Нейромышечное соединение
Ацетилхолин является нейротрансмиттером в синапсе между моторным нейроном и клеткой мускулатуры, который часто называют нервно-мышечным (или нейромышечным] соединением (neuromuscular junction^ или нейромускулярным (или нервно-мышечным) синапсом.
Нейрон может образовывать несколько сотен синапсов с одной мышечной клеткой.Приход потенциала действия в пресинаптическую часть моторного нейрона открывает Са2+-каналы (рисунок 154(A)), а ионы кальция стимулируют экзоцитоз ацетилхолина.
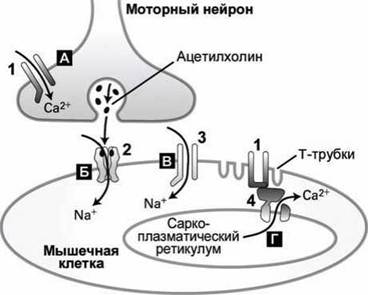
Рисунок 154 - Последовательная активизация ионных каналов в синапсе нейромышечного соединения: 1 - потенциалочувствительные кальциевые каналы: 2 - никотиновый ацетилхолиновый рецептор: 3 - потенциалочувствительный натриевый канал: 4 - Са2+-канал саркоплазматического ретикулума
Чувствительность к ацетилхолину обеспечивается холинорецепто- рами, среди которых различают два основных типа: мускариновый (в гладких мышцах и мозге) и никотиновый (в скелетных мышцах и вегетативных ганглиях).
Никотиновый ацетилхолиновый рецептор, который экспрессируется в клетках мускулатуры, является лиганд-чувствительным ионным каналом, который пропускает и Na+, и К+. В открытом состоянии такой канал способен пропускать 15-30 тысяч ионов Na+ или К+ за миллисекунду (или 15-30-106 ионов в секунду).
Однако, поскольку потенциал покоя мышечной клетки близок к равновесному потенциалу для ионов калия, то открытие канала в ацетилхолиновом рецепторе практически не стимулирует перенос ионов калия внутрь клетки, а ионы натрия, напротив, перемещаются по такому каналу внутрь клетки под действием как электрического поля (в состоянии покоя цитозоль клетки заряжен отрицательно), так и градиента концентрации (рисунок 154(B)).
Одновременное увеличение проницаемости постсинаптической мембраны для ионов Na+ или К+ вследствие действия ацетилхолина на рецептор приводит к деполяризации мембраны до -15 мВ от потенциала покоя около -90 мВ. Такая локальная деполяризация стимулирует открытие потенциалочувствительных Ыа+-каналов (рисунок 154(B)), что в свою очередь индуцирует генерацию и распространение потенциала действия на поверхности мышечной клетки по тому же механизму, который был описан выше для нейронов.
Когда деполяризация мембраны доходит до Т-трубок (специализированных впячиваний плазматической мембраны), то потенциалочувствительные Са2+-каналы в этой области мембраны изменяют конформацию, что вызывает также конформационные переходы и в связанных с ними рианодиновых Са2+-каналах (рисунок 141) на поверхности саркоплазматического ретикулума клетки (рисунок 154(Г)). За этим следует выход ионов Са2+ из саркоплазматического ретикулума в цитозоль, что и стимулирует сокращение мышечной клетки.
16.4.
Еще по теме Нейромышечное соединение:
- Мониторинг нейромышечного блока.
- Электромиографическая оценка нейромышечной координации жевательных мышц
- Фторорганические соединения.
- Щелевые соединения
- Типы соединения костей.
- Экстрасистолы из A-В соединения:
- Б. Фосфорорганические соединения: повышение их избирательности.
- Ассоциативность и коммутативность соединения
- Фосфорорганические соединения и карбаматы
- А. Фосфорорганические соединения: химизм действия.
- 209. Поражения мышьяком и его соединениями
- Мембранотропные соединения как индукторы автолиза
- I. Серосодержащие соединения
- 206. Поражения соединениями хрома