ЭНЕРГЕТИЧЕСКИЙ РАСХОД ОРГАНИЗМА
ПойкнлОтермные животные. Потребность в пище и ее удовлетворение тесно связаны с энергетическими затратами, в свою очередь зависящими от ряда факторов внешней среды, из которых главное место занимает температура.
Кроме того, энергетические затраты животного зависят от размеров тела, подвижности, ряда сложных биологических циклов (суточного, сезонного, лунного и т. д.), стадии развития в онтогенезе, условий питания, а также от наличия других организмов того же вида. Эти очень сложные связи не могут быть сведены к какой-либо элементарной схеме, тем более, что безусловно не существует никакой зависимости между уровнем энергетических затрат (потреблением кислорода) и уровнем эволюционного развития (Hemming- sen, 1950).Наиболее общая зависимость, которая здесь наблюдается,— это правило поверхности тела, установленное впервые для гомойотермных организмов и затем широко исследованное для пойкилотермных (Hemmingsen, 1950). В основе этого правила (закона) лежит факт, что между весом тела и уровнем энергетического обмена есть логарифмическая зависимость. Так как поверхность тела представляет собой экспоненциальную функцию веса тела, то при изображении такой кривой в логарифмическом масштабе можно наблюдать прямую, наклон которой зависит от возрастания уровня обмена в связи с весом тела. На рисунке 76 приведены данные этих зависимостей, полученные для одноклеточных (пойкилотермных и гомойотермных) организмов. Ясно видно, что увеличение размеров (веса) организма приводит к относительному снижению общего обмена веществ. Это наблюдается на бактериях, жгутиконосцах, амебах, инфузориях и яйцах морских животных и насекомых.
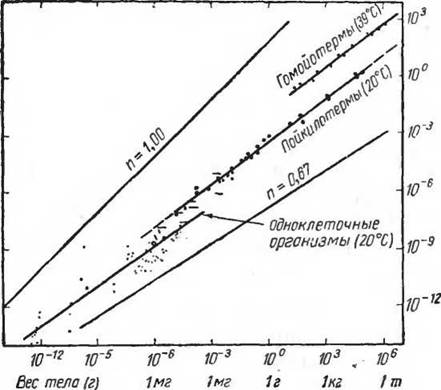
Рис. 76. Потребление кислорода у одноклеточных пойкилотермных и гомойтермных организмов в зависимости от веса тела (по Ilemmingsen, 1959)
Объяснение этому обычно видят в способности кислорода проникать внутрь свободно живущей клетки.
Однако у многоклеточных интенсивность обмена уже зависит от поверхности.Появление органов дыхания еще более усложняет положение (Krvwienczyk, 1956). При жаберном дыхании моллюсков (L amellibrcnichiata), ракообразных (Isopo- da) потребление кислорода зависит ог поверхности гела. У насекомых, дышащих через трахеи, окислительные процессы в большей степени зависят от веса тела. Тем не менее для пойкилотермных организмов, как и для гомой- отсрмпых, обмен веществ в покое может быть рассчитан по формуле
M = KW\
где М— общий обмен веществ (кал), W— вес тела, b — константа, отличающая наклон кривой, полученной в эксперименте, от кривой соотношений веса (по оси абсцисс) и теоретического потребления кислорода (по оси ординат) в логарифмических выражениях. Константа К показывает уровень окислительных процессов, на котором ведется отсчет. Для млекопитающих она равна примерно 10. Величина b обычно меньше 1,0 и у млекопитающих равна 0,70. У пойкилотермных величина b также меньше 1,0. Так, для амфибий она равна 0,738, для моллюсков и равноногих раков (Isopoda) около 0,66, для насекомых около 1, что означает, что закон поверхности тела для них практически отсутствует (Bertalanffy, 1949). Впрочем, у насекомых разного веса эти отношения оказываются различными. Так, у жуков-чернотелок (Tenebrionidae) метаболизм возрастает на единицу веса до 5 мг, затем быстро снижается до 45 мг и остается постоянным до 140 мг (Chauvin, 1956). У насекомых с полным превращением b приближается к 1,0, а у насекомых с неполным превращением этот коэффициент ниже. Так, у тараканов и палочников он равен примерно 0,8 (Enger a. Savalov, 1958). Для очень мелких беспозвоночных весом менее 40 мг коэффициент b равен 0,95 (Zeuthen, 1953). У более крупных ракообразных значение b снижается, т. е. выявляется правило поверхности тела.
В последнее время выявлены зависимости между интенсивностью дыхания и содержанием азота в теле животных (Zeuthen, 1953). С увеличением содержания азота интенсивность газообмена снижается.
Это хорошо выявлено у рыб, ракообразных и насекомых.Механизмы, лежащие в основе изложенных выше зависимостей, неясны. В исследованиях дыхания тканей установлена обратная корреляция между интенсивностью Q О2 птиц и размерами тела (Bertalanffy a. Pirozynski, 1951). Некоторые ткани (печень, мозг, мышцы) дышат с одинаковой интенсивностью у рыб разного веса (Vern- berg, 1954).
Вместе с тем наблюдается четкая зависимость между количеством митохондрий на вес ткани и общим обменом веществ. Это же относится и к содержанию некоторых окислительных ферментов (сукциноксидазы, дегидрогеназы) (Prosser a. Brown, 1962).
Таким образом, у пойкилотермных организмов такая же тенденция к зависимости уровня обмена от поверхности тела, как и у гомойотермных. Однако очень большое значение приобретает механизм дыхания — снабжение кислородом отдельных тканей, что приобретает особое значение и в связи с тем, что большинство пойкилотермных организмов подчиняет уровень окислительных процессов содержанию кислорода в окружающей среде (конформация дыхания), т. е. потребление кислорода зависит от его доставки (Tang, 1931). Особенно ярко это выражено у беспозвоночных, не имеющих дыхательных пигментов. Впрочем, у многих форм пигмент насыщается в зависимости от содержания кислорода в среде и потому тип дыхания становится зависимым (конформируемым).
Гомойотермные животные. Общая величина энергетических затрат организма складывается из затрат энергии в полном покое, дополнительного расхода энергии на пищеварение, всасывание, ассимиляцию, дополнительного расхода на терморегуляцию, расхода на поддержание положения тела, сопротивление току воды и т. п. затраты энергии в полном покое — основной обмен.
Понятие об основном обмене, разработанное главным образом для человека, лишь с большим трудом может быть применено к животным и особенно к тем формам, у которых процессы пищеварения и всасывания идут непрерывно. Именно для этих животных состояние голодания (состояние «натощак») не является физиологическим и достигается уже в результате настоящего голодания с распадом собственных тканей тела.
Так обстоит дело, например, у всех копытных. Некоторые грызуны (кролики, зайцы) при лишении их пищи также очень долго сохраняют в желудке пищевые массы. По-видимому, это касается и многих других травоядных млекопитающих. Изучение основного обмена v всех этих животных возможно только на какой-то стадии пищеварения и всасывания и, следовательно, сравнивать их с хищными, обезьянами и человеком как с организмами с фазовым типом пищеварения и всасывания очень трудно.Другой стороной рассматриваемого вопроса является мышечная деятельность. Для многих мелких видов млекопитающих и птиц исключить двигательную активность практически невозможно. Многие мышевидные грызуны, некоторые мелкие хищные (хорьки, ласки, виверровые), воробьиные птицы очень редко остаются в состоянии покоя, а при исключении мышечной деятельности у них значительно снижается температура тела.
Учение об основном обмене — следствие работ дерпт- ских физиологов середины XIX столетия Биддера и Шмидта, установивших у собаки удивительно постоянный уровень метаболизма изо дня в день. Эти факты были подтверждены большим числом исследователей.
Под основным обменом всегда понимают величину энергетических затрат (в единицу времени), отнесенную к весу тела животного. Таким образом, величины основного обмена всегда отражают интенсивность тканевых окислительных превращений. Общеизвестно, что уровень обмена веществ, рассчитанный на единицу веса тела, резко падает с увеличением веса тела животного, с уменьшением его относительной поверхности. Поскольку последняя является важным фактором, определяющим величину теплоотдачи животных, возник важнейший биологический принцип, получивший название закона поверхности тела. Впервые положение о значении поверхности тела для уровня обмена веществ было сформулировано более 100 лет назад (Sarrus et Rameaux, 1837, 1838; Rameaux et Sarrus, 1838, 1839). Эти исследователи впервые сделали вывод, что уровень теплоотдачи зависит от величины «свободной поверхности животного», а потребление кислорода пропорционально количеству вдыхаемого воздуха.
Отсюда был сделан вывод, что потребление кислорода пропорционально потере тепла и, следовательно, зависит от соотношений веса и поверхности тела. Интересно, что эти авторы включили в число факторов, определяющих поверхность тела, еще и общую поверхность легких и количество дыханий. Саррю и Рамо представили взаимоотношения возрастания веса и снижения при этом числа пульсовых ударов и частоты дыханий, продукции и потерн тепла по формуле:Y = a\F 2/3,
где У— величины перечисленных физиологических функций, а — константа для каждого животного, U’—вес тела (2). Таким образом, первые положения о законе поверхности связывались с уровнем основных физиологических функции животных. Дальнейшие исследования ряда физиологов подтвердили эти положения о связи величин обмена веществ и величин, характеризующих легочное дыхание и кровообращение.
Важным этапом в развитии представлений о роли поверхности тела организмов в регуляции их жизнедеятельности явились положения Бергманна (Bergmann, 1847—1848). Бергманн, впервые употребивший термины «гомойотермний» и «пойкилотермный», обратил внимание на то, что первые от полярных стран к тропическим (у близких в систематическом отношении видов) имеют все меньшие размеры (вес) тела; у пойкилотермных организмов, наоборот, тропические формы обладают большими размерами тела, чем обитатели северных широт. Эти закономерности важны не только для эколого- и зоо- географических сопоставлений, но и служат основанием для физиологического истолкования поверхности тела как важнейшего фактора, определяющего обмен веществ в организме.
Правило Бергманна было использовано для физиологического исследования обмена веществ на собаках (Rubner, 1883) и кроликах (Richet, 1889). На сельскохозяйственных животных и птицах было выявлено значение поверхности тела для интенсивности охлаждения и повышения обмена вєщєсте при низких температурах среды (Kleiber, 1932).
Формулировка закона поверхности тела принадлежит Рубнеру (Rubner, 1883).
Он обосновывал закон поверхности тела тем, что при расчете на единицу поверхности тела обмен веществ собаки остается почти постоянным, тогда как при расчете на единицу веса он резко изменяется. Положения основывались на исследовании теплопродукции в полном покое и в голодном состоянии у 7 собак весом от 31 до 3 кг (табл. 32). Такие же явления наблюдаются и при сопоставлении теплопродукции у животных разных видов (табл. 33).На рисунке 77 приведены данные величин основного обмена (Benedict, 1938), нанесенные на полулогарифмическую шкалу. Из схемы видно, что обмен веществ, рас-
Таблица 32
| Сибакм (№) | Вес тела (кг) | Теплопродукции (кал за 24 ч) | |
| на I кг | ня 1 м~ поверхности тела | ||
| 1 | 31,20 | 35,68 | 1036 |
| 2 | 24,00 | 40,91 | 1112 |
| 3 | 19,80 | 15,87 | 1207 |
| 4 | 18,20 | 46,20 | 1097 |
| 5 | 9,61 | 65,16 | 1183 |
| 6 | 6,50 | 66,07 | 1153 |
| 7 | 3,19 | 88,07 | 1212 |
Таблица 33
Теплопродукция у животных разных видов
| Объект | В.с тела (кг) | Теплопртукция (кал за 24 ч) | |
| на 1 кг веса тела | на 1 м* п верхно- сти тела | ||
| Лошадь.............................. | 441 | 11,3 | 948 |
| Свинья .............................. | 128 | 19,1 | 1078 |
| Человек ............................ | 64,3 | 32,1 | 1042 |
| Собака .............................. | 15,2 | 51,5 | 1039 |
| Кролик............................... | 2,3 | 75,1 | 776 |
| Г усь.................................. | 3,5 | 66,7 | 969 |
| Курица .............................. | 2,0 | 71,0 | 943 |
Теплопродукция у собак различною веса (по Rubner, 1883)
считанный на единицу поверхности тела, возрастает очень медленно от самых маленьких животных (мышь, воробей и т. д.) к средним (до 100 кг), но затем у крупных животных (копытных — пони, першерона, быка, слона) — очень резко. Следовательно, относительно очень малая поверхность тела не является фактором, снижающим уровень окислительных процессов. Для протекания жизненных процессов гомойотермного организма существует какой-то минимум, связанный, по-видимому, с возможностями ферментативных реакций и с процессами возбуждения, ниже которого в нормальных условиях жизни обмен веществ понижаться не может (Слоним, 1952). Поэтому очень крупные гомойотермные животные в условиях наземного существования представляют значительную редкость (другое дело — водные организмы,
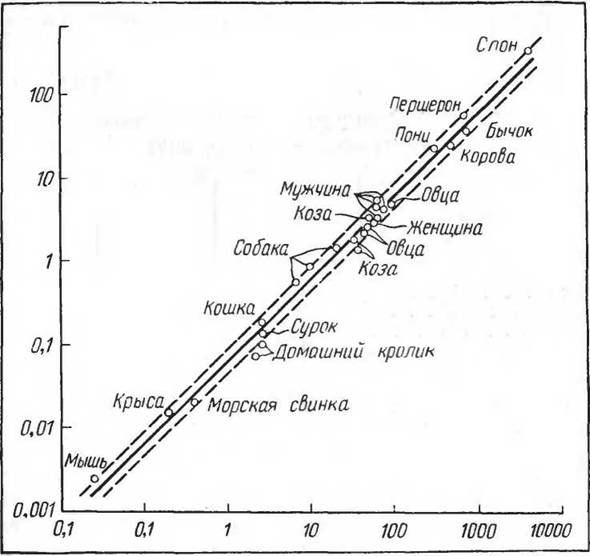
Рис. 77. Соотношения между пульсом, уровнем обмена и величиной тела у млекопитающих (по Brody, 1945). По оси ординат — отношение потребления О2 (мл) к частоте пульса; по оси абсцисс — вес тела (кг)
у которых теплоотдача в воде значительно более интенсивна), а уровень обмена веществ у них, рассчитанный на поверхность тела, оказывается в значительной мере сниженным по сравнению с животными среднего веса, представляющими огромное большинство ныне живущих видов. Вместе с тем отношение уровня основного обме-
на и частоты пульса у гомойотермных организмов удивительно постоянны (см. рис. 76) и нанесенные на логарифмическую шкалу (по весу тела) представляют такую же прямую зависимость, как и величина тела и потребление кислорода у пойкилотермных.
Были изучены газообмен и некоторые физиологические функции у самых маленьких животных — землероек весом около 3,5 г. В таблице 34 дано сравнение самых маленьких млекопитающих с животными среднего и крупного веса.
Таблица 34
Основной обмен у различных млекопитающих (по Morrison a. Pearson, 1947)
| Объект | Btc (кг) | Основной < бмен (кал) на: | |
| кг/сутки | м*/сутки | ||
| Бык..................................... | 300 | 12 | 1090 |
| Человек ............................. | 65 | 2 | 920 |
| Макак................................. | 4,2 | 49 | 670 |
| Морская свинка .... | 0,41 | 86 | 710 |
| Мышь белая....................... | 0,021 | 170 | 530 |
| Землеройка (Blarina) | 0,020 | 370 | 1020 |
| Землеройка (Sorex) . . | 0,0035 | 830 | 1570 |
Согласно этим данным, землеройки двух исследованных родов Blarina и Sorex сильно отличаются по своим физиологическим особенностям от близких им по весу тела грызунов. Авторы объясняют это чрезвычайной прожорливостью землероек, не могущих оставаться без пищи дольше 10—11 ч.
У крупных животных ритм сердечных сокращений уменьшается в связи с весом животного (Brody, 1945; Аршавский и Пяткин, 1958). Изменение сердечной деятельности крупного рогатого скота в онтогенезе связано с развитием и возбудимостью антигравитационной мускулатуры. Зависимости между весом и поверхностью тела и частотой пульса и дыхания не менее сложны, чем эти же зависимости при основном обмене веществ.
Закон поверхности тела может быть прослежен на разных этапах развития млекопитающих; его нельзя 354
установить, вероятно, только для летучих мышей (Слоним, 1952). У Chiroptera летательные перепонки, периодически наполняющиеся кровью и периодически — во время состояния гомойотермии и полета — отдающие значительное количество тепла, в состоянии пойкилотер- мии и дневного сна не принимают сколько-нибудь значительного участия в теплоотдаче. В таблице 36 приведена теплопродукция у летучих мышей (2 вида) в состоянии гомойотермнії 'в сопоставлении с теплопродукцией желтогорлон мыши (Apodemus flavicollis), близкой ей по весу.
Таблица 35
Газообмен и теплопродукция у летучих мышей и желтогорлон мыши
(по Слониму, І952)
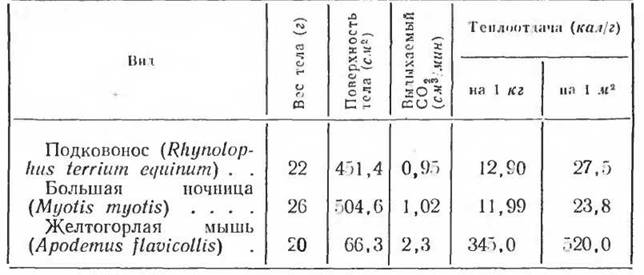
Несмотря иа большую поверхность тела летучих мышей, их основной обмен при температуре среды около 30° С в гомойотермном состоянии оказывается более низким, чем у мыши. Это объясняется тем, что температура большей части поверхности тела летучей мыши очень низка — почти совпадает с температурой внешней среды. Условия кровообращения в летательных перепонках летучей мыши таковы, что в покое (при определении основного обмена) отдача тепла происходит почти исключительно с поверхности туловища.
Правило Бергманна, хорошо подтвержденное современными исследователями (Allen, 1906; Botticher, 1915; Rensch, 1932, 1939; Лукин, 1940), в известной мере отражает значение климатического фактора для формирования высших организмов. Вместе с тем вся организация животного — его относительный вес и длина кишечника, поверхность легких, вес сердца, печени и т. д. тесно связана с основными жизненными процессами в организме, с их интенсивностью. Эти особенности развиваются в процессе эволюции или параллельно, или в тесной зависимости от основных энергетических потребностей животного организма. Способность организма отдавать и терять тепло в окружающую среду определяет в значительной мере тепловой баланс организма, а потому величина поверхности тела в установлении уровня общей жизнедеятельности гомойотермных организмов несомненно играет выдающуюся роль.
Рассмотрение большого материала внутри отдельных отрядов (близких в систематическом отношении видов) позволяет выявить значение закона поверхности тела как общей закономерности, свойственной всем без исключения организмам. Однако на эту зависимость накладываются влияния экологического порядка, связанные с особенностями питания, пищедобывательной деятельностью и с длительным воздействием температуры среды обитания.
Еще по теме ЭНЕРГЕТИЧЕСКИЙ РАСХОД ОРГАНИЗМА:
- 6.3. Расстройство энергетического гомеостазав опухолевом организме
- Энергетический баланс
- Нарушения энергетического обмена
- Контроль за энергетической адекватностью питания.
- Компенсаторные реакции при дефиците энергетического субстрата и кислорода
- 2.1.4. Тестирование энергетических возможностей
- Энергетическая и направляющая компонента мотивации
- Прогнозирование, появление и развития заболеваний по энергетическим характеристикам биологически активных точек
- Пример 3: Объяснение расходов в здравоохранении
- Налоговые льготы по расходам на исследования и разработки.
- Справедливость в расходах на здравоохранение на душу населения