АДАПТИВНЫЕ ОСОБЕННОСТИ ТЕПЛООТДАЧИ И СЛОЖНЫЕ ФОРМЫ ТЕРМОРЕГУЛЯЦИИ
Отдача тепла во внешнюю среду происходит тремя путями. Во-первых, конвекцией — нагреванием окружающего воздуха, когда нагретый воздух расширяется и поднимается вверх, а новые порции холодного воздуха поступают на его место и омывают тело животного.
Таким образом, чем больше разница температур кожи и окружающего воздуха, тем интенсивнее происходит теплоотдача. Во-вторых, радиацией, которая заключается в том, что между излучением более холодных окружающих предметов и более теплой поверхностью тела животного устанавливается известное равновесие, при котором животное отдает больше тепла с длинноволновой радиацией и получает меньше тепловой энергии от окружающих предметов. Этот второй путь также очень важен. Некоторые исследователи считают, что около 50% всего тепла в обычных условиях среды отдается путем тепловой радиации. В-третьих, испарением воды с поверхности кожи, слизистых, легких. Это основной путь отдачи тепла при высокой температуре среды, когда отдача тепла конвекцией и радиацией невозможна. Значение таких физических путей отдачи тепла в природных условиях может резко изменяться как для животных, так и для человека.Теплоотдача с поверхности тела осуществляется по правилу Ньютона:
Tb-Ta=KXJXH,
где Ть — температура тела животного; Та — температура окружающего воздуха; К—константа; / — устойчивость
к потере тепла (теплоизоляция), И — теплопродукция. Зная величину критической точки и температуру тела, можно вычислить величину теплоизоляции, т. е. температурный перепад, который не оказывает влияния на теплопродукцию животного и человека.
По американским данным (Irving, 1964), для человека эта температура оказывается
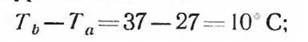
для арктического грызуна лемминга
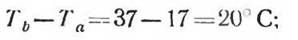
для арктического хищника песца
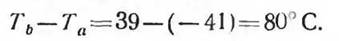
По данным таких расчетов, арктические животные могут практически не изменять обмена веществ при очень низких температурах за счет весьма ограниченной теплоотдачи.
Но схема построена на основе весьма грубых расчетов п может быть только иллюстрацией значения теплоизоляции.Прежде всего в условиях основного обмена, т. е. у голодных и неподвижно сидящих животных никогда не поддерживается ректальная температура (температура тела) на постоянном уровне. Это касается не только мелких животных — мышевидных грызунов, воробьиных птиц и т. д., но даже таких крупных, как верблюды, лошади, овцы. Более постоянная температура у представителей семейства собак (Canidae), например домашних собак, лисиц, песцов. В табл. 10 приведены колебания температуры тела в зоне температур от 15 до 30° С при 2—3-часовой экспозиции животных в условиях основного обмена.
Таким образом, расчеты американских исследователей (Scholander a. oth, 1950; Irving, 1964; Hart, 1964 и др.), несмотря на их привлекательную простоту, не отражают всей сложности процессов, происходящих в организме при короткой экспозиции животного организма в разных температурных условиях среды.
Вторым возражением против этой схемы является вопрос о длительности воздействия холода. В многочисленных исследованиях (Слоним, 1952, 1962) было показано, что первые применения холодовых воздействий (если раздражитель не очень силен) вызывают обычно лишь слабо
Колебания температуры тела у некоторых видов млекопитающих при экспозиции в зоне температур от 15 до 30° С (по Слониму, 1952)
| Животное | Колебания ректальной темпера іvpw ( С)’ | Ав гор |
| Утконос (Ornithorhync- hus anatinus).................................. | 3,5 | Мартин, 1902 |
| Ехидна (Echidna аси- | ||
| Icata).................................. | 3,0 | » |
| Летучая мышь (Myotis myotis) ................... | Более 20 | Слоним, 1952 |
| Еж европейский (Eri- naceus europaeus) . . . | 8,0 | » |
| Полевка общественная (Microtus socialis) . . | 2,2 | » |
| Ондатра (Ondatra zi- bethica) | 6.2 | » |
| Индийская крыса (Nesokia indica) | 8,5 | » |
| Крыса лабораторная Большая песчанка | 2.8 | Слоним, 1952 |
| (Rhombomys opimus) | 4,2 | » |
| Собака домашняя . . | 1.2 | » |
| Шакал (Canis aureus) Песец (Alopex Iago- | 1.0 | » |
| pus) | 0.2 | |
| Барсук (Meles tneles) Гамадрил (Papio ha- | 0.5 | |
| madrylis) ................... | 2.2 | » |
| Макак-резус (Macaca mulatto.) | 2,3 | » |
выраженную реакцию обмена веществ. Примером может быть собака — животное, у которого вообще химическая терморегуляция достаточно хорошо выражена. Еще более это заметно у человека при сравнительно коротком воздействии слабых холодовых раздражителей. Лучше выявляется реакция химической терморегуляции у грызунов— лабораторных крыс, мышей. У многих представителей диких грызунов, особенно субтропических, например незокии {Nesokia indica), александрийской крысы {Rattus rattus alexandrinus), туркестанской крысы {Rattus turkestanicus), большой песчанки {Rhombomis opi-
7—1204
193
mus), песчанок малоазиатской (Meriones tristrami) и краснохвостой (Meriones erythrourus), первые применении холодового раздражителя вызывают слабо выраженное повышение газообмена, даже несмотря на значительное снижение температуры тела. Слабо выражена химическая терморегуляция и у тропических видов обезьян, например макак-резусов и макак-лаиундер.
Отсутствие значительной реакции терморегуляции при кратковременных воздействиях характерно для всех гомойотермных организмов.
Значительное изменение реакции терморегуляции можно наблюдать при ориентировочной реакции животных (резкое охлаждение может вызывать такую реакцию), эмоциональном возбуждении и т. д. В этой связи следует отметить значительные колебания температуры тела, главным образом функциональную гипертермию, часто наблюдаемую при измерении температуры у диких и мало приученных к этой процедуре животных, например обезьян, лемуров и даже рептилий (Britton a. Klein, 1939; Слоним, 1952). При повторных измерениях (приучении животных) эта гипертермия может исчезать, а при воздействиях холода возрастает реакция на это воздействие, т. е. повышение газообмена.
Весьма важное место в регуляции теплоотдачи занимает изменение легочного дыхания — его резкое учащение при воздействии на организм тепла и замедление при воздействии холода. Реакция дыхания — полипноэ, изучалась неоднократно и служит часто простым и надежным критерием «напряжения терморегуляции», так как обычно появляется тогда, когда возможности регулирования температуры тела снижением обмена оказываются исчерпанными (Слоним, 1952).
При понижении температуры среды у животных, хорошо поддерживающих температуру тела и приспособленных к охлаждению, наблюдается углубление и уреже- ние дыхания (Калабухов, 1944; Раушенбах, 1958). Эта реакция, как бы обратная термическому полипноэ, еще мало изучена. Она сводит до минимума потери тепла испарением с поверхности легких и ведет к повышению коэффициента утилизации кислорода в легких. Таким образом уменьшаются потери тепла во внешнюю среду. Отдача тепла легкими — изменением дыхания — при высоких температурах особенно выражена у хищников, хотя полипноэ наблюдается в какой-то мере и у всех остальных млекопитающих. Нет полипноэ только у обезьян, непарнокопытных и у человека. Между реакцией полипноэ и потоотделением у животных существует обратная зависимость: если функционируют потовые железы и они хорошо развиты, то полипноэ отсутствует и наоборот.
Между особенностями протекания полипноэ у млекопитающих различных отрядов установлены существенные различия. Так, полипноэ хищных выражено резче; частота дыхания достигает огромных величин (до 600 дыханий в 1 мин), дыхание очень поверхностное. Однако у хищных полипноэ не может продолжаться очень долго, так как требует большого расхода энергии и ведет в конечном счете к явлениям перегревания. Особенность полипноэ хищных (особенно у Canidae) заключается в том, что оно возникает при почти не изменившейся под влиянием нагревания температуре тела (Richet, 1898; Anrep a. Ham- niouda, 1933; Нестеровский и Слоним, 1935: Веселкин, 1945). Полипноэ сопровождается отделением жидкой слюны (Парфенов, 1906; Александров, 1954; Уждавини, 1960).Для полипноэ характерны значительная акапния и сдвиги щелочно-кислотного равновесия. Обнаружено, что у собак в молодом возрасте при полипноэ наблюдается большее падение содержания СОг в крови, чем у старых. Оно сопровождается значительным учащением пульса. Нарушенный водный баланс организма оказывает влияние на полипноэ. В опытах на собаках установлено, что газообмен, легочная вентиляция и частота дыхания при недостатке воды в организме снижаются. У грызунов полипноэ всегда возникает на основе значительного повышения температуры тела, хотя дыхание и становится более поверхностным. По-видимому, у них полипноэ также не может долго поддерживать температуру тела при нагревании, и животные гибнут от гипертермии (Stigler, 1930).
Полипноэ у насекомоядных (ежей) изучено Веселкиным (1945) и Филатовой (1949в). Частота дыхания у ежей при нагревании увеличивается до 240 в 1 мин. Температура тела при этом возрастает с 34,5 до 40,0° С. У жвачных полипноэ отличается сравнительно небольшой частотой дыхания — обычно до 200 в 1 мин — и очень большой продолжительностью этой реакции. У крупного
7*
195
рогатого скота, овец, коз полипноэ может продолжаться всю жаркую часть дня и особенно усиливается при инсоляции. Для жвачных при полипноэ характерно усиление теплообразования в межреберных мышцах, компенсирующееся теплоотдачей; таким образом, кровь, направляющаяся к коже, проходит в большом количестве через межреберные артерии в межреберные мышцы (Goodall a.
Young, 1954) с последующим разветвлением в коже на спине и на боках (тип «чудесной» сети) (рис. 45).При резких термических воздействиях обычно наблюдаются очень яркие проявления сосудистой регуляции. Особенно это выражено у человека, но отчасти наблюдается и у животных. В последнее время установлены и специальные механизмы — «теплообменники» в конечностях. При этом артерия, окруженная венами, отдает тепло в венозную кровь и таким образом в зависимости от просвета вен в большей или меньшей степени нагревает самую конечность. Подобные «теплообменники» существуют, по-виднмому, в ряде органов животных: хвосте у грызунов, конечностях у морских млекопитающих, ушах у жвачных и т. д. Схема такого органа представлена на рисунке 46.
На основании большого экспериментального материала показано, что метаболическая реакция при охлаждении начинается лишь после того, как исчерпаны возможности физической терморегуляции, в том числе и теплоизоляции, сосудистой реакции и т. д. (Hart, 19646). Однако эта схема едва ли может быть принята в связи с наличием кортикального механизма терморегуляции, сильно изменяющего эти взаимоотношения на первых этапах адаптации к теплу и холоду (Быков и Слоним, 1960).
Еще по теме АДАПТИВНЫЕ ОСОБЕННОСТИ ТЕПЛООТДАЧИ И СЛОЖНЫЕ ФОРМЫ ТЕРМОРЕГУЛЯЦИИ:
- Цитокины, влияющие на терморегуляцию, механизм их воздействия
- 52. Сложные нарушения развития. Подходы к классификации. Психическое развитие при сложных нарушениях
- Возможные пути воздействия вирусов герпеса на терморегуляцию
- Адаптивна організація менеджменту
- Особенности и различные формы организации психотерапевтической помощи.
- 32. Понятие, формы и особенности установления психологического контакта.
- Билет 55. Особенности и различные формы организации психотерапевтической помощи.
- Спектр мутаций в гене LRRK2 и особенности клинического течения LRR^-ассоциированной формы БП
- Билет 46.Шкала социально-психологической адаптивности.
- 14.1.4. Адаптивность
- Состояние врожденного и адаптивного иммунитета при эритемах
- Синаптическая пластичность как основа адаптивных возможностей нейронных сетей
- Параграф пятый. О сложных болезнях
- Параграф одиннадцатый. Сложные лекарства
- Образование условных рефлексов на сложное раздражение.
- Дискуссия! Является ли человеческийорганизм более сложным и совершенным, чем ПК?
- Адаптивное управление разверткой видеокадра. Алгоритм управления
- Сложные ситуации и необходимые действия.
- Сложные методы окраски бактерий