ю. тНЯТНПНПЦЮМХВЕЯЙХЕ ЯНЕДХМЕМХЪ: УХЛХГЛ ДЕИЯРБХЪ.
еЫЕ
Б ЯЮЛНЛ МЮВЮКЕ ХГСВЕМХЪ ЩРХУ ЯНЕДХМЕМХИ АШКН ГЮЛЕВЕМН, ВРН С КХЖ, ПЮАНРЮЧЫХУ Я ЯНЕДХМЕМХЪЛХ РХОЮ ДХТКНЯЮ (13.26), БНГЛНФ╜МШ НДШЬЙЮ, ОНБШЬЕММЮЪ ВСБЯРБХРЕКЭМНЯРЭ Й ЯБЕРС, БПЕЛЕММЮЪ ОНРЕПЪ ЯНГМЮМХЪ Х НАПЮРХЛЮЪ ОНРЕПЪ ГПЕМХЪ [Lange, Kreuger, 1932; Schrader, 1963].
сЙЮГЮММНЕ ДЕИЯРБХЕ ТНЯТНПНПЦЮМХВЕЯЙХУ ЯНЕДХМЕМХИ ОПЕДОНКНФХРЕКЭМН ЯБЪГШБЮКХ Я ХМЦХАХПНБЮМХЕЛ юущ[10]. хГСВЕМХЕ ТНЯТНПХКХПНБЮМХЪ ТЕПЛЕМРЮ Я ОПХЛЕМЕМХЕЛ ПЮДХНЮЙРХБМНЦН ХГНРНОЮ 32п, ХМРЕМЯХБМНЦН ХЯРНВМХЙЮ ^-ВЮЯРХЖ Я ОЕПХНДНЛ ОНКСПЮЯОЮДЮ 14 ДМЕИ, ОНЙЮГЮКН, ВРН ДХТКНЯ (0,0-ДХХ- ГНОПНОХКТРНПТНЯТЮР) (13.26) ЮЖХКХПСЕР ЦХДПНЙЯХЦПСООС НЯРЮР╜ЙЮ ЯЕПХМЮ Б ТЕПЛЕМРЕ, ЪБКЪЧЫСЧЯЪ ЙЮРЮКХРХВЕЯЙХЛ ЯЮИРНЛ ДКЪ юу. оПХ ЩРНЛ ОПНХЯУНДХР НРЫЕОКЕМХЕ ТРНПХЯРНЦН БНДНПНДЮ [Schaffer, May, Summerson, 1954]. оПХ ЦХДПНКХГЕ ТНЯТНПХКХПН- БЮММНЦН ТЕПЛЕМРЮ БШДЕКЪЕРЯЪ н,н-ДХХГНОПНОХКТНЯТНПХКЯЕПХМ.дХТКНЯ ХЯОНКЭГНБЮКХ ДКЪ ЯМХФЕМХЪ БМСРПХЦКЮГМНЦН ДЮБКЕМХЪ ОПХ ЦКЮСЙНЛЕ, МН Б МЮЯРНЪЫЕЕ БПЕЛЪ ДКЪ ЩРНИ ЖЕКХ ЯНГДЮМШ МН╜БШЕ, ЛЕМЕЕ РНЙЯХВМШЕ КЕЙЮПЯРБЕММШЕ ОПЕОЮПЮРШ.
дЕИЯРБХЕ НПЦЮМХВЕЯЙХУ ТНЯТЮРНБ ХГСВЮКХ РЮЙФЕ МЮ ОПХЛЕПЕ ОЮПЮНЙЯНМЮ (13.25), ОПЕДЯРЮБКЪЧЫЕЦН ЯНАНИ ЮЙРХБМСЧ ТНПЛС ОПЕОЮПЮРЮ ОЮПЮРХНМЮ (13.24) [Gage, 1953]. оЮПЮНЙЯНМ ОНДНАМН ДХТКНЯС ТНЯТНПХКХПСЕР ЦХДПНЙЯХЦПСООС ЯЕПХМЮ, ОПХ ЩРНЛ БШ╜ДЕКЪЕРЯЪ ОЮПЮ-МХРПНТЕМНК [Hartley, Kilby, 1952]. мЮ НЯМНБЮМХХ ЩРХУ ХЯЯКЕДНБЮМХИ АШК ЯДЕКЮМ БШБНД, ВРН Б юущ ЯСЫЕЯРБСЕР РНКЭЙН НДХМ ЙЮРЮКХРХВЕЯЙХИ ЮЙРХБМШИ СВЮЯРНЙ≈∙ ЦХДПНЙЯХЦПСООЮ ЯЕПХМЮ. нДМЮЙН ЯСАЯРПЮРС юущ ПЮБМН МЕНАУНДХЛ БРНПНИ ЮЙРХБ╜МШИ СВЮЯРНЙ, ЯБЪГШБЮМХЕ ЯСАЯРПЮРЮ Я ЙНРНПШЛ БШГШБЮЕР ЙНМТНП╜ЛЮЖХНММШЕ ХГЛЕМЕМХЪ ТЕПЛЕМРЮ, ЯОНЯНАЯРБСЧЫХЕ ЙЮРЮКХРХВЕ╜ЯЙНЛС ДЕИЯРБХЧ ЮЙРХБМНЦН ЖЕМРПЮ [O▓Brien, 1971]. оПЕДОНКЮЦЮ╜ЧР, ВРН ЕЯРЭ РПХ РХОЮ РЮЙХУ ╚БРНПХВМШУ╩ ЮЙРХБМШУ ЖЕМРПНБ: ОЕПБШИ ≈ ДКЪ юущ Х РЕРПЮЩРХКЮЛЛНМХЕБШУ ЯНКЕИ, БРНПНИ ≈ ДКЪ ТНЯТНПНПЦЮМХВЕЯЙХУ ЯНЕДХМЕМХИ Х РПЕРХИ ≈ДКЪ ЙЮПАЮЛЮРНБ. рЮЙХЛ НАПЮГНЛ, ╚БРНПХВМШИ╩ ЮЙРХБМШИ ЖЕМРП (ЯЛ.
ЛНДЕКЭ ПЕЖЕО╜РНПЮ МЮ ПХЯ. 12.3) ЪБКЪЕРЯЪ МЕ ЮМХНММШЛ, Ю КХОНТХКЭМШЛ.йХМЕРХВЕЯЙХЕ ХЯЯКЕДНБЮМХЪ ОНЙЮГЮКХ, ВРН ТНЯТНПНПЦЮМХВЕЯ╜ЙХЕ ЯНЕДХМЕМХЪ Х ТЕПЛЕМР ЯМЮВЮКЮ НАПЮГСЧР ОЕПЕУНДМШИ ЙНЛ╜ОКЕЙЯ, УЮПЮЙРЕПХГСЧЫХИЯЪ ЙНМЯРЮМРНИ ЮТТХММНЯРХ й [Aldridge, 1950; Main, 1964]. б ПЕГСКЭРЮРЕ ТНЯТНПХКХПНБЮМХЪ ТЕПЛЕМРЮ ЩРНР ЙНЛОКЕЙЯ ПЮЯОЮДЮЕРЯЪ Х БШРЮКЙХБЮЕР СУНДЪЫСЧ ЦПСООС. уЮПЮЙРЕПХЯРХЙНИ ЩРНЦН ОПЮЙРХВЕЯЙХ МЕНАПЮРХЛНЦН ОПНЖЕЯЯЮ ЯКС╜ФХР ЙП ≈ ЙНМЯРЮМРЮ ТНЯТНПХКХПНБЮМХЪ (ХКХ ХМЮЙРХБЮЖХХ). гМЮВЕМХЪ й Х ЙП НОПЕДЕКЕМШ ДКЪ ЛМНЦХУ ТНЯТНПНПЦЮМХВЕЯЙХУ ЯНЕДХМЕМХИ Х юущ, БШДЕКЕММШУ ХГ ПЮГМШУ ХЯРНВМХЙНБ.
щРХ ЙХМЕРХВЕЯЙХЕ ХЯЯКЕДНБЮМХЪ ОНЯКСФХКХ НЯМНБНИ ДКЪ ХГС╜ВЕМХЪ ЯБЪГХ ЯРПСЙРСПЮ ≈ ЮЙРХБМНЯРЭ. мЮОПХЛЕП, ДКЪ ПЪДЮ ЮМЮ╜КНЦНБ ОЮПЮНЙЯНМЮ (13.25) ОПХ ХГЛЕМЕМХХ ЮКЙНЙЯХЦПСОО (НР ЛЕ╜РНЙЯХ ДН АСРНЙЯХ) Х БЮПЭХПНБЮМХХ ЩКЕЙРПНМНЮЙЖЕОРНПМШУ ГЮЛЕЯРХРЕКЕИ Б АЕМГНКЭМНЛ ЙНКЭЖЕ АШКН СЯРЮМНБКЕМН, ВРН МХ ЮКЙНЙЯХЦПСООШ, МХ ОПХПНДЮ СУНДЪЫЕИ ЦПСООШ МЕ БКХЪЧР МЮ БЕКХВХМС ЙП. щРН ЯОПЮБЕДКХБН Х Б ЯКСВЮЕ ХГЛЕМЕМХЪ ВЕРШПЕУ ЮКЙХКЭМШУ ГЮЛЕЯРХРЕКЕИ Б ЛНКЕЙСКЕ ЛЮКЮНЙЯНМЮ (13.27). нДМЮ╜ЙН ГМЮВЕМХЪ й ДКЪ ОПНХГБНДМШУ ОЮПЮНЙЯНМЮ Б 2≈10 ПЮГ БШЬЕ, ВЕЛ ДКЪ ЛЮКЮНЙЯНМЮ. оН-БХДХЛНЛС, БШЬЕСЙЮГЮММШЕ ХГЛЕМЕМХЪ УХЛХВЕЯЙНИ ЯРПСЙРСПШ ЛНЦСР ХГЛЕМЪРЭ ЯПНДЯРБН Й ТЕПЛЕМРС, МН МЕ БКХЪРЭ МЮ ТНЯТНПХКХПСЧЫСЧ ЮЙРХБМНЯРЭ [Chiu, Main, Dau- terman, 1969]. оПХЯСРЯРБХЕ Б ЛНКЕЙСКЕ ЩКЕЙРПНМНЮЙЖЕОРНПМШУ ГЮЛЕЯРХРЕКЕИ Б СУНДЪЫЕИ ЦПСООЕ ЯОНЯНАЯРБСЕР ОНБШЬЕМХЧ ЮМРХ- УНКХМЩЯРЕПЮГМНИ ЮЙРХБМНЯРХ ДН НОПЕДЕКЕММНЦН ЛЮЙЯХЛСЛЮ, НДМЮ╜ЙН ГМЮВХРЕКЭМНЕ СЛЕМЭЬЕМХЕ ОПНВМНЯРХ ЯБЪГХ СУНДЪЫЕИ ЦПСООШ Я НЯРЮКЭМНИ ВЮЯРЭЧ ЛНКЕЙСКШ ОПХБНДХР Й НАПЮГНБЮМХЧ ЯНЕДХ╜МЕМХИ, КЕЦЙН ЦХДПНКХГСЧЫХУЯЪ ОНД ДЕИЯРБХЕЛ БНДШ [Aldridge, Barnes, 1952; Aldridge, Davison, 1952]. мХФЕ ЩРНЦН ЛЮЙЯХЛСЛЮ ЯСЫЕЯРБСЕР КХМЕИМЮЪ ГЮБХЯХЛНЯРЭ ЛЕФДС КНЦЮПХТЛНЛ ЙНМЯРЮМ╜РШ ЦХДПНКХГЮ Х КНЦЮПХТЛНЛ ЙНМЯРЮМРШ ЯЙНПНЯРХ АХЛНКЕЙСКЪП-
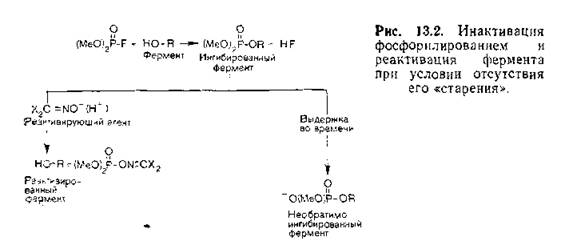
МНИ ПЕЮЙЖХХ БГЮХЛНДЕИЯРБХЪ ТНЯТНПНПЦЮМХВЕЯЙНЦН ЯНЕДХМЕМХЪ Я ТЕПЛЕМРНЛ.
щТТЕЙРХБМШЕ ХМЯЕЙРХЖХДШ МЮИДЕМШ ЯПЕДХ ЩТХПНБ ТНЯТНП╜МНИ, ТНЯТХМНБНИ Х ТНЯТНМНБНИ ЙХЯКНР.
б ЩТХПЮУ ТНЯТХМНБНИ ЙХЯКНРШ НАЕ ЮКЙХКЭМШЕ ЦПСООШ ОПХЯНЕДХМЕМШ МЕОНЯПЕДЯРБЕММН Й ЮРНЛС ТНЯТНПЮ, Б ЩТХПЮУ ТНЯТНПМНИ ЙХЯКНРШ НАЕ ОПХЯНЕДХ╜МЕМШ ВЕПЕГ ЙХЯКНПНД; ЩТХПШ ТНЯТНМНБНИ ЙХЯКНРШ ГЮМХЛЮЧР ОПНЛЕФСРНВМНЕ ОНКНФЕМХЕ.мЮ ПХЯ. 13.2 ХГНАПЮФЕМШ НЯМНБМШЕ ОПНЖЕЯЯШ, ОПНХЯУНДЪЫХЕ ОПХ ТНЯТНПХКХПНБЮМХХ ТЕПЛЕМРЮ. еЯКХ ЕЫЕ МЕ ОПНХГНЬКН ╚ЯРЮ╜ПЕМХЪ╩ ХМЦХАХПСЕЛНЦН ТЕПЛЕМРЮ, РН НМ ЛНФЕР АШРЭ ПЕЮЙРХБХПН╜БЮМ Б ПЕГСКЭРЮРЕ БГЮХЛНДЕИЯРБХЪ Я ЮМХНММНИ ЦПСООНИ ЦХДПН- ЙЯХКЮЛХМЮ. ъБКЕМХЕ ╚ЯРЮПЕМХЪ╩ АШКН НРЙПШРН Hobbiger (1955), НАМЮПСФХБЬХЛ, ВРН ЯКНФМШЕ ЩТХПШ БРНПХВМШУ ЯОХПРНБ ЯОНЯНА╜ЯРБСЧР ╚ЯРЮПЕМХЧ╩ Б АНКЭЬЕИ ЯРЕОЕМХ, ВЕЛ ЯННРБЕРЯРБСЧЫХЕ ЩТХПШ ОЕПБХВМШУ ЯОХПРНБ. оПЮБХКЭМНЕ ОПЕДЯРЮБКЕМХЕ Н ╚ЯРЮПЕ╜МХХ╩, ЙЮЙ Н ОПНЖЕЯЯЕ, ЯЙНПНЯРЭ ЙНРНПНЦН ОПНОНПЖХНМЮКЭМЮ ЯЙНПН╜ЯРХ ЦХДПНКХГЮ ЮКЙНЙЯХЦПСООШ, ОПЕБПЮЫЮЧЫЕИЯЪ ОПХ ЩРНЛ Б ЮМХНМ, ОНЪБХКНЯЭ ОНГДМЕЕ [Berends et al., 1959]. мЕВСБЯРБХРЕКЭ╜МНЯРЭ ХМЮЙРХБХПНБЮММНЦН, ╚ОНЯРЮПЕБЬЕЦН╩ ТЕПЛЕМРЮ Й ДЕИЯРБХЧ ЮМРХДНРЮ НАСЯКНБКЕМЮ ЯХКЮЛХ ЙСКНМНБЯЙНЦН НРРЮКЙХБЮМХЪ.
б НОШРЮУ МЮ МЮЯЕЙНЛШУ АШКН ОНЙЮГЮМН, ВРН ОНД ДЕИЯРБХЕЛ ТНЯТНПНПЦЮМХВЕЯЙХУ ЯНЕДХМЕМХИ Б ОЕПБШИ ВЮЯ ЯХКЭМН ХМЦХАХ╜ПСЕРЯЪ юущ Х ПЕГЙН БНГПЮЯРЮЕР СПНБЕМЭ ЯБНАНДМНЦН юу [Small- man, Fiske, 1958]. н ДЕИЯРБХХ ЩРХУ ЮЦЕМРНБ МЮ МЕПБМСЧ ЯХЯРЕЛС МЮЯЕЙНЛШУ ЯЛ. ПЮГД. 6.4.1.
юЖЕРХКУНКХМЩЯРЕПЮГШ, БШДЕКЕММШЕ ХГ ПЮГКХВМШУ БХДНБ МЮ╜ЯЕЙНЛШУ, ГМЮВХРЕКЭМН НРКХВЮЧРЯЪ ОН ВСБЯРБХРЕКЭМНЯРХ Й ТНЯТНП╜НПЦЮМХВЕЯЙХЛ ХМЯЕЙРХЖХДЮЛ [R. O▓Brien, 1967]. юущ С МЮЯЕЙНЛШУ ЛЕМЕЕ ДНЯРСОМЮ ДКЪ ЩЙГНЦЕММШУ ЯНЕДХМЕМХИ, ВЕЛ С ЛКЕЙНОХРЮ╜ЧЫХУ, ОНЩРНЛС ХНМХГХПНБЮММШЕ ЮМЮКНЦХ МЕ ЛНЦСР АШРЭ ХЯОНКЭ╜ГНБЮМШ Б ЙЮВЕЯРБЕ ХМЯЕЙРХЖХДНБ, РЮЙ ЙЮЙ ВСФЕПНДМШЕ ХНМШ МЕ ЛНЦСР ОПНМХЙЮРЭ ВЕПЕГ ЛЕЛАПЮМС (ПЮГД. 3.2).