ТЕРМИЧЕСКИЕ АДАПТАЦИИ У ГОМОЙОТЕРМНЫХ ОРГАНИЗМОВ
Физиологические механизмы терморегуляции гомойотермных организмов включают как изменения теплообразования, так и изменения теплоотдачи. В эти процессы вовлекаются почти все органы и системы и только в итоге направленного изменения функции каждой системы температура тела млекопитающих и птиц остается на постоянном уровне или в рамках закономерных физиологических колебаний.
Кроме того, большую роль в поддержании термического гомеостазиса играют реакции поведения как более простые — приближение или удаление от источников тепла или холода, смена температурной зоны (уход в тень, в нору, выход на солнце, скучивание особей (одного или разных видов), так и более 'сложные — постройка гнезд, нор, убежищ и т. д.
На примере изучения реакций животных при различной температуре среды в природных и экспериментальных условиях можно хорошо проследить взаимоотношения индивидуальных и видовых адаптаций и сопоставить лежащие в их основе физиологические механизмы.
Наиболее изучены адаптивные изменения реакций обмена веществ в организме млекопитающих и птиц на изменение температуры внешней среды. Эти реакции в физиологии уже давно получили наименование химической терморегуляции.
Кривую изменений газообмена у гомойотермного организма под влиянием различных температур среды можно представить себе следующим образом (рис. 35). При низких температурах среды, совместимых с жизнью данного вида, газообмен и теплопродукция повышены. При переходе к более высоким температурам теплопродукция
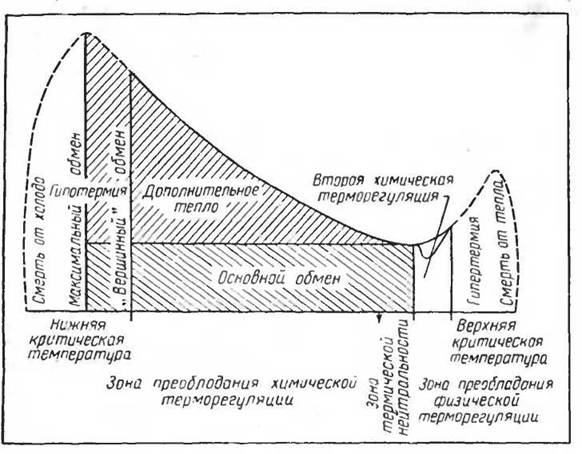
Рис. 35. Схема химической терморегуляции у гомойотермних организмов (по Gelineo, 1964)
понижается и при какой-то температуре (для разных видов млекопитающих от 5—6° С до 33—35° С) достигает наименьших величин. Эту температуру при наименьшем обмене веществ в покое обычно называют критической точкой.
С дальнейшим повышением температуры среды газообмен начинает повышаться как благодаря включению механизмов физической терморегуляции — полипноэ или обильного потоотделения, так и вследствие повышения температуры тканей тела при высокой температуре среды. Обычно у животных, имеющих хорошо выраженное полипноэ, критическая точка обмена веществ также хорошо выражена. У животных, не имеющих полипноэ (непарнокопытных, обезьян), и человека критическая точка выражена слабо и заменяется довольно обширной (до 15° С) зоной постоянного уровня обмена веществ. При дальнейшем повышении температуры среды и температуры тела газообмен начинает повышаться, но это обычно уже связано с явлениями перегревания. При очень низких температурах среды и наступающем охлаждении тела газообмен может снижаться, что сопровождается значительным снижением температуры тела и может знаменовать наступление гипотермии.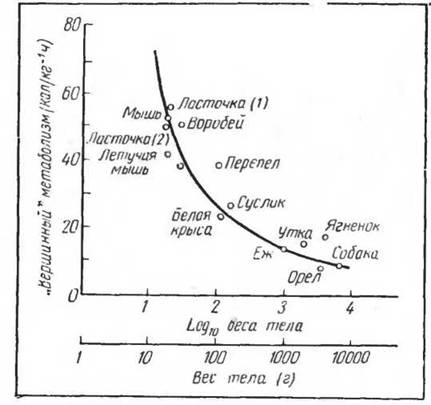
Рис. 36. «Вершинный» обмен (summit metabolism), отнесенный к весу тела у гомойотермних организмов (по Gelineo et Giaja, 1924, 1962)
Вместе с тем реакция обмена веществ на крайнее охлаждение довольно однотипна для различных видов млекопитающих и птиц. Наибольшее повышение обмена (metabolisme du sommet) (Giaja, 1934) продолжается различное время в связи с разной скоростью охлаждения ледяной водой (рис. 36) животных с различной поверхностью тела, теплоизоляцией и т. д.
Как правило, у животных, обитающих в условиях низких температур, критическая точка ниже, чем у обитателей мест с более высокой температурой (рис. 37). Снижение критической точки наблюдается у водных млекопитающих— нутрии (Слоним, 1952), ондатры (Щеглова, 1954).
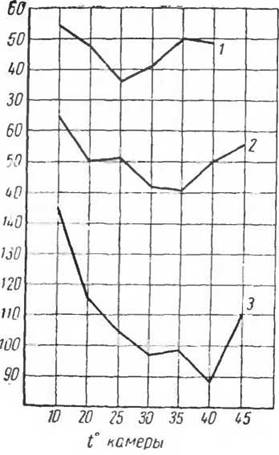
Рис. 37. Химическая терморегуляция {Qi!мл!мин) и критическая точка у млекопитающих (по Слониму, 1952):
I — собака. 2 — шакал. 3 — динго
Между устойчивостью организма к высокой или низкой температуре среды и систематическим положением животного данного вида едва ли существует какое-либо соответствие.
В любом климатическом поясе встречаются представители многих отрядов млекопитающих и только отдельные отряды приспособлены в своем распространении к узким условиям существования, например обезьяны, полуобезьяны п др. к субтропическому или тропическому климату.У разных животных химическая терморегуляция развита различно. Можно сослаться на общую формулу, указывающую, что по мере восхождения по эволюционной лестнице роль изменений теплопродукции при разных температурах среды— химической терморегуляции— падает. Так, у ежей можно наблюдать 3—4-кратное повышение обмена при низких температурах (5— 6° С) по сравнению с обменом при температуре 25° С. У более высокостоящих грызунов— двукратное повышение обмена, а у хищников, в частности собак, — только в 0,7—0,8 раза. Это находит свое отражение в увеличении нормы питания лисиц и собак, содержащихся в питомниках в зимнее время. У обезьян и человека химическая терморегуляция имеет небольшое значение. Особенно это касается человека, мало тренированного к воздействию низкой температуры среды. У разных видов обезьян эти явления выражены в зависимости от того, какие климатические районы они населяют: если это животные тропического пояса, то химическая терморегуляция приближается по интенсивности к таковой у человека.
Первое исследование, характеризовавшее особенности популяций, взятых из разных широт (Руттенбург, 1950), показало, что в зоне температур от 0 до 35° С популяции серой крысы (Rcittas norvegicus) из Мурманска, Ленинграда, Севастополя и Сухими резко отличаются и по интенсивности химической терморегуляции и по критической точке обмена веществ.
С другой стороны, популяции черной крысы из Мурманска и Севастополя, а также александрийской крысы из Ленинграда и Севастополя совершенно не отличались. Этот весьма важный факт указывает на малую вариа- бильность популяционных адаптаций у подвидов с более узким географическим распространением — черной и александрийской крысы — по сравнению с широко распространенной под всеми широтами серой крысой.
Южные формы (черная и александрийская), заносимые на север морскими судами, образуют популяции в отапливаемых зданиях, кухнях и т. д. и, таким образом, холод и отсутствие способности к адаптации ограничивают их широкое распространение в условиях Севера.
Уровень обмена и химическая терморегуляция значительно отличались у двух популяций желтогорлых мышей (Apodemus flavicollis), выловленных под Харьковом и в Крыму (Калабухов и Ладыгина, 1953). У крымской популяции наблюдалось более низкое потребление кислорода при температуре от 0 до 25° С.
Географические отличия в теплообмене сказываются очень ярко даже у популяций полевок Microtus arvalis, разделенных с севера на юг расстоянием около 100 км. Северная популяция полевок обладала более высоким обменом, чем южная.
Между интенсивностью химической терморегуляции и устойчивостью организма к холоду нет прямой зависимости. Например,- интенсивность терморегуляции у крысы выше, чем у кролика (Gelineo, 1948); вместе с тем общая устойчивость организма кролика к холоду выше, чем у крысы. Так, кролик способен поддерживать химическую терморегуляцию до —45° С, тогда как крыса только до — 20° С. Это различие зависит от величины теплоотдачи с поверхности тела кролика и крысы, которая у крысы, конечно, оказывается значительно большей на единицу массы тела.
Повышение интенсивности обмена и химической терморегуляции при переходе с юга на север не распространяется на организмы, живущие за Полярным кругом. Полярные животные, с трудом добывающие корм в условиях зимы, не могли бы существовать, если бы так резко увеличивался их обмен веществ при низких температурах. В зимних условиях на побережье Ледовитого океана песцы, например, не добывают двойного количества пищи, а довольствуются питанием, уменьшенным по сравнению с летним. Отсюда исключительно теплый мех — приспособление, которое ограничивает повышение обмена веществ при действии холода. Лучше всего выражена химическая терморегуляция у животных, обитающих в средних широтах в континентальном климате, а в условиях тропиков или в условиях Крайнего Севера она развита слабо.
Большую роль в интенсивности реакций обмена веществ на охлаждение играет теплоизоляция кожных покровов (мех и подкожный жировой слой). Эти особенности могут резко уменьшать реакцию на охлаждение. Это имеет место у животных в зимнем мехе или, например, у полярных видов (Scholander, Hock, Walters а. Irving, 1950). Однако только этот фактор (см. ниже) не может объяснить своеобразия химической терморегуляции ни в зимний период, ни в условиях Арктики. Но, несмотря на то, что отдельные виды обладают характерными чертами приспособления к определенным климатическим областям, некоторые нз них имеют очень обширное географическое распространение, не ограниченное определенной климатической зоной. Подобная особенность — способность к существованию в разных температурных условиях—получила в экологии название эвритермнн. Эвритермпыми организмами среди млекопитающих и птиц являются многие грызуны (серая крыса, серый хомячок), красная лисица, заяц-русак, волк, домовый воробей, серая ворона и др. (Hesse, 1924). В основе эврн- термин лежит способность животного приспосабливаться к измененным температурным условиям — способность к быстрой и медленной стойкой акклиматизации.
Особенно ярко выражены эти особенности терморегуляции у животных, приспособленных к континентально- ISO
му климату, где имеются огромные суточные перепады температуры (суточная амплитуда колебаний температуры в условиях Казахстана 30—32° С). Эти значительные перепады приводят к формированию соответствующих реакций терморегуляции. Животные, которые здесь обитают (лисица-караганка, степной хорек и др.), показывают очень хорошую картину терморегуляции и приспособлены к любым температурам внешней среды благодаря чрезвычайно подвижной реакции обмена веществ — химической терморегуляции н полипноэ. Эти реакции обмена и их подвижность осуществляются за счет чрезвычайно быстрого образования условных рефлексов, их быстрого дифференцирования и угасания (Ольнянская и Слоним, 1947; Исаакян, 1949; Слоним, 1952).
Географические различия в химической терморегуляции у птиц изучены очень мало. Наблюдаются значительные различия критической точки у различных видов гусей, гнездящихся в Северном полушарии (Якубанис, 1967).
Влиянию индивидуальной адаптации на химическую терморегуляцию посвящено большое количество исследований. Классическими многолетними исследованиями (Gelineo, 1934а, б, 1936, 1948) было показано, что как интенсивность химической терморегуляции, так и температура критической точки изменяются под влиянием длительного (3—4-недельного) содержания при разных температурах внешней среды. Джелнпео назвал эти сдвиги теплопродукции, изученные им у птиц, белых крыс и сусликов, адаптационными изменениями обмена веществ и терморегуляции. По Джелннео, величина теплопродукции складывается из двух компонентов: а) уровня обмена веществ, который при всех температурах среды после длительного пребывания в условиях низких температур повышается, а после пребывания при высоких, наоборот, падает; б) химической терморегуляции, которая после пребывания в условиях низких температур возрастает, а после пребывания при высоких — падает. Эти отношения хорошо видны на рисунке 38, где приведены данные для зеленушки (Chrysomitris spinus) после содержания ее при температурах среды —2°—8° С; 15°—20,5° С и 29°— ( + 32°) С. На этом рисунке видно, что критическая точка сдвигается после адаптации к более высоким температурам с 30° С к 34° С. Данные адаптационных изменений
газообмена у крыс приведены в таблице 9. Здесь видно, что изменяется нс только нижняя, но и верхняя критическая точка (температуры); животные становятся намного устойчивее к гипотермии после содержания при температуре О 2° С.
В настоящее время твердо установлено, что индивидуальная адаптация к холоду связана у гомойотермных
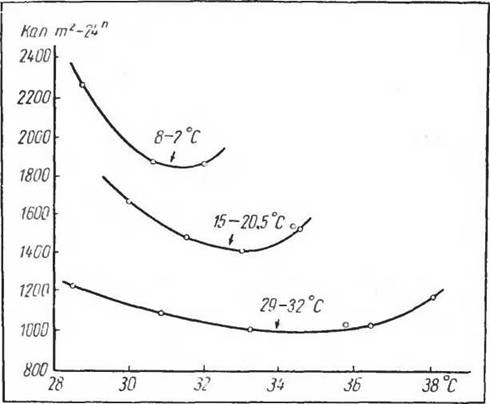
Рис. 38 Химическая терморегуляция при адаптации зеленушек к высоким и низким температурам срезы (по Gelineo, 1934):
1 — при адаптации к температуре среды 8 2Р; 2 — при адаптации к температуре 15—20,5°; 3 — при адаптации к
температуре 29—.32 С. По оси ординат — теплопродукция (кал,'сутки); но оси абсцисс — температура среды
организмов прежде всего с увеличенной способностью поддержания температуры тела при охлаждении и временем выживания организма при воздействии очень низких температур среды (Hart, 1964а).
Обширные исследования посвящены в последние годы проблеме источников повышенного теплообразования в процессе адаптации к холоду. Наиболее ярким фактом является то, что животные при длительных или повторных охлаждениях поддерживают повышенный уровень обмена при одновременном снижении мышечной ак-
Адаптация к различным температурам и потребление кислорода у некоторых пойкилотермных организмов
Теплопродукция и ее особенности у крыс, адаптированных к трем различным температурам среды
(по Celineo, 1934)
| Температура адаптации | 30—32° с | 16-20° С | 0—2° С |
| Термическая нейтральность .................................. | 32-34.5 | 30—32 | 29—30 |
| Объем в покое (кг/кал/лі2/24 ч} ... . | 564 | 640 | 911 |
| Избыточное теплообразование на ГС (кг/кал/24 ч) | 29 | 40 | 55 |
| «Вершинный обмен» (кг/кал/м2}24 ч) ... . | 746 | 1817 | 3100 |
| Низшая критическая точка (°С)............................ | 24 | 2 | -10 |
| Температура, при которой обмен максимален (°С) | 2 | -10 | -10 |
| Максимальный уровень обмена (кг/кал/лі2/24 ч) | 1514 | 2313 | 3100 |
тивности, т. е. видимой дрожи или определяемой с помощью специальной аппаратуры высокой электрической активности скелетной мускулатуры (Heroux, 1962; Hart, 19646; Бисерова и Гембчински, 1964; Иванов, 1965). Так возникло представление о несократительном термогенезе, т. е. таком повышении обмена веществ, которое не связано с усиленной функцией скелетной мышцы как сокращающегося органа. Прямым доказательством существования несократительного термогенеза явилось влияние норадреналина на общий газообмен у адаптированных к холоду крыс и на газообмен мышц тех же крыс, изученный in situ (Hsieh, Carlson, Gray, 1957; Depocas, 1960; Heroux, 1962; Ткаченко, 1968). Норадреналин в данном случае не является основным веществом, поддерживающим высокий метаболизм в целом организме. Приведенные выше данные (Исаакян, Макарова, Щеглова, 1968; Jansky a. Hart, 1968) свидетельствуют о значительном повышении газообмена во внутренних органах, что также приводит к возрастанию роли несократительного термо-
генеза. Возрастает и кровоток во внутренних органах и жировой ткани (Jansky a. Hart, 1968).
Несократительный термогенез ярко выражен в раннем постпатальном периоде (исследования на морских свинках) и связывается в этом случае с развитием бурой жировой ткани в межлопаточной области (см. также гл. IV), затем он в нормальных условиях развития животного заменяется сократительным термогенезом, а при холодовой адаптации вновь проявляется (Bruck, u. Wiinnenberg, 1965, 1966; Moore a. Simmonds, 1966).
Сократительный термогенез и его распределение в организме лучше всего может быть изучено при отведении игольчатыми электродами биотоков с разных групп мышц у животного, подвергающегося охлаждению (Слоним, 1964, 1966; Иванов, 1965; Баженов, 1968; Давыдов и Кузьмо, 1968 и др.). Как общую схему здесь можно принять, что наибольшую электрическую активность при охлаждении обнаруживают мышцы шеи, жевательные мышцы н отчасти мышцы спины и передних конечностей. Мышцы задних конечностей вовлекаются в терморегуля- циоиную активность в последнюю очередь при очень интенсивном охлаждении. В эту схему, характерную для всех млекопитающих, необходимо, однако, ввести поправки для водных организмов (нутрия, ондатра), у которых наибольшая реакция наблюдается в мышцах спины и конечностей, т. е. в мышцах, непосредственно подвергающихся охлаждению в воде (Давыдов и Кузьмо, 1968). У грызунов терморегуляционный тонус выражен вообще относительно слабо; он заменяется в жевательных мышцах активными мышечными сокращениями при охлаждении (рис. 39).
Такая же картина распределения мышечного терморегуляционного тонуса наблюдается у птиц. На рисунке 40 приведены данные, характеризующие электрическую активность мышц груди и мышц бедра у представителей воробьиных п водоплавающих. У последних мышцы бедра являются наиболее активными в отношении сократительного теплообразования.
В процессе адаптации наблюдаются некоторые изменения в химической динамике скечетной мускулатуры. Они касаются взаимоотношений между процессами дыхания и окислительного фосфорилирования в процессе теплообразования (так называемый коэффициент Р/О)
Рис. 39. Схема распределения терморегуляционного
тонуса у грызунов(по Иванову, 1964; Слони- му, 1964). /—белая мыть; //— крыса; /// — кролик: а—до охлаждения, б— после охлаждения
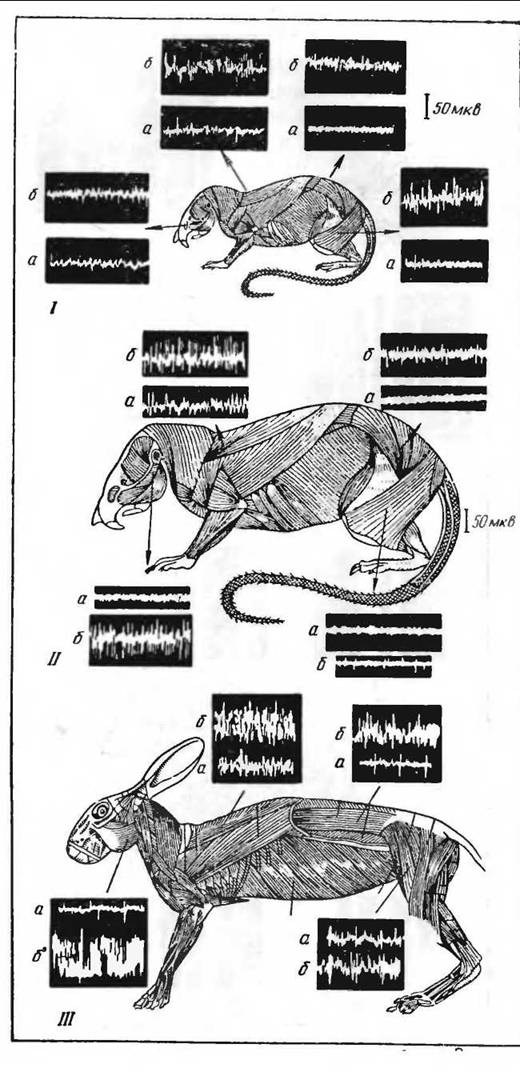
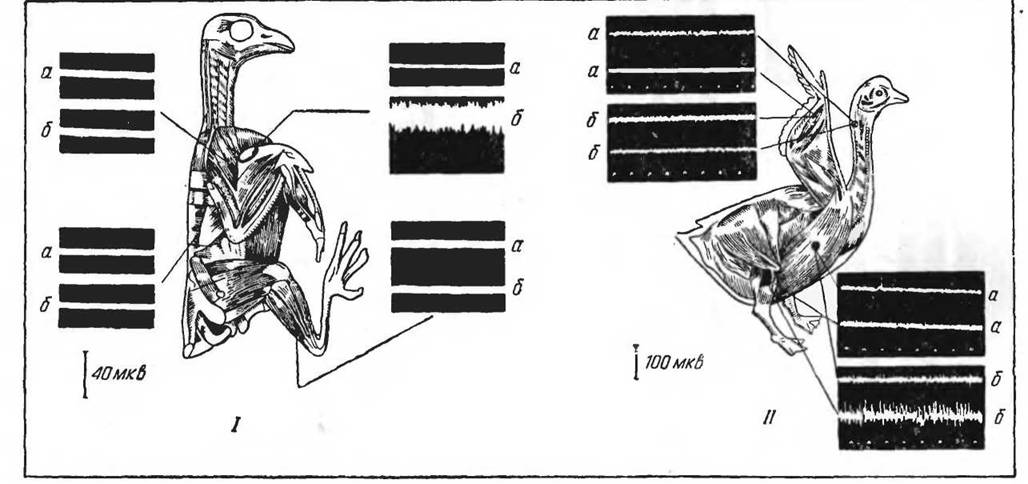
Рис. 40. Схема распределения терморегуляционного тонуса у птиц (по Кескпайку и Давыдову, 1966 и Якуба- иису, 1966),
/ — черный дрозд; II—домашний гусь: а — до охлаждения, б — после охлаждения (прп —10° С). Отметка времени 0,1 сек.
(Lianides a. Beyer, 1960; Smith 1960; Скулачев, 1964 и др.). Однако эти изменения доказаны в основном по отношению к митохондриям печени и не охватывают скелетной мускулатуры. Было показано в то же время, что инъекции норадреналина не повышают газообмена в печени адаптированных к холоду крыс (Jansky, 1965), оказывая это влияние на скелетную мускулатуру. В то же
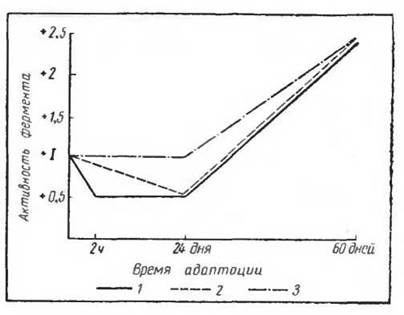
Рис. 41. Активность фермента гналурони- дазы в разных мышцах крыс в процессе адаптации к холоду (по Дерибас, 1967):
1 _ m. rectus femoris, 2 — in. sterno-cleido-иіа- stoideus, 3 — m. trapecius
время установлено изменение ферментативной сукцинде- гидрогеназной активности в процессе холодовой адаптации. Гистохимические исследования скелетной мускулатуры крыс, адаптированных к холоду (Дерибас и сотр., 1967), показали, что в мышцах конечностей можно наблюдать увеличение числа мышечных волокон, содержащих большое количество фермента. Характерно, что эти сдвиги наблюдались в большей мере в мышцах конечностей, нежели в мышцах шеи и жевательных (рис. 41). Таким образом, в процессе адаптации происходит вовлечение в терморегуляторную активность (несократительную) именно той части мускулатуры, которая не является активной в обычных условиях теплообмена.
В настоящее время есть основания полагать, что эти данные, добытые в основном на крысах, могут быть пе- ренессны на большинство млекопитающих, но не на птиц, где явления несократительного термогенеза до настоящего времени не обнаружены. В то же время у обезьян Saitniri sciurea при адаптации к холоду обнаружены изменения общего газообмена, окислительных ферментов в тканях, скорости окисления а-глицероф.осфата в бурой жировой ткани, митохондриях печени, совершенно аналогичные тем, которые описаны для лабораторных крыс (Chaffee, Allen, Brewer, Horvath, Mason a. Smith, 1966).
Изменения основного обмена (измеренного при определенной температуре) при адаптации к холоду представляются в следующем виде. В течение первых пяти дней у крыс наблюдается значительное повышение газообмена, затем газообмен продолжает медленно повышаться до 15—20-го дня, адаптация несколько снижается после 44-го дня к 60-му (рис. 42) (Cottle a. Carlson, 1954; Щеглова, 1967). Однако совершенно иные картины адаптационных изменений газообмена были получены при длительном содержании с температурой около 5° С представителей субтропических видов грызунов — индийской крысы незокнп (Nesokia indica) и большой песчанки (Rhombomis opitnus) (рис. 43). Оказалось, что грызуны— обитатели субтропиков и тропиков при температуре около 5° С не обнаруживают никаких адаптационных изменений обмена веществ. Температура тела остается пониженной и не поднимается в процессе адаптации, как это имеет место у крыс. Следовательно, изменения газообмена являются весьма важными звеньями в процессе термической адаптации и их отсутствие указывает на невозможность образования адаптированных к изменившимся условиям популяций.
В противоположность адаптации к холоду содержание этих животных в тепле приводит к стойкому снижению обмена, более выраженному, чем у крыс. Отсюда понятна возможность расселения этих организмов далеко на юг и возможность образования стойких популяций с изменением обмена веществ и химической терморегуляции (рис. 43, 44).
Следовательно, адаптационные изменения газообмена к холоду образуются только в том случае, если температурные условия среды не ведут к значительным нарушениям терморегуляции, например к значительному понижению температуры тела. Прп склонности многих мыше- 18«
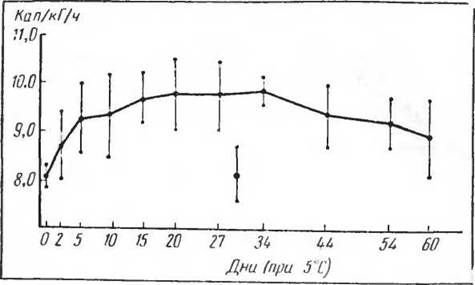
Рис. 42. Теплопродукция у белой крысы в процессе адаптации к холоду (по Cottle a. Carlson, 1954)
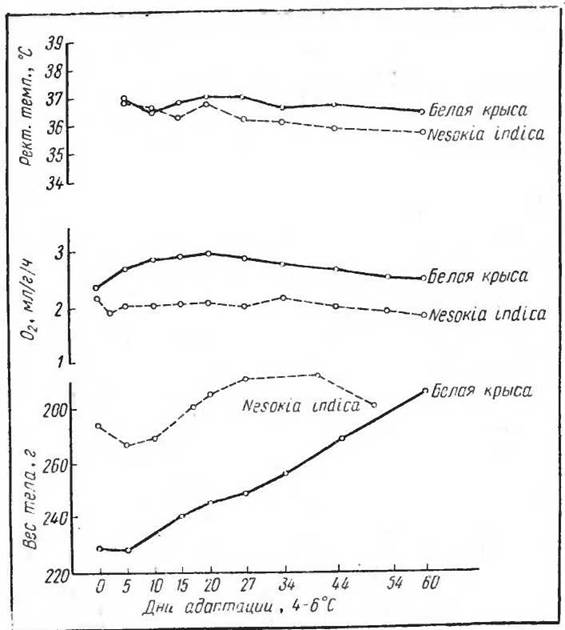
Рис. 43. Изменение веса тела, потребления О2 (млігіч) и ректальной температуры у лабораторной крысы и тропической крысы незокии (по Исаакян, Макаровой, Щегловой, 1969)
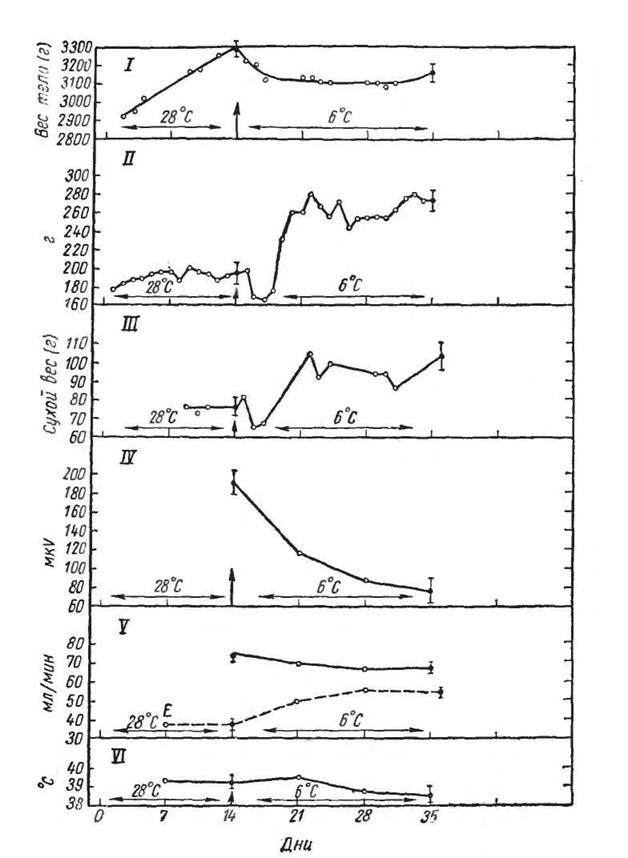
Рис. 44. Динамика физиологических функций у остриженных кроликов во время двухнедельной адаптации к 28° С и последующей 3-недельной адаптации к 6° С. I—вес тела (г); II— потребление пиши (г/декь); III — образование кала (г/день); IV — электрическая активность мышц (mkV/ЗО мин); V — потребление кислорода при 28°С (млІминІЗХ) мин)’, VI — температура тела (°С) (по Нёгоих, 1966)
видных грызунов (полевок) снижать уровень температуры тела и обмена при действии низкой температуры адаптационные изменения обмена полностью отсутствуют. Так, у содержавшихся в холоде пеструшек обмен под влиянием охлаждения оказывался ниже, чем у тех же пеструшек, содержавшихся в тепле (Гладкина, Мейер, Мокеева, 1958). Вероятно, только благодаря адаптационным изменениям теплообмена некоторые исследователи обнаруживают крайнюю нестойкость эколого-физиологических изменений, связанных с термическими условиями среды обитания.
Еще по теме ТЕРМИЧЕСКИЕ АДАПТАЦИИ У ГОМОЙОТЕРМНЫХ ОРГАНИЗМОВ:
- 1. Представления о процессе адаптации организма.
- Термический ожог
- Термическая чувствительность.
- Термические ожоги трахеи
- 2.6.2. Термические ожоги
- 180. Термические ожоги глаз
- Действие термических факторов
- Термические ожоги
- 27. Профессиональная адаптация. Причины трудностей профессиональной адаптации
- Лекция 12. Тема: «Помощь пострадавшим с термическими поражениями»
- Современные методы реанимационной терапии тяжелых химических и термических ожогов глаз
- №3 Организм человека как единая биологическая система. Уровни организации человеческого организма: молекулярный, клеточный, тканевой, органный, системный, организменный.
- 7.2.2. Нарушения со стороны пищевода, связанные с механическими и термическими воздействиями