Ретроэлементы генома современного віща Homo sapiens
Классификация транспозируемых ретроэлементов. He- L TR-ретроэлемен- ты. L TR-ретроэлементы. Эволюционная роль HERV- К. Эволюционная роль L 1-ретроэлементов. Эволюционная роль Alu-элементов.
Эволюционная роль ретропсевдогенов. Прекращение инвазии транспозируемых элементов.Геном — полная генетическая система клетки, определяющая характер онтогенетического развития организма и наследственную передачу в ряду поколений всех его структурных и функциональных признаков. Суммарные данные о содержании разных видов последовательностей в геноме человека приведены в табл. 1.
Таблица I
Нуклеотидные последовательности, входящие в состав генома человека*
| Тип последовательности | Содержание, Ж |
| Экзоны генов | 1 |
| Интроны генов | 25 |
| Транспозируемые элементы | 45 |
| Большие дупликации | 5 |
| Простые повторы (микросателлиты)** | 3 |
| Другие межгенные последовательности | 20 |
* Из кн.: Тарантул В. 3., 2003.
** Повторяющаяся ДНК (repetitive DNA) в геноме человека включает гены, которые кодируют компоненты рибосом, организованные как тандемные повторы, повторяющиеся по 150-200 раз. Подобным же образом организована высокоповторяющаяся ДНК (highly repetitive DNA), включающая очень короткие последовательности, которые могут повторяться миллионы раз, — это так называемая миниса- теллишая ДНК (minisatellite DNA). Она включает блоки длиной по 10-100 п. о., характерные для каждого индивидуума (в данной работе не рассматриваются).
Классификация транспозируемых элементов.
Почти половину генома человека составляют различные транспозируемые элементы (transposable elements, TEs). Они делятся на два основных класса: ДНК-транспозоны (DNA transposones) и ретроэлементы (retroelements). Классификация транспозируемых элементов, их процентное содержание и приблизительное количество показаны на рис. 1.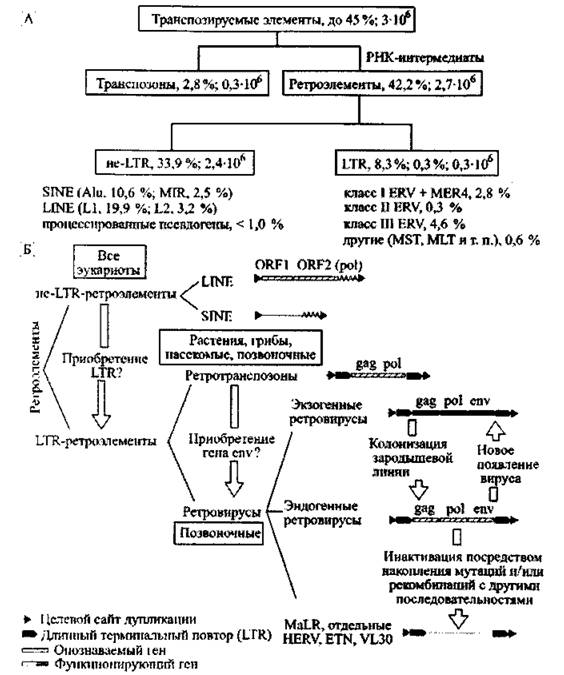
Рис. 1. Классификация транспонируемых элементов генома человека А. Классификация транспозируемых элементов человека по N. Banner! и R. Kurtli (2004). Б. Классификация ретроэлементов по N. de Par* seval и Т. Heidmann (2005). Короткие прямые повторы фланкируют все ретроэлементы, интегрировавшиеся с геномом хозяина через транспозицию. VL30, ETN — повторяющиеся LTR-элементы, найденные у мышей. MaLR— LTR-ретротранспозон, обнаруженный у млекопитающих
ДНК-транспозоны млекопитающих структурно сходны с бактериальными транспозонами. Подобно другим транспозируемым элементам (см. ниже), ДНК-транспозоны изменяют эволюционную траекторию своего хозяина благодаря следующим механизмам: 1) через изменение функции генов путем вставок генов, регуляторных элементов, «перетасовок» экзо- нов и интронов; 2) через индукцию хромосомных перестановок; 3) как источники кодирующей и не кодирующей ДНК, которая позволяет появляться различным генетическим новинкам, таким как новые гены и регуляторные последовательности (Feschotte C., Prithatn E,, 2007),
ДНК-транспозоны реплицируются без РНК-производного и теоретически способны перемещаться по геному по типу «разрезал и встроился» («cut and paste») через использование фермента транспозазы (transposase), Они фланкированы посредством инвертированных терминальных повторов (inverted terminal repeats, ITRs) и имеют одну открытую рамку считывания (open reading frame, ORF), кодирующую фермент транспозазу. ДНК- транспозоны фланкированы через короткие прямые повторы (short direct repeats, DRs), «приобретенные» в ходе интеграционных процессов; не образуют вирусных частиц и не могут покинуть клетку.
Эволюционная история ДНК-транспозонов приматов закончилась еще до «расщепления» приматов на виды Старого и Нового Света. J. К. Расе II и С. Feschotte (2007) исследовали не менее 40 семейств ДНК-транспозо- нов человека, включающих до 98 тыс. таких элементов, и установили высокую активность ДНК-транспозонов в эволюции млекопитающих и, в частности ранних приматов. Но перед радиацией приматов на предков видов-антропоидов (anthropoid primate ancestor) их транспозиционная активность прекратилась. Исследователи не обнаружили ДНК-транспозоны «моложе» 37 млн лет. Поэтому такие транспозируемые элементы считаются ими своего рода «окаменелостями» генома приматов (рис, 2).
Млекопитающие сосуществовали с рептилиями в триасе (230-190 млн лет назад), но лишь как дополнение к многообразию последних на планете. Основная масса ДНК-транспозонов (85 %; примерно 291 тыс. элементов) распространились среди млекопитающих в меловом периоде (135-66 млн лет назад), когда происходило вымирание рептилий. «Расцвет» млекопитающих пришелся на палеоген (66-25 млн лет назад). До 29 семейств (74 тыс. элементов) были активны у приматов перед их «растеплением» на антропоидов, и 11 семейств (23 тыс, элементов) интегрировались с геномом антропоидов. Следовательно, в ходе эволюции приматов наблюдаюсь устойчивое снижение активности ДНК-транспозонов, но почему это произошло, еще предстоит установить. Более подробно о роли ДНК-транспозонов в эволюции эукариотов см, в работе С. Feschotte, Е. Pritham (2007).
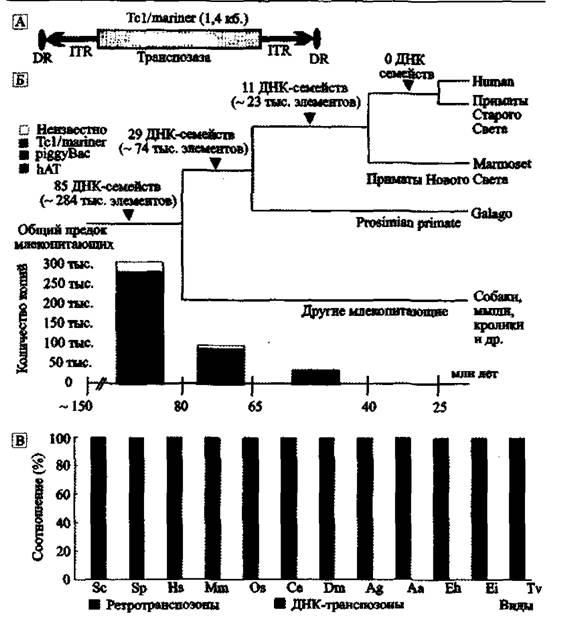
Рис. 2. ДНК-транспозоны в эволюции эукариотов А. Структура наиболее распространенного среди млекопитающих ДНК-транс- позона. По Е. М. Ostertag, Н. Kazazian (2001). Б. Активность ДНК-транспо- зонов в эволюции приматов. Столбики соответствуют количеству ДНК-эле- ментов, активных на отдельных этапах эволюции приматов (разбиты на суперсемейства). ДНК-транспозонов, активных после появления обезьян Нового Света, в геноме человека не обнаружено.
HAT, piggyBac и Ec I /mariner—семейства ДНК-транспозонов, обнаруженные в геноме человека. По J. К. Расе II, С. Feschotte (2007). В. Процентное соотношение ДНК-транспозонов и ретро- транс позонов относительно общего количества транспозируемых элементов эукариотических видов. По С. Feschotte, Е. Pritham (2007)Ретротранстзоны. В противоположность ДHК-транспозонаМ, они кодируют обратную транскриптазу (reverse transcriptase, RT) и перемещаются по геному через образование РНК-транскрипта. Образовавшийся транскрипт транскрибируется «обратно» в ДНК-транскрипт и встраивается в хромосому клетки. При перемещении ретротранспозонов соблюдается принцип — «копировался и вставился» («сору and paste»), как в «доброе старое время» протоклеточных образований. Ретроэлементы составляют 90 % всех транспонируемых элементов генома человека, большинство из них не активны, однако некоторые из них остались способными к ретротранспозиции. По этим данным получается, что структуры, подобные ВИЧ, обобщенно называемые ретроэлементами, и есть геном человека.
Ретротранспозируемые элементы (ретроэлементы) делят на две большие группы: способные к автономному существованию (эту их «автономность» нельзя понимать буквально, так как они зависят от ферментов репарации ДНК хозяина, необходимых для их транспозиции) и неавтономные ретроэлементы.
Выделены два класса автономных ретроэлементов: не-LTR (non-LTR) и LTR-элементы (LTR-elements), структурно сходные с ретровирусами, но утратившие функционирующий епѵ ген.
Длинные концевые повторы или LTR (long terminal repeats) — это прямые повторяющиеся последовательности на концах ДНК-копии генома ретровирусов, образовавшейся в результате обратной транскрипции. Каждый такой повтор состоит из трех элементов: U3-R-U5, длина которых составляет соответственно 170-1250, 10-80 и 80-100 т. п. о.; 3'-конец U5 сам содержит короткий инвертированный повтор, гомологичный последовательности на 5'-конце элемента U3, т. е. сама последовательность LTR фланкирована короткими инвертированными повторами; LTR участвуют в интеграции ДНК-копии генома ретровируса в геном клетки-хозяина, кроме того, область U3 каждого LTR несет промотор.
Промотор левого LTR участвует в транскрипции ДНК провируса, а промотор правого — последовательности ДНК клетки-хозяина вблизи сайта интеграции ретровируса. LTR фланкируют сложные элементы генома и участвуют в процессе их транспозиции (Khodosevich К. et аі., 2002).Неавтономные ретротранспозоны не кодируют белков. К ним относятся Alu-повторы (в геноме человека их более миллиона; см. «Эволюционная роль Alu-элементов»); псевдогены, образовавшиеся в результате обратной транскрипции (см. «Эволюционная роль псевдогенов»); и SVA-элементы (аббревиатура от заглавных букв S1NE-R, VNTR и Alu, описаны как «композитные ретропозоны; более подробно о SVA-элементах см. в разд. 4.3). Для своей транспозиции они нуждаются в активности автономных ретроэлементов. Например, SVA-элементы ретротранспозируются с помощью Ll-транспозонов (более подробно о механизмах транспозиции неавтономных ретротранспозонов см. в работе Ostertag Е. M., Kazazian H., 2001).
He-LTR-ретроэлементы. Очень древние ретроэлементы. Широко представлены среди простейших организмов. Два представителя этого семейства ретроэлементов встречаются в геноме человека в больших количествах. Это короткие вставочные элементы (short interspersed elements, SINE) с преобладанием Alu- и MIR-повторов и длинные терминальные вставочные повторы (long-terminal interspersed elements, LINE), представленные автономными LI и L2 последовательностями (см. «Эволюционная роль Ll-pe- троэлементов»). SINE обычно имеют длину IOCMOO п. о. и по большей части происходят от транскриптов генов тРНК, синтезированных посредством РНК-полимеразы III (pol III). SINE имеют внутренний промотор, который позволяет их транскрипцию РНК-полимеразой III. Они не имеют емкости, достаточной для поддержания генов амплификации, и их размножение в геноме человека зависит от LINE.
LTR-ретроэлементы. Составляют до 8 % генома человека, К ним относятся ретротранспозоны (retrotransposons), эндогенные ретровирусы (endogenous retroviruses, ERVs), человеческие эндогенные ретровирусы (human endogenous retroviruses, HERVs) и повторяющиеся элементы эндогенных ретровирусов человека (repeat elements with HERV origin), такие как SINE-R ретропозоны (SINE-R retroposons), Последние содержат участки последовательностей LTR HERV-K (оис.
3).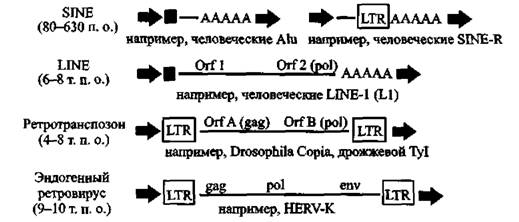
Рис. S. Структурные особенности наиболее частовстречающихся ретроэлементов
Стрелками показаны повторяющиеся последовательности, образованные во время интеграционных процессов. Заполненные квадраты соответствуют про- моторным регионам. A-последовательности показывают первоначальное полиаденилирование (original polyadenylation). По N, Bannert, R, Kurth (2004)
В геноме человека содержится не менее 98 тыс. ERV (Paces J. et al., 2002). Ген env ERV придает способность ретровирусу распространяться между клетками и индивидуумами. Отдельные LTR эндогенных ретровирусов и ретротранспозоны, образовавшиеся благодаря рекомбинационным процессам, в которых участвуют сами LTR-ретроэлементы, встречаются в геноме человека на один-два порядка чаще, чем эндогенные ретровирусы. Всего выделяется 6 суперсемейств LTR-ретроэлементов (табл. 2).
Таблица 2
Суперсемейства LTR-содержапшх ретроэлементов*

* По Р. Medstrand, L. N. van de Lagemaat, D. L, Mager (2002).
Эндогенные ретровирусы классов I и II проникли в зародышевую линию примитивных приматов как инфекционные ретровирусы и в последующем подвергались взрывной амплификации и транспозиции в периоды активной эволюции приматов. Другие суперсемейства, вероятно, соответствуют древним ретротранспозонам, которые амплифицировались на ранних этапах эволюции млекопитающих. Только небольшая часть «молодых» субтипов Alu и Ll non-LTR-элементов остаются активными в геноме человека (Medstrand Р. et аі., 2002).
В механизме «перехода» экзогенного ретровируса в эндогенный много неясного. Эндогенизации у людей известных экзогенных ретровирусов пока не зафиксировано. Вероятность таких событий для генома нашего вида нельзя исключать, так как сходные процессы обнаружены у других млекопитающих. Например, вирус рака молочной железы мышей, вирус мышиной лейкемии, вирус птичьей лейкемии и вирус кошачьей лейкемии встречаются у своих хозяев как в эндо-, таки в экзовариантах (Medstrand P., Mager D. L., 1998). Как будет показано ниже (см. «Эволюционная роль H E RV-К»), эндогенизация ретровирусов у гоминид происходит не только очень редко (в масштабах времени скорее геологических, чем отражающих продолжительность существования вида), но и в периоды каких-то важных эволюционных событий для этого семейства в целом.
Попытки таксономии эндогенных ретровирусов человека приводят ученых к замешательству. Предпочтительным приемом таких исследований является использование аминокислотной специфичности тРНК, которая гибридизуется с праймер-связывающим сайтом (primer-binding site). Например, HERV-K использует лизинспецифическую тРНК как праймер для инициации реакции обратной транскрипции. Но сегодня известно, что весьма различные ретровирусы используют этот же праймер. Кроме того, неполная информация по коротким участкам вирусов, содержащим мутации и делеции, делает их классификацию почти невозможной. К этому надо добавить лабораторные ошибки, когда отдельные ретровирусы выделяются и именуются произвольно в разных лабораториях (Bannert N., Kurth R., 2004).
Эволюционная роль HERV-K. Наиболее вероятной схемой появления в геноме человека мультикопийных семейств HERV N. de Parseval и Т. Heidmann (2005) считают следующую: экзогенные ретровирусы случайно инфицируют клетки зародышевой линии (germline cells) во время развития плода. После первичной «колонизации» генома ставшие эндогенными ретровирусы передаются вертикально. Амплификация копий «предкового» провируса осуществляется путем внутриклеточной ретротранспозиции и повторной интеграции в клетки зародышевой линии. Каждая новая колонизация таких клеток дает новое семейство или линию ERV.
Очень немногие из 30-50 идентифицированных групп эндогенных ретровирусов человека содержат открытые рамки считывания для генов трех основных структурных белков Gag, Pol и Env (см. разд. 1.3). Все они принадлежат к молодому семейству HERV-K, которое поддерживается в геноме приматов Старого Света (Old World monkeys, OWMs), включая человекообразных обезьян и людей. Всего же, по данным роі-специфического микроанализа, среди OWM выявлено не менее 8 семейств гаммаретровиру- сов, 9 семейств бетаретровирусов и 5 субгрупп HERV-L-элементов (Greenwood A. D, et аі., 2005). Происхождение многих эндогенных ретровирусов человека уходит в глубину эволюционной истории только приматов примерно на 30-45 млн лет (см. работы Sverdlov Е. D., 2000; и Hughes J. F., Coffin J. M,, 2005). В действительности они должы быть намного древнее. По крайней мере, 31 семейство HERV являются потомками независимых актов инфекции экзогенными ретровирусами (Belshaw R. et аі., 2005).
На рис. 4. показаны гипотетические модели дивергенции видов приматов, произошедшей в периоды «после» эпизоотий ретровирусных инфекций, имевших место 38-5 млн лет назад. Правда J. М. Coffin (2004), составивший большую часть схемы А данного рисунка, представляет дивергенцию приматов бесхитростно, как некое прогрессивное явление,
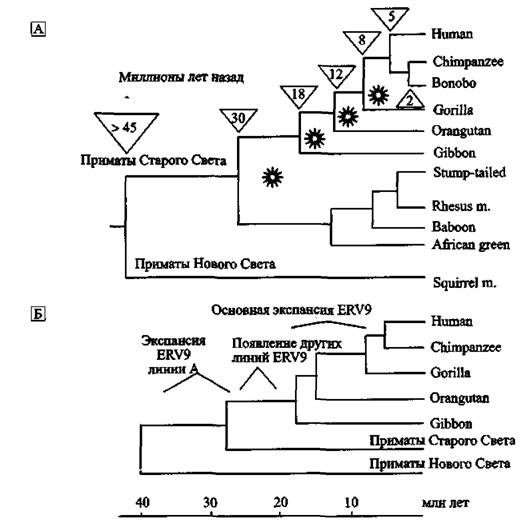
Рис. 4. Эндогенные ретровирусы в дивергенции видов приматов
А. Показано распределение эндогенных ретровирусов, интегрировавшихся с геномом передкового вида современных приматов в период от 30 до 5 млн лет назад. «Звездочками» указаны периоды ретровирусных эпизоотий, следы которых обнаруживаются в геноме современных приматов (см табл. 3). Числа в перевернутых треугольниках показывает ориентировочное время (в млн лет) расхождения отдельных эволюционных ветвей приматов (за основу мною взята схема Coffin J. M., 2004). Б. Эволюционная история эндогенного ретро- вируса семейства ERV9 на фоне эволюционного древа приматов — частный случай участия ретровирусов в дивергенции видов приматов. Экспансия ERV9 (линия А ретровируса) в геноме предкових видов современных приматов Старого Света началась 38-30 млн лет назад. Но наиболее активно экспансия ERV9 по геному приматов осуществлялась в период их дивергенции от гиббонов на высшие виды обезьян (16-6 млн лет назад). Максимум транспозиционной активности семейством ERV9 достигнут 8-6 млн лег назад, затем это ретровирусное семейство «угасло». По J. Costas, Н, Naverira (2000)
имеющее своей целью создание именно тех их видов, во главе которых сегодня как венец творения Природы стоит человек, на чем этот процесс, разумеется, заканчивается. Но так не бывает. У природы нет вечных любимчиков, маховик эволюции может ускоряться и замедляться, но вращаться он будет постоянно, пока существует жизнь.
Если придерживаться распространенной точки зрения на эндогени- зацию ретровирусов как на процесс «перехода» экзогенного ретровируса, вызывающего эпидемии среди своих новых хозяев (а для наших отдаленных предков правильнее использовать термин «эпизоотии»), в эндогенный вирус-мутант, неспособный образовывать вирусные частицы и передаваться горизонтально, то надо предполагать еще и ту цену, которую заплатил отряд приматов за эту «интеграцию». Такая «эндогенизация ретровируса» неизбежно должна сопровождаться массовым вымиранием отдельных видов и даже семейств приматов. Учитывая особенности этих инфекционных процессов, «эндогенизация» в нашем восприятии времени длилась бы десятки тысяч лет (см. Супотницкий М. В., 2000; 2006).
Оценки возраста ретровирусов делаются на основе сравнения последовательностей двух LTR и знания того, что HERV накапливают мутации со скоростью от 2,3х 10'9 до 5x10-9 замен нуклеотидов в год, т. е. одно изменение в нуклеотидной последовательности LTR каждые 200 тыс. — 450 тыс. лет. Некоторые ретроэлементы геномов приматов Старого Света имеют возраст не менее 55 млн лет (Bannert N., Kurth R., 2004).
Интересно отметить то, что обезьяны Нового Света (New World monkeys, NWMs) обычно либо вообще не имеют, либо имеют только сильно редуцированные копии ERV большинства классов (Greenwood A. D. et аі,, 2005). Следовательно, ретровирусное инфицирование вида возможно не при всех сценариях его существования. Однако если оно произошло и привело к эндогенизации ретровируса, то влечет за собой труднопрогнозируемые эволюционные последствия на протяжении нескольких миллионов лет. Объяснение причин отсутствия следов «ретровирусных атак» в геноме обезьян Нового Света представляет собой не менее интересную задачу, чем объяснение их наличия для обезьян Старого Света. Получается, что существуют либо неизвестные источники ретровирусов для приматов (и с ними не соприкасались приматы Американского континента), либо в природе имеются какие-то терминаторы ретровирусных эпизоотий, которые не представлены в Старом Свете.
А вот результат эволюции без «эндогенизации» ретровирусов, как говорится, «на лицо». Обезьян Нового Света относят к надсемейству примитивных широконосых обезьян. Это мелкие обезьяны с широкой хрящевой носовой перегородкой, с направленными вперед ноздрями и с когте-
образными ногтями. Большой палец не противопоставляется другим, полушария мозга гладкие. Свою цену за эволюцию они явно не заплатили.
Но теперь вернемся к тем, кто «платил за все». Ретровирусы семейства HERV-K у приматов Старого Света были активны перед и после эволюционного разделения человека и шимпанзе 5-6 млн лет назад. Некоторые из них встречаются только у людей, тем самым показывая, что они интегрировались с его геномом уже после разделения этих линий (табл. 3).
Таблица 3
Видовое распределение и время интеграции HERV-K-элементой в геном человека[3]
| HERV-K | Положение B хромосоме человека | Наибольшая дистанция до вида, в котором провирус был обнаружен | Оценка времени интеграции (млн лет) | Дата «расхождения» с общим предком (млн лет) |
| 4q32 | 166274445-166281673 | шимпанзе | 7,2-10,5 | 6 |
| HERVK(II) (Chr, 3) | 102893427-102902549 | горилла | 4,9-5,9 | 7 |
| I2q24 | 132277472-132283414 | горилла | 6,6-9,8 | 7 |
| I0pl4 | 6906147-6915609 | горилла | 9,0-12,6 | 7 |
| 19р13.1IA | 22549664-22556401 | горилла | 10,3-15,4 | 7 |
| 22qll | 22204481-22215171 | горилла | 28,6-38,9 | 7 |
| 9q34.3 | 136950603-136960065 | орангутан | 11,1-12,7 | 14 |
| Зр25 | 9864346-9871236 | орангутан | 13,4-19,8 | 14 |
| Iq23 | 163306258-163311916 | орангутан | 15,9-17,3 | 14 |
| t9p 13.11B | 20248400-20258515 | орангутан | 26,4-28,1 | 14 |
| I Iqll | 61892539-61907139 | гиббон | 17,5-21,0 | 18 |
| 19q 13.1 | 42289389-42298906 | гиббон | 21,0-36,3 | 18 |
| 6p22 | 28758347-28768714 | гиббон | 25,0-32,4 | 18 |
| 20q 11 | 32179289-32188037 | OWM | 12,8-18,3 | 25 |
| 6p21 | 42969390-42979344 | OWM | 7,4-13,1 | 25 |
гомологичной рекомбинации. По крайней мере, 1/3 из исследованных провирусов подверглась эктопической рекомбинации (ectopic recombination) (Hughes J. F., Coffin J. M., 2005). J. M. Coffin (2004) указывал на периоды в эволюции приматов «без активности ретровирусов». R. Belshaw et аі. (2004), исследовавшие епѵ ген HERV восьми семейств, считают, что реинфекция является наиболее общим механизмом поддержания и пролиферации эндогенных ретровирусов в их хозяевах (см. разд. 1.3, «Реинфекция»). Однако высокая частота реинфицирующих событий затрудняет точную оценку времени эвдогенизации ретровирусов на основе оценок дивергенции между LTR (рис. 5).
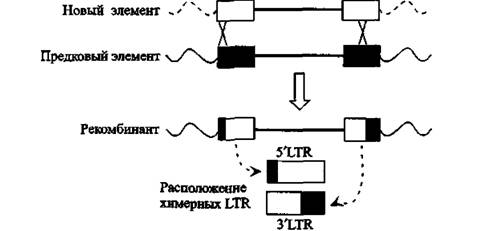
Рис. 5. Образование через гомологичную рекомбинацию химерных LTR в HERV-K6p21
LTR заново интегрировавшихся HERV-K-элементов показаны белым цветом. По J. F. Hughes и J. М. CofFm (2005)
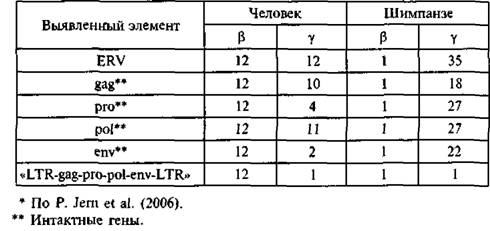
По данным Р. Jem et аі. (2006), в эволюции шимпанзе и людей участвовали разные эндогенные вирусы и с разными сценариями активности. Эти авторы нашли различия в недавней (т, е. имевшей место в ближайшие 5 млн лет) активности β- и γ-подобных эндогенных ретровирусов в геномах шимпанзе и людей. Две большие группы γ-подобных эндогенных ретровирусов (PtGl и PtG2) поддерживались в геноме шимпанзе и отсутствовали у людей; PtG-последователъности были наиболее сходны с двумя ERV бабуинов, но не с ретровирусами данного типа других шимпанзе или людей. Сама же γ-ретровирусная интеграционная активность у шимпанзе была разделена во времени от β-ретровирусной (табл. 4).
Для исследователей роли ретроэлементов в эволюции человека должно представлять интерес и обнаружение D. J. Hedges et аі. (2004) разных сценариев эволюционной активности Alu-элементов, также начавших свойотсчет после дивергенции видов Н, sapiens и Р. troglodytes (см. «Эволю-
Таблица 4
Свойства эндогенных ретровирусов, недавно интегрировавшихся с геномом людей н шимпанзе*
ционная роль Alu-элементов»), Еще более любопытные результаты дает сравнительный анализ экспрессии эндогенных ретровирусов в различных тканях разных видов приматов. Например, анализ 215 образцов РНК, полученных из мозга людей, показал явную специфичность экспрессионного профиля HERV разных семейств и классов (Frank О. et аі., 2005).
Недавние эксперименты позволили установить, что фундаментальные биологические различия между видами приматов являются следствием не столько вариаций в их генах, сколько результатом различий в экспрессии и регуляции одних и тех же генов (эволюция по типу анагенеза). Например, исследования, основанные на микроанализе ДНК, показывают, что экспрессия сложных генов человеческого мозга значительно превышает их же экспрессию у нечеловекообразных приматов. Но ткани, иные чем мозг, у этих же приматов не показывают значительных различий в экспрессии генов (Stengel A. et аі., 2006).
А. Stengel et аі. (2006) сообщили о собственных экспериментах по оценке экспрессии генов HERV в различных тканях приматов разных видов. Ими установлено, что большинство анализируемых HERV активно экспрессировались в тканях мозга человека, но оказывались либо полностью неактивными в аналогичных тканях обезьян Старого Света, либо их экспрессия была незначительной.
Данные, полученные О. Frank et аі. (2005) и А. Stengel et аі. (2006), интересно сопоставить с более ранними наблюдениями палеоантропологов по эволюции мозга человекообразных приматов, обобщенных в работе С. Оппенгеймера (2004). Его собственные объяснения эволюции человека сводятся к необходимости приспособления приматов к внешним факторам, среди которых он на первое место ставит похолодание климата, начавшееся 7-S млн лет назад. И в качестве адаптивного признака к холоду антрополог Оппенгеймер почему-то видит увеличение объема мозга человекообразных приматов, а не увеличение длины их шерсти или толщины костей черепа. Проанализируем собранные им данные применительно к вышеуказанным работам и к результатам исследования дивергенции видов приматов, полученных другими авторами (см. рис. 4).
По данным антропологических исследований, примерно 7-8 млн лет назад произошло резкое сокращение числа видов человекообразных приматов, совпавшее по времени с расширением площади безлесых травяных степей и глобальным похолоданием, продолжавшимся несколько миллионов лет. Но именно в этот период произошла дивергенция какого-то неизвестного вида приматов на виды, в последующем дивергировавшие на гоминоидов (наших ближайших предков), горилл, орангутанов, бабуинов и шимпанзе. «Списать» оба эти процесса только на «похолодание» не удается, так как тогда же вспыхнули массовые эпизоотии ретровирусных инфекций, оставивших в качестве «отпечатков» в геноме этих видов не менее семи типов эндогенных ретровирусов. Эпизоотии были настолько масштабными, что почти не сохранили в геноме выживших видов приматов «следов» других подобных эпизоотий за предшествующие несколько миллионов лет (см. табл. 3), Массовая гибель приматов снизила заполненность занимаемых ими экологических ниш и способствовала увеличению темпов видообразования у тех представителей их отряда, которые «прошли» через процесс эндогенизации новых ретровирусов. Тогда же стали появляться виды приматов (гоминоиды), которых сегодня палеоантропологи считают нашими предшественниками, т. е. приматов с увеличенным объемом и активностью мозга. Следовательно, ретровирусная эндогени- зация при условии освобождения экологических ниш, создала условия для эволюции отряда приматов по типу кладогенеза (рис. 6).
Любопытно и то, что с этого периода времени у предков шимпанзе и предков человека функционируют разные эндогенные вирусы и с разными сценариями активности (см. выше данные Jem Р. et аі., 2006). У крупных травоядных обезьян за весь период сравнения (5 млн лет) не было выявлено никаких признаков увеличения объема мозга, тогда как у гоминоидов обеих ветвей Homo (ergaster и babilis) и Parantbropus (boisei) такие изменения произошли. В этот период появилось не только несколько новых видов Ношо и Parantbropus со значительно большим объемом мозга, но и, что весьма показательно, объем мозга увеличился у всех гоминоидов в пределах каждого вида с 400 до 900 см3 (Elton S. et аі., 2001).
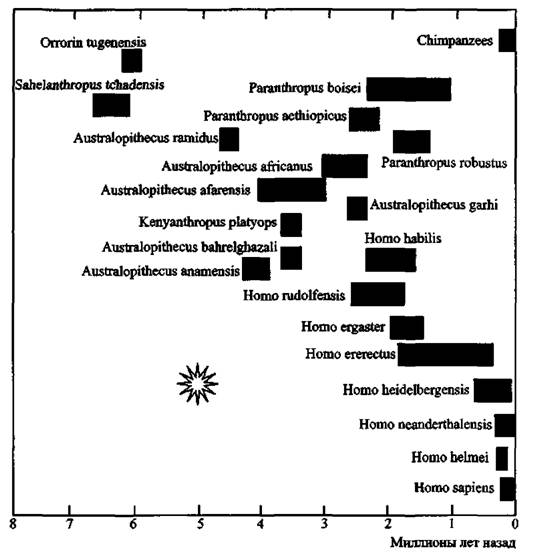
Рис. 6. *Взрывная» дивергенция видов гоминоидов (эволюция по типу кладогенеза)
Произошла 5-2 млн лет назад после процессов эндогенизации ретровирусов, наиболее представленных в нашем геноме в настоящее время. «Звездочкой» обозначена массовая ретровирусная эпизоотия, ее «следы» сегодня обнаруживаются в геноме приматов в качестве эндогенных ретровирусов. За основу взята схема С. Оппенгеймера (2004)
С. Оппенгеймер (2004) отмечает прерывистость увеличения объема мозга при переходе от древних гоминоидов к современному человеку. Он приводит следующий пример. Увеличение объема мозга между древнейшим Homo babilis, жившим примерно 2 млн лет назад, и Homo rbodesiense, жившим 1,07-1,3 млн лет назад, т. е. в период 700 тыс. лет, составило более
чем в 2,5 раза. В последующие же 1,2 млн лег, несмотря на тот факт, что гоминоидам было присуще некоторое увеличение объема мозга, от достижения объема мозга современного человека их отделяло всего б %. Фактически же за последние 150 тыс. лет у человека современного типа имело место снижение объема мозга, Оппенгеймер, привязавший свою теорию эволюции гоминоидов к похолоданиям климата, не смог скрыть своего удивления, когда не нашел очередных скачков роста объема их мозга в ледниковые периоды последнего миллиона лет и, в частности, ледникового периода, закончившегося 30 тыс. лет назад.
Эндогенные ретровирусы участвуют в эволюции приматов не только через увеличение активности и объема их мозга, но и посредством «частных улучшений» отдельных генов, когда они берут «на себя» новую функцию для генома (рис. 7).
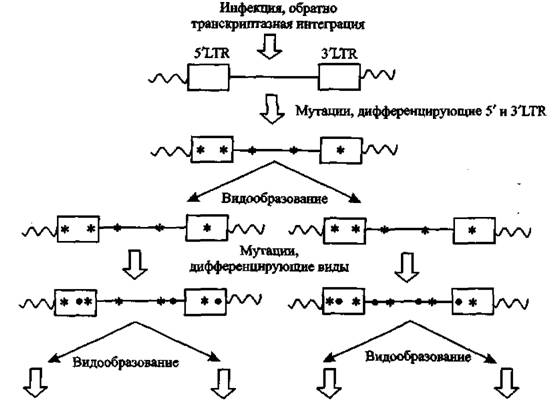
Рис. 7. Эндогенная ретровирусная эволюция LT R-Tiocледовательн остям интегрированного провируса соответствуют прямоугольники, между ними последовательность ДНК вируса (прямая линия). Клеточная ДНК показана волнистой линией. Мутации изображены как звездочки или точки. Их два типа: те, которые видоизменяют у потомков 5' и 3'LTR (звездочки), и после интеграции только предшествуют видообразованию; и те, которые непосредственно способствуют дифференциации видов, т. е. видообразованию (точки). По J. F. Hughes и J. М. CofTm (2005)
Количество задокументированных случаев, в которых эндогенные ретровирусы человека функционировали миллионы лет как промоторы или энхансеры (эволюция по типу анагенеза), в последние годы становится все большим (табл. 5).
Таблица 5
Примеры вовлечения ретровирусных последовательностей в регуляцию генов клеточных белков*

Сопоставление палеонтологических доказательств S. Elton et аі. (2001) и С. Оппенгеймера (2004) прерывистости в увеличении мозга гоминид с результатами исследований Р, Jem et аі. (2006) и J. М. CofTm (2004) по эндо- генизации ретровирусов в геноме приматов, позволяет сделать предположение, что оба этих явления находятся в причинно-следственной связи и «укладываются» в теорию прерывистого равновесия эволюции видов (пунктуализм), сформулированную в 1972 г. С. Гоуддом (Stephen Jay Gould1 1941-2002) и Н. Элдриджем (Niles Eldredge1 р, 1943). Их теория дополняет дарвиновскую теорию постепенной эволюции (градуализм).
Теория прерывистого равновесия эволюции видов предполагает чередование длительных периодов стабильности, когда основные черты вида сохраняются неизменными, и коротких периодов быстрых изменений, в ходе которых вид преобразуется — либо целиком превращается в другой вид, либо делится на два или более новых вида, либо «отпочковывает» их от себя (Pagel М. et аі., 2006). Судя по приведенным выше данным, реализуется этот эволюционный механизм после массовых ретровирусных эпизоотий, заканчивающихся эндогенизацией ретровирусов в геноме выживших видов и наращиванием генома вица-хозяина путем образования новых собственных копий ретроэлементов; его усложнения путем обра-
зования НОВЫХ экзо но в из интронов и/или увеличения количества генов, подвергающихся альтернативному сплайсингу.
Заканчивая рассмотрение роли эндогенных ретровирусов в эволюции человека, важно отметить тот факт, что процесс эндогенизации отдельных ретровирусов имел место и после сформирования вида Homo sapiens, т. е. приблизительно 170 тыс. лет назад — оценка С. Оппенгеймера (2004). Например, возраст обнаруженного G. Turneret аі. (2001) провируса HERV- Kll3 не превышает 100 тыс. лет. Он локализован в хромосоме 19 (19р13.11) и не полностью зафиксирован в человеческих популяциях. Генотипиро- вание генетически различных популяций показало зависимость его аллельной частоты от исследуемой этнической группы. Провирус весьма распространен среди людей, живущих в отдельных регионах Африки и Азии.
Основываясь на данных G. Turner et аі. (2001), N. Bannert и R. Kurth
(2004) предположили, что полностью интактные и активные аллели HERV все же либо представлены в популяциях людей с очень низкой частотой, либо встречаются с высокой частотой, но в генетически разделенных этнических группах. G. Turner et аі. (2001) полагают, что их находка показывает способность HERV-K реинфицировать популяции людей в недавнем эволюционном прошлом, и что HERV-K113 продолжает реинфицировать людей даже сегодня (см, разд. 1.3, «Реинфекция» и раэд. 4.3, «Эпидемиология LT R-p етроэле ме нтов»>).
Эволюционная роль Ll-ретроэлемеитов. Обнаруженные в геноме современного человека Ll-ретротранспозоны имеют свою эволюционную историю, насчитывающую не менее 100-150 млн лет (Furano А,V., 2000, Han К. et аі., 2005); т. е. в известном нам вдде они существовали у класса млекопитающих еще во времена господства рептилий. Сегодня Ll образуют 16 различных семейств (L1PA16-L1PA1). Их активность в геноме человека значительно большая, чем HERV. Эффективно дуплицируя сам себя, LI, вероятно, играют ключевую роль в увеличении генома вида посредством размножения нетранспозируемых Alu- и SVA-элементов и образования ретропсевдогенов (см. ниже «Эволюционная роль ретро псевдогенов»). Структура типичного полноразмерного Ll-элемента показана на рис. S.
Механизм репликации Ll аналогичен другим РНК, он включает образование транскриптов других ретроэлементов, например, Alu. У млекопитающих большинство Lt неактивны из-за вставок, делеций и точечных мутаций. Из 520 тыс. Ll-ретротранспозонов генома человека только около 5 тыс. имеют полноразмерный геном. Вставочная история Ll-элементов генома человека в основном написана семейством Та-1, составляющим до 90 % их популяции. Это семейство содержит значительно большее количество полноразмерных транспозонов, чем L1PA2- и L1 РА5-семейства,
І.2- Ретроэлементы генома современного вида Homo sapiens
35
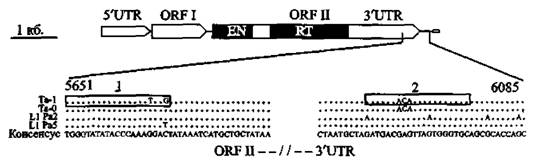
Рис. 8. Структура типичного полноразмерного Ll-элемента 5'-нетранслируемый регион (5'-UTR) — является внутренним промотором транспозона и не зависит от окружающих его нуклеотидных последовательностей, Далее идут: открытая рамка считывания I (ORF I), кодирующая 40 кДа (р40) РНК-связывающий белок (RNA-binding protein); короткий межгенный регион (intergenic region) протяженностью 63 п. о., содержащий стоп-кодон для всех рамок считывания (мРНК Ll атипична для мРНК млекопитающих, так как она бицистронна); и малоэффективная открытая рамка считывания 11 (ORF II), кодирующая Ll-репликаэу (LI replicase). Она содержит три консервативных домена: NH2-TepMJiHaaьный домен эндонуклеазы (EN), центральный домен обратной транскриптазы (RT) и СООН-терминальный цинк-шарнирный домен (СООH-termіnal zinc knuckle-like domain — на схеме не показан). Далее идет З'-нетранспируемый регион (З'-UTR), содержащий консервативный G-обогащенный полипуриновый мотив (G-rich polypurine motif). AATAAA поли A-сигнал (ААТААА роІІА signal) требуется для терминации РНК полимеразы II типа. Геномные копии Ll заканчиваются А-обогащенным участком (poliA tail, показан прямоугольником). На рисунке также показаны нуклеотидные вариации разных семейств LI-элементов (Та-1, Та-О, L1PA2 и L1PA5). По Е. М. Ostertag,
Н. Kazazian (2001); S. Boissinot et аі. (2004)
которые считают более древними. Для интеграции Ta-I предпочитают области с низким содержанием GC. S, Boissinot et аі. (2004) нашли, что Ta-1-эле- менты вставлены в интроны 46 известных генов и они в два раза чаще ориентированы в антисмысловом направлении в отношении этого гена (67 %), чем в смысловом (33 %).
Механизмы воздействия Ll на геном человека показаны на рис. 9.
А. LI-элемент распространяется по геному через транспозицию. Он копируется по принципу «сору and pasty». Белки LI синтезируются с их мРНК (ш-предпочтение). Следовательно, активны только полноразмерные элементы с двумя открытыми рамками считывания. Активный LI сначала транскрибируется m siti, затем его РНК обратно транскрибируется и интегрируется с геномом в новом участке. И-ретротранспозон может интегрироваться с хромосомой в виде полноразмерной копии (А). Однако- часто он оказывается 5'-усеченным (В) или 5'-инвертированным и усеченным (С), образуя неактивные копии исходного регротранспозона.
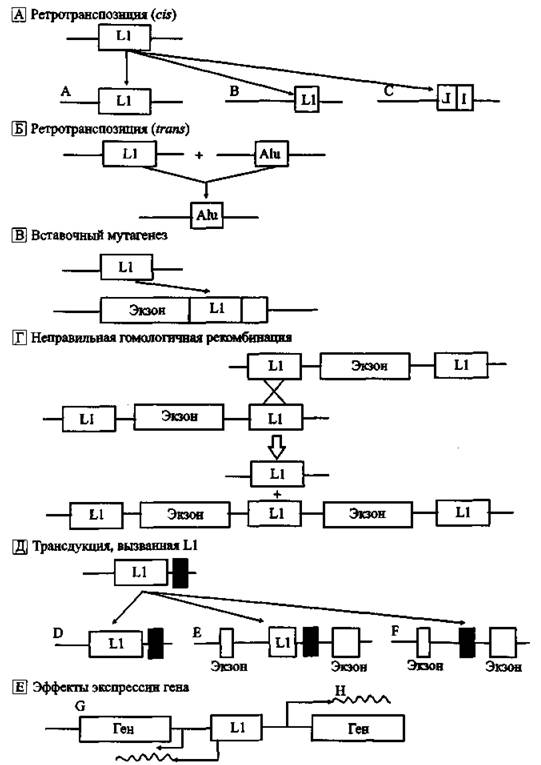
Рис. 9. Механизмы воздействия Ll-ретротранспозонов но геном человека По Е. М, Osiertag, Н, Kazazian (2001)
Б, отдельные неавтономные ретротранспозоны, такие как Alu, могут быть транспозированны in irons механизмом, используемым LI, в даль* нейшем способствуя увеличению размера генома.
В. Ll случайно вставляется в последовательность гена, вызывая генетическую болезнь (см. разя. 4.3).
Г. Ll может вызывать генетическое поражение через дупликацию или делению гена после неправильной гомологичной рекомбинации.
Д. РНК-полимераза Ll часто обходит принадлежащий ретроэлементу поли A-сигнал, использует такой же, лежащий по направлению транскрипции, поэтому последовательность ДНК хромосомы, фланирующая Ll З'-саЙт (заштрихованный прямоугольник), может быть перенесена в другой участок генома (D). Ретропозиция З'-экзона в другой ген может привести к «перетасовке* экзонов (Е, F). Поданным! L. Gooldieret аі. (2000), приблизительно 20 % Ll вставок содержат З'-трансдуцируемые последовательности. Обычно размер таких вставок находится в пределах 500-3 кб.
Е. Экспрессия гена может быть усилена через присутствие LI. Отдельные Ll имеют антисмысловые Pol ІІ-промоторы, которые влияют на экспрессию находящихся в непосредственной близости генов (G). Другие Ll могут выполнять функции энхансеров и регулировать гены, находящиеся на некотором расстоянии от них (Н).
Примером участия Ll-ретротранспозонов в эволюции человека по типу анагенеза является образование секретируемых форм человеческого трансмембранного белка атграктина (attractin). Ll-ретротранспозируемый элемент обеспечил преждевременный стоп-кодон и полиааенилашюнный сайт, ответственные за синтез усеченного растворимого аттрактина, Обе формы, трансмембранный и растворимый белки, вовлекаются в клеточные взаимодействия в течение воспалительного процесса. Таким образом, вставки Ll-ретроэлементов в данном конкретном случае создали для вида Homo sapiens более тонкие механизмы регуляции воспалительных ответов (Tang W. et аі., 2000).
Еще по теме Ретроэлементы генома современного віща Homo sapiens:
- Ретровирусное окружение вида Homo sapiens
- ГЕНОМ ЧЕЛОВЕКА
- Нелетальные повреждения генома клетки
- Нестабильность генома.
- Общее деметилирование генома.
- Патологические процессы, вызываемые перемещением по геном; человека LI*
- 7. Понятия генотипа, генома и генофонда. Понятие фенотипа.
- М.В. Угрюмова. НЕЙРОДЕГЕНЕРАТИВНЫЕ ЗАБОЛЕВАНИЯ: от генома до целостного организма. В 2-х томах. Том 1 / Под ред. М.В. Угрюмова. - М.: Научный мир,2014. - 580 с., 2014
- М.В. Угрюмова. НЕЙРОДЕГЕНЕРАТИВНЫЕ ЗАБОЛЕВАНИЯ: от генома до целостного организма. В 2-х томах. Том 2 / Под ред. М.В. Угрюмова. - М.: Научный мир,2014. - 848 с., 2014
- Билет 13. Функции современной психодиагностики.
- Современные рекомендации
- 9.2. Современная социальная психология
- Современные психологические подходы
- Современные подходы к оценке эффективности технологий в АГС.