Эволюционная роль Alu-элементов.
Среди других семейств ретроэлементов, Alu наиболее многочисленны в геноме человека. Древние Alu представлены более чем 1,4 млн копий, которые соответствуют 10 % всей массы генома.
Их число продолжает расти, и они встраиваются во все новые сайты с частотой примерно одно новое встраивание на 100-200 новорожденных (Аст Г., 2005).Происхождение первых Alu-мономеров, называемых также «окаменелыми» Alu-мономерами (fossil Alu monomers, FAMs), неизвестно; и их история уходит в глубину геологического времени. Они имели размер примерно в 160 кб. и в геноме человека представлены незначительно. «Современные*
Alu-эле менты появились не ранее чем 55 млн лет назад, в эпоху «до при- матов-антропоидов». Эти ретроэлементы представляют собой продукт слияния «голова к хвосту» двух различных FAM, которые дали начало димерной структуре, состоящей из двух сходных, но не идентичных мономеров (левое и правое плечо Alu), соединенных через А(аденин)-обога- щенный линкер. Транскрибированная с Alu-элементов Alu-PHK является высоко структурированной и поддерживает строгое структурное сходство со своей предковой РНК (Hasler J. и Strub К., 2006).
«Современные» Alu-повторы классифицируют в соответствии с их возрастом. AluJ-субсемейство было активно в период дивергенции приматов на его основные классы. AluS-субсемейство наиболее активно амплифи- цировалось в геноме приматов в период появления первых антропоидов, т, е. приблизительно 40 млн лет назад. AluY — самые «молодые» из них. Они по-прежнему активны в отдельных семействах человекообразных приматов (Quentin Y., 1988). В геноме человека все Alu-вставки приходятся на AluY-субсемейство или его производные. Из них наиболее представлены AluYaS- и AluYbS-субсемейства. 25 и 38 % локусов соответственно (Hedges D. J. et al., 2004).
D. J. Hedges et al. (2004) нашли двукратное увеличение вставок Alu у людей, по сравнению с их количеством у шимпанзе (Pan.
troglodytes). Но уровень различий Alu, интегрировавшихся с геномом шимпанзе, в 1,7 раз выше, чем у людей. Эти исследователи, также как и Р. Jem et 'al. (2006), обнаруживших разные сценарии активности эндогенных ретровирусов у людей и шимпанзе, пришли к выводу, что Alu-элементы у людей уже несколько миллионов лет проявляют значительно большую эволюционную активность, чем у шимпанзе (см. «Эволюционная роль HERV-Κ») (рис. 10).
Рис. 10. Субсемейства Alu, представленные в геноме человека (справа) и шимпанзе (слева). По D. J. Hedges et at. (2004)
Распределение Alu-повторов по геному человека неравномерно как между хромосомами, таки по их длине. Вхромосомах 14, 16, 21 А1и-по- следовательности концентрируются в области центромеры, а в хромосомах 4, 19, 20, X и Y человека выраженные кластеры Alu-повторов не найдены (рис. 11 и 12).
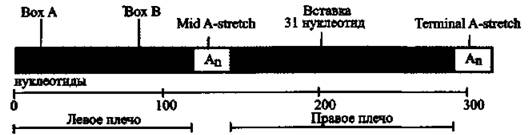
Рис. 11. Архитектура АІи-элемента
Размер Alu-повтора около 300 п. о. В 170 п. о. от начала элемента расположен сайт узнавания рестриктазой AluI (отсюда и его название). Повтор имеет характерную димерную структуру. Состоит из двух похожих, но не эквивалентных прямых повторов длиной около 130 п. о. — левое и правое плечи (left and right arms) Alu-элемента. Правое плечо содержит вставку из 31 нуклеотида, богатую аденином. Плечи разделены A-богатым регионом (Mid A-stretch), правое плечо закачивается коротким poly (А)-хвостом (terminal A-stretch). Левое плечо содержит функционирующие, но слабые А и В боксы (boxes) внутреннего промотора PHК-полимеразы III. По J. Hasler и К. Stmb (2006)
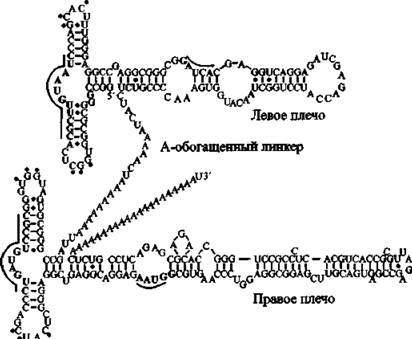
Рис. 12. Вторинная структура Alu-PHK Подчеркнутые буквы и точки показывают связывающие сайты SRP9/14 и спаривание оснований третичной структуры между двумя петлями, соответственно. По О. Weichenrieder et at.
(2000)Alu-элементы амплифицируются через РНК-производное посредством механизма ретротранспозиции, зависящего от других транспозонов (например, LINE-1), Сами они не кодируют генов белков. К настоящему времени установлено их участие в следующих процессах, способствующих эволюции приматов по типу анагенеза.
Экзонизация. Из тысяч Alu-ретроэлементов, найденных в интронах генов человека, значительная часть их последовательностей полностью или частично представлены в кодирующих регионах мРНК. Но и некодирующие участки мРНК не остаются без «внимания» ретроэлементов — более подробно см. «A-I-редактирование». Присутствие отдельных потенциально подверженных сплайсингу сайтов в Alu-консенснусных последовательностях дает основание предположить, что они были вовлечены в кодирующий регион через экзонизацию — процесс образования экзонов в интронных областях. Он стал возможен благодаря существованию у Alu-последователь- ностей участков (motifs), имеющих сходство с сайтами сплайсинга, или они образуют такой сайт посредством вариаций отдельных нуклеотидов интегрировавшимся Alu-элементом (рис. 13).
R. Sorek et аі. (2002) идентифицировали подмножество альтернативных сплайсиншрованных внутренних экзонов, из которых до 5 % были производными от Alu-элементов, и установили, что все экзоны, содержащие Alu, образовались в результате альтернативного сплайсинга. Эти же авторы показали, что до 85 % экзонов, содержащих Alu, являются производными от антисмысловых Alu-элементов и что благодаря существующим взаимодействиям между сайтами сплайсинга, некоторые мутации способны привести к повороту сплайсинга от альтернативного к конститутивному.
Эволюционный процесс, приводящий к экзонизации частичные и полные Alu-элементы, представляет собой случайное совпадение мутаций. Например, S. Singer et аі. (2004) реконструировали последовательность событий, приведших к образованию альтернативного 5'-экзона гена рецептора фактора некроза человека (p75TNFR). По крайней мере, пять мутационных событий, произошедших в течение 63 млн лет эволюции приматов, оказались необходимыми для случайной экзонизации и фиксации гена p75TNFR: 1) интеграция с геном примата Alu-элемента; 2) приобретение альтернативного сайта начала транскрипции; 3) образование альтернативного стартового кодона; 4) формирование сайта сплайсинга; и только после этого 5) случайная, в семь нуклеотидов, делеция привела к образованию открытой рамки считывания.
На рис, 14 суммированы результаты М. KruIl et аі. (2005), полученные ими при оценке возраста четырех генов и ранее описанного Singer et аі. (2004) гена p75TNFR.
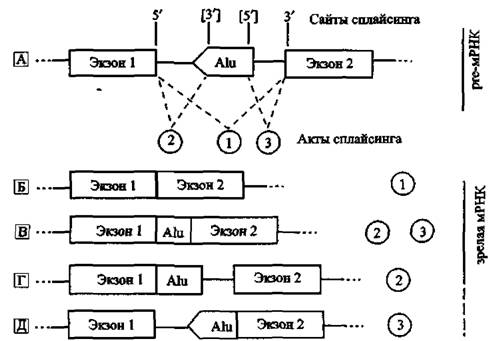
Рис. 13. Экзонизация интронов АШ-элементами Гипотетически Alu-элементы вставляются в ориентации, которая противоположна смысловой транскрипции интронного региона гена. Этот элемент имеет мажорный З'-сайт сплайсинга вблизи позиции 275 и мажорный 5'-сайт сплайсинга вблизи позиции 158. Использование альтернативного сайта сплайсинга Alu-элементом ведет к вариациям зрелой (матричной) РНК. А. Рге-мРНК подвергается актам сплайсинга (1; 2; 3). 3'- и 5'-сплайсинг-сайты показаны без скобок. Альтернативные сплайсинг-сайты включены в квадратные скобки. Б, Под актом 1 показан правильный сплайсинг без Alii-экзонизации. В. Экзонизация интронного Alu-элемента через использование его 5'- и 3'-сплайсинг-сайтов вместе с 5'-сплайсинг-сайтом экзона 1 и З'-сплайсинг-сайтом экзона 2, результат актов сплайсинга 2 и 3. Экзонизирован только AJu-элемент. Г. Экзонизация интронного Alu-элемента посредством его З'-сплайсинг-сайта вместе с 5'-сплай- синг-сайтом экзона 1; результат акта сплайсинга 2. Alu-элемент и З’-конец интрона 2 были экзонизированы. Д. Экзонизация интронного Alu-элемента посредством использования 5'-сплайсинг-сайта вместе с З'-сплайсинг-сайтом экзона 2; результат акта сплайсинга 3, Alu-элемент и 5'-конец интрона 1 экзонизированы.
По J. Hasler, К. Strub (2006)
Показанный на рис. 14 феномен аннулирования экзонизации уже сам по себе свидетельствует не ТОЛЬКО о возможности «прогрессивной эволюции* под воздействием процессов, в которых участвуют ретроэлементы, но и, наоборот, о возможности регресса вида и его замещения более примитивными эволюционными ветвями. Судя по этим данным, большинство из экзонизированных элементов интегрировались с геномом приматов перед их дивергенцией на антропоидов, В локусе p75TNFR отдельные изменения
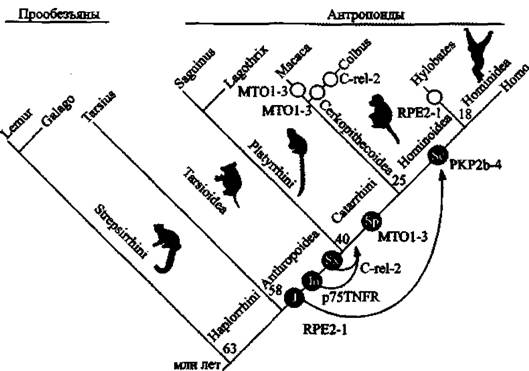
Рис.
14. Оценка возраста интеграционных актов пяти исследованных Alu-элементов и их аффинации в субклассыИнтеграционные акты показаны в виде черных кружков с белыми надписями, указывающими субсемейства Alu. Стрелками показано спроецированное время экзонизации в миллионах лет после интеграции Alu. Вероятное полное аннулирование экзонизации этих же генов для Cercopithecoidea и Hylobates показано открытыми кольцами. Ген RPE2-1 у людей найден в хромосоме 2q32-q33.3. Ген С-геІ-2 (изоформа C-rel прото-онкогенного протеина) у людей расположен в хромосоме 2р13-р12. Ген MTOI у людей расположен в хромосоме 6ql3.
Ген РКР2 (plakophilin) у людей расположен в хромосоме І2р11
последовали за интеграцией через период в несколько миллионов лет, рекрутировавшие эти последовательности как экзоны и зафиксировавшие их в линии, ведущей к Catarrhines. Теоретически Alu-элементы, ретро- позировав в MTO1-3 и в РКР2-4, могли проявитъ активность немедленно после интеграции. Ретропозированный в МТ01-3 Alu-элемент изначально обладал необходимой последовательностью для альтернативного сплайсинга. В случаях RPE2-I и РКР2Ь-4, вставки Alu-элементов были активированы критическими (cryptic — загадочный) 5'- или З'-сплайсинг-сайтами в интронных последовательностях, которые в случае с РКР2Ь-4 привели к дополнительной экзонизации рандоминизированной (randomized) интронной последовательности. Более подробно эти наблюдения описаны в работе М. Krull et аі. (2005).
Вставки Alu-экзонов вводят преждевременные терминальные кодоны или рамки считывания, а сами Alu-элементы генома человека действуют как очень большой резервуар альтернативных экзонов. В большинстве случаев экзонизированные последовательности являются либо нейтральными мутациями, либо проявляются вредным действием для отдельной особи, но это их влияние на особь незначительно, так как новый альтернативный продукт сплайсинга составляет только небольшую часть продукта обычного сплайсинга зрелой мРНК.
А-І-редактирование. Редактирование РНК — процесс, посредством которого нуклеотидные последовательности молекул РНК изменяются во время транскрипции и после нее. Модификация РНК включает нуклеотидные вставки и делеции, и модификации оснований.
Наиболее эффективным приемом модификации оснований является реакция гидролитического деаминирования, посредством которой цитозин конвертируется в урацил; а аденозин (А) в инозин (I), другое название процесса — А-І-редактирование. Эта реакция в условиях in vivo катализируется ферментами семейства аденозиндезаминаз (adenosine deaminase, ADAR), предпочтительно редактирующих аденозин, расположенный в двуцепочечном регионе молекулы РНК (Valente L., Nishikura К., 2005) (рис, 15).Точная роль А-І-редактирования в метаболизме клеток неизвестна, но она жизненно необходима для реализации клеточного цикла. Было показано, что нокаут (knockout — выбивать, испортить) гена ADARl эмбрионов мышей приводит к летальному эффекту через разрушение печени (Hartner J. С. et аі., 2004). Замена А на I является широко распространенным механизмом редактирования РНК. Более 90 % всех А-І-замен происходят в пределах Alu-элементов, содержащихся в мРНК. Установлено что А-І-редактирование предпочтительно встречается в некодирующих регионах мРНК. А так как ADAR-ферменты не являются специфичными для двунитевой РНК определенных сайтов, то граничащие с Alu основания также часто подвергаются редактированию.
РНК-редактирование может серьезно воздействовать на экспрессию генов на отдельных этапах жизненного цикла клетки. Так как инозин не может спариваться с урацилом, а только с цитозином, то редактирование может воздействовать на стабильность молекулы РНК посредством создания и разрушения вторичных структур.
Когда инозин будет распознан как гуанозин, то посредством аппарата трансляции и сплайсинга А-І-редактирование способно привести к аминокислотным заменам в кодирующей последовательности или к модификации сайта сплайсинга в нитроне, приводя к преждевременной терминации транскрипции или к искажению рамки считывания гена белка.
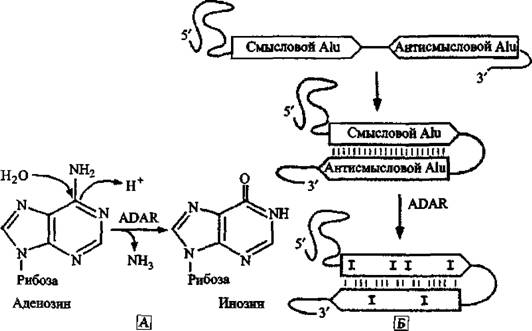
Рис, 15. Л-І-редактирование в Atu-злементах А. Реакция деаминирования аденозина через ADAR, ведущая к образованию инозина. Б. Внутримолекулярное спаривание оснований двух мРНК, содержащих Alu-элементы в противоположной ориентации. Спаривание оснований двух Alu-элементов приводит к формированию длинного стабильного двунитевого РНК-региона, в котором ADAR выполняет замену А на I. Два близко вставившихся Alu-элемента становятся субстратом для ADAR. По J. Hasler и К. Sirub (2006)
Участие в трансляции белков. C начала 1990-х гг. известно, что AIu-PHK, транскрибированная с ДНК Alu-элементов, постоянно представлена в цитозоле клеток приматов. Хотя Alu-элементы и содержат внутренние А- и В-боксы промотора РНК полимеразы III, но этот внутренний промоторный элемент слишком слаб для осуществления эффективной транскрипции Alu-элементов. В обычных условиях Alu-PHK представлена в очень низком количестве копий в цитозоле (IOi-IO4 молекулы на клетку). В стрессовых же условиях, таких как вирусная инфекция, экспозиция циклогексе- мидом или тепловой шок, уровень их экспрессии значительно увеличивается. AIu-элементы имеют высокий потенциал модуляции генной транскрипции посредством связывания отдельных транскрипционных факторов. AIu-PHK выполняет какую-то специфическую функцию в клеточном метаболизме и необходима для выживания клетки в условиях стресса, а транскрибируемая с ДНК Alu-элементов «утиль-РНК» («junk RNA») участвует в процессах клеточного метаболизма.
Первичные Alu-элементы были не более чем «эгоистичной ДНК» (см. «Проретроэлементы и проретровирусы»), но давление естественного отбора адаптировало их в геноме через закрепление за ними важных функций в регуляции генной экспрессии. Этот выигрыш регуляционной функции, известный как экзаптация (exaptation), участвовал в эволюции приматов по типу клало генеза и помог их дивергенции среди других млекопитающих (Brosius J., Gould S. J., 1992).
ARE. Значительная часть мРНК млекопитающих в З'-нетранслируемых регионах (untranslated regions, UTR) представлена последовательностями, содержащими аденин-урацилобогащенные элементы (adenine and uracil rich elements, AREs). Такие мРНК-последовательности обычно стабильны. Однако в последние годы обнаружены их разновидности, производные от Alu1 уменьшающие стабильность мРНК и приводящие се к распаду. А1и-по- вторы составляют до 5 % 3' UTR-последовательностей мРНК. На рис. 16 показан возможный механизм образования ARE.
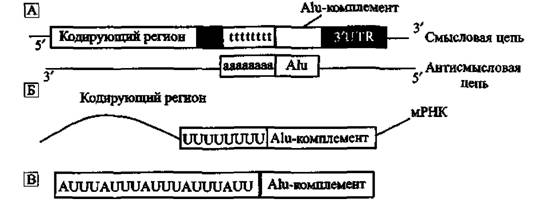
Рис- 16. Схематическое изображение образования ARE
A. Alu содержат poly-adenine (ро1у-А)-регион в концевой части. На схеме он показан как «аааааааа» (также см. рис. 10). Poly-A Alu в антисмысловой ориентации становится poly-Т (комплементарной poly-A) в смысловой цепи ДНК. Эта последовательность показана как «tUttttt». Б. Теперь, после транскрипции poly-T-pe- гиона, мРНК содержит poiy-uracile (poly-U(-регион. В. AU-обогашенные элементы находят именно в этих poly-U-регионах. По Jun An Hyeong et аі. (2004)
Hyeong Jun An et аі. (2004) обнаружили, что не менее половины наиболее протяженных ARE являются производными от (роІу-Т)-регионов, комплементарных (роІу-А)-регионам Alu, Они предположили, что Alu не только участвуют в экспрессии 5'-генов и в альтернативном сплайсинге интрон- ньіх регионов генома, но и, перемещаясь по геному через ретропозицию (см. «Эволюционная роль Ll-ретроэлементов»), они способны увеличивать или сокращать период полураспада мРНК (особенно в области 3' UTR) через образование ARE. Это еще один и, видимо, далеко не последний, из механизмов воздействия на экспрессию отдельных генов, обусловленный активностью ретроэлементов.
Эволюционная роль ретропсевдогенов. В геноме человека имеется более 20 тыс. псевдогенов — «молчащих» копий известных генов. Ретропсевдо- генез — это процесс образования псевдогенов через обратную транскрипцию с мРНК. Вследствие утраты регуляторных элементов ретропсевдогены почти всегда функционально неактивны, начиная с того самого времени, когда они впервые внедрились в хромосомную ДНК. По этой причине они обычно не подвергаются природной селекции и, следовательно, являются идеальным объектом для изучения нейтральной эволюции (Csuros M.. MikJos I., 2005). Например, по нуклеотидным различиям в псевдогенах дыхательного фермента цитохрома С. Grossman et аі. (2001) показали ускоренную эволюцию этого белка в период формирования предка человекоподобных приматов 40 млн лет назад.
Ретропсевдогены могут участвовать в образовании ретроэлементов, что продемонстрировали К. Szafranski et аі. (2004), использовавшие в качестве экспериментальной модели одноклеточный эукариотический организм — Dictyostelium discoideum. Эта саркодовая амеба содержит семейство LINE- подобных ретротранспозонов, специфически интегрирующихся с генами тРНК (TRE-элементы). Исследователями было установлено, что ретро- транспозированный рибосомальный 5Э(г58)-РНК-псевдоген в геноме амебы D. discoideum содержит в своем З'-концевом участке последовательность в 8 п. о,, производную от З'-конца TRE и полиаденилового хвоста. Ретропсевдоген фланкирован через дупликации, имеющие сайт узнавания (tar- get-site duplications), которые характерны для TRE и вставлены «выше» гена тРНК, точно так же, как типичный TRE. г5 S -ретропсевдоген имеет структурные особенности SINE, но он не может быть амплифицирован, вероятно, вследствие 5'-усечения, которое происходит во время инициации транспозиции. Обнаружение такого ретропсевдогена означает то, что SINE могут создаваться de поѵо путем обратной транскрипции LINE-транскриптов, если кодируемая LINE обратная транскриптаза диссоциирует от LINE РНК и «перескакивает» к другим клеточным РНК, транскрибируемым РНК-по- лимеразой III (см. «Классификация транспозируемых ретроэлементов» и рис, 3). К. Szafranski et аі. (2004) предположили, что высокая концентрация транскриптов генов полимеразы III в ядре клетки может способствовать переключению обратной транскриптазы на создание новых SINE, Эти короткие вставочные элементы и другие копии фрагментов геномной ДНК будут занимать новые участки в хромосоме, приводя к модификации уже существующие гены.
Однако роль самих псевдогенов в эволюции еще только выясняется. Обращает на себя внимание и то обстоятельство, что их количество в геноме многих современных позвоночных на порядки превышает количество активных геновліредшественн и ков. Например, число псевдогенов В-тубу- лина человека превышает количество функционирующих генов этого белка в 10 раз; а число псевдогенов гл ице рал ьде гид-3-ф ос ф атдегидроген азы мышей превышает количество активных генов этого фермента в 200 раз. К тому же дальнейшая судьба гена после его псевдогенизации не предполагает его участия в эволюции таксона. По данным D. Graur D. et аі. (1989), после того как ген теряет свою функцию (т. е. становится псевдогеном), он подвергается воздействию двух процессов, обычно относимых учеными к проявлениям нейтральной эволюции.
Первый включает в себя быстрое накопление точечных мутаций и неизбежно стирает сходство последовательностей псевдогена и его функционального гомолога, который эволюционирует гораздо медленнее. Нуклеотидный состав псевдогена будет становиться все более и более похожим на состав его нефункционального окружения, он будет «смешиваться» с этим окружением. Этот процесс получил название композиционной ассимиляции.
Второй процесс характеризуется тем, что псевдоген становится все короче по сравнению с функциональным геном. Сокращение его длины вызывается преобладанием делеций над инсерциями. Оценки показывают, что процессируемые псевдогены млекопитающих теряют около половины своей ДНК примерно за 400 млн лет. Но первые млекопитающие появились значительно позже, т. е. этот процесс у отдельных псевдогенов начался еще при появлении первых позвоночных. Геном человека, например, все еще содержит основные участки ДНК псевдогенов, найденные у самых отдаленных предков. Следовательно, процесс образования псевдогенов — это торможение эволюции таксона. По своему биологическому значению он противоположен ретровирусной эволюции и уравновешивает ее (более подробно см. в разд. 2.3).
Еще по теме Эволюционная роль Alu-элементов.:
- Патологические процессы, вызываемые перемещением по геному человека Alu-элементов*
- Эволюционные истоки половых различий в предпочтении партнера
- Активирование глиальных элементов коры.
- 2.3. Составные элементы маркетинга
- Супотницкий М. В.. Эволюционная патология. К вопросу о месте ВИЧ-инфекции и ВИЧ/СПИД-пандемии среди других инфекционных, эпидемических и пандемических процессов: монография / М. В, Супотницкий. — M.: Вузовская книга,2009. — 400 с., 2009
- Прекращение инвазии транснознруемых элементов.
- Конструкты и элементы медицинского технологического процесса
- 8.4.1 Опухоли из гладкомышечных элементов
- Элементы анатомии глаза
- 2. Аспекты изучения человека как элемента различных систем.
- Морфологические элементы.
- Элементы власти
- 3.2 Элементы классификации
- На коже имеют место морфологические элементы: первичные и вторичные.
- 1.2. Властные отношения как элемент власти
- Первичные полостные элементы