратного прохождения части пищи через кишечник. Олсен и Мадсен обнаружили, что одевание воротника на кроликов ведет к падению веса и привес в процессе развития уменьшается. Они рассматривали эти явления как результат уменьшенного снабжения организма витаминами, которые образуются в результате деятельности бактерий и поедаются животными. Однако более детальные исследования (Harder, 1950; Frank, Hadeler a. Harder, 1951; В. К- Шепелева) установили несомненную роль слепой кишки в происхождении упомянутых выше шариков-катышков.Хардер приводит довольно противоречивые данные о влиянии выключения цекогрофпи на рост и поддержание веса у грызунов. У лабораторных мышей надевание воротника, исключающего поедание шариков-катышков, быстро приводит к гибели; у кролика в зимний период можно наблюдать значительное падение веса, в летний период при поедании зеленого растительного корма падения в весе почти не происходит. По-видимому, здесь имеют значение и растительные ферменты расщепления клетчатки, содержащиеся в зеленом корме.
Некоторые исследователи считают копрофагию у кроликов (домашних и диких), а также зайцев нормальным физиологическим явлением и критерием полноценности пищи в отношении содержания витаминов группы В (Anghi, 1957; Rodel, 1957).
У бобров поедание кала наблюдается главным образом при питании травой, в меньшей мере — картофелем и еще меньше при введении кефирной подкормки (Шид- ловская, I960). Автор считает, что копрофагпя (цекотро- фпя) обеспечивает витаминный обмен в организме, главным образом витаминов группы В (В12). Поедание так называемого мягкого кала (цекотрофа) бобрами происходит непосредственно из клоаки.
На основании детального исследования поедания катышков, их химического состава выдвинута теория цекотрофии, т. е. значения образующихся в слепой кишке шариков-цекотрофов в пищеварении грызунов (Harder, 1950). Цекотрофы содержат значительное количество белка, большое количество бактериапьных тел (кал — 2700 млн. бактериальных тел, цекотрофы — 95G0 млн.). Цекотрофы, формирующиеся в слепой кишке, поедаются затем животными сразу после выхода из анального отверстия и откладываются в слепом конце желудка. Оттуда отдельными порциями они поступают в кишечный канал и поддерживают на протяжении всего пищеварительного канала расщепление целлюлозы. Наиболее развит (да и лучше всего изучен) механизм цекотрофии у кролика, хотя есть все основания придавать ему большое значение у всех без исключения травоядных грызунов.
Еще по теме СИМБИОНТНОЕ ПИТАНИЕ И ПИЩЕВАРЕНИЕ:
- Органы пищеварения и питания.
- Особенности организации питания, медицинский контроль и гигиенические требования к рациону питания детей и подростков в образовательных учреждениях
- Организация диетического питания и питания по лечебному пайку
- Часть II. Неправильное питание и его последствия Заболевания, связанные с употреблением в пищу недоброкачественных продуктов питания
- Нарушения кишечного пищеварения
- Параграф третий. Причины слабости пищеварения
- Параграф первый. Порча пищеварения
- Параграф пятый. Признаки расстройства пищеварения
- Параграф второй. Расстройство пищеварения
- Нарушение мембранного пищеварения, всасывательной и выделительной функции кишок
- Патология системы пищеварения
- Тема3. Система органов пищеварения
- Глава 22. Патологическая физиология пищеварения
- Слюноотделение при патологии пищеварении
- Врожденные пороки органов пищеварения