2.1. Эмбриогенез и дифференцировка органов репродуктивно-половой системы
Прежде чем коснуться аспектов эмбриогенеза, следует напомнить некоторые постулаты и принципы функционирования генетического аппарата человека (см. цв. вкл., рис. 2.1 – 2.3).
Первый уровень построения генома предполагает организацию ДНК с гистоновыми белками – образование нуклеосом.
Две молекулы специальных нуклеосомных белков образуют октамер в виде «катушки», на которую наматывается нить ДНК. На одной нуклеосоме размещается около 200 пар оснований. Между нуклеосомами остается фрагмент ДНК размером до 60 пар оснований, называемый линкером. Этот уровень укладки позволяет уменьшить линейные размеры ДНК в 6 – 7 раз (см. цв. вкл., рис. 2.4).На следующем уровне нуклеосомы укладываются в фибриллу (соленоид). Каждый виток составляет 6 – 7 нуклеосом, при этом линейные размеры ДНК уменьшаются до 1 мм, то есть в 25 – 30 раз.
Третий уровень компактизации – петельная укладка фибрилл – образование петельных доменов, которые под углом отходят от основной оси хромосомы. Их можно увидеть в световой микроскоп как интерфазные хромосомы типа «ламповых щеток». Поперечная исчерченность, характерная для митотических хромосом, отражает в какой-то степени порядок расположения генов в молекуле ДНК.
Если у прокариот линейные размеры гена согласуются с размерами структурного белка, то у эукариот размеры ДНК намного превосходят суммарные размеры значимых генов. Это объясняется мозаичным (экзон-интронным) строением гена, когда экзоны – фрагменты, подлежащие транскрипции, – чередуются с интронами – незначащими участками (рис. 2.5, см. цв. вкл., рис. 2.6, 2.7).
Рис. 2.5. Гаплоидный набор хромосом у мужчины
Последовательность генов сначала полностью транскрибируется синтезирующейся молекулой РНК, из которой затем вырезаются интроны. Экзоны сшиваются, и в таком виде информация с молекулы иРНК считывается на рибосоме.
Причиной колоссальных размеров ДНК является большое количество повторяющихся генов. Некоторые гены повторяются десятки или сотни раз, а есть и такие, у которых встречается до 1 млн повторов на геном. Например, ген, кодирующий рРНК, повторяется около 2 тыс. раз.Развитие половых желез в эмбриогенезе (Балаболкин М. И., 2002) обусловлено набором половых хромосом, образующихся после оплодотворения яйцеклетки. Кариотип 46XX определяет развитие яичников, а 46XY – яичек.
Мужские и женские гонады развиваются из трех различных компонентов: целомического эпителия, мезенхимы и примордиальных герминальных клеток. Первичная половая дифференцировка – это процесс развития половых гонад, который начинается на 6 – 7-й неделе эмбрионального развития. Почки, надпочечники и половые железы развиваются в тесном взаимодействии, имеют общее происхождение и являются производными одной и той же области примитивной мезодермы. Под влиянием факторов транскрипции, в частности фактора WT1 (туморосупрессор опухоли Вилмса), клетки мезодермы трансформируются в почечный примордиум и адреногенитальный примордиум. Последний под влиянием двух факторов транскрипции – SF1 (ген стероидогенного фактора 1, СФ1) и DAX1 – в последующем развивается в кору надпочечника и первичную гонаду. Для развития и функционирования как коры надпочечников, так и первичной гонады требуется нормальная экспрессия SF1. Принято считать, что его действие заключается не в инициации, а в поддержании развития и образования первичной гонады и ранней фазы развития репродуктивного тракта. Следует отметить, что за развитие коры надпочечников отвечает фактор транскрипции DAX1.
Ген SF1 у человека локализуется на хромосоме 9q33 и содержит семь экзонов, включая два некодирующих. SF1 является ядерно-рецепторным фактором транскрипции и регулирует экспрессию многих генов, в том числе генов, кодирующих стероидогенез кортикостероидов. Исключительная важность SF1 в эмбриогенезе гонад подтверждается работой J. C. Acherman (1999), в которой доказано, что мутация гена SF1 сопровождается нарушением половой дифференцировки.
У больного с женским фенотипом сразу после рождения отмечалась надпочечниковая недостаточность. Однако при обследовании был установлен мужской кариотип 46XY. Кроме того, в области живота определялись структуры протоков Мюллера и маленькие тестисподобные образования.Кроме SF1, первичная роль в развитии примордиальной гонады бипотенциальных протоков принадлежит опухольсупрессорному фактору Вилмса (WT1 или ВО1). Локус гена WT1 у человека локализуется на хромосоме 11p13 и фактически состоит из двух генов – WT1 и WIT1. Экспрессия этого гена у эмбриона человека выявляется начиная с 28-го дня беременности в тканях производных мезодермы – почках, гонаде (клетки Сертоли), мезотелии, а также в спинном и головном мозге. Мутации гена WT1 идентифицированы у больных с Дэнис – Дрэш синдромом (DDS). Указанный синдром характеризуется триадой:
1) гонадальный дисгенез с нарушенной маскулинизацией у эмбриона мужского пола;
2) нефропатия;
3) предрасположенность к опухоли Вилмса.
Как показали исследования S. Barbaux (1997), при синдроме Фрайзера (FS) также имеется мутация гена WT1. Клиническая картина синдрома Фрайзера незначительно отличается от триады Дэнис – Дрэш синдрома и включает:
1) гонадальный дисгенез;
2) нарушенную маскулинизацию плода мужского пола;
3) прогрессивную гломерулопатию;
4) гонадобластому.
Описан также синдром, по клинической картине близкий синдрому Фрайзера, при котором имеет место гонадальный дисгенез.
Таким образом, на самых ранних стадиях развития примордиальной гонады и бипотенциальных протоков большое значение имеют гены SF1 и WT1, а для развития бипотенциальных протоков еще и ген Wnt4. Зародыш гонад (примордиальная гонада) бипотенциален и состоит из двух частей: кортикальной и медуллярной. При двух функционально нормальных Х-хромосомах кортикальная часть индифферентной гонады развивается в яичник. Гены, расположенные в перицентромерной области Y-хромосомы, определяют развитие медуллярной части в яичко. Однако процесс половой дифференцировки более специфичен и помимо указанных хромосом контролируется дополнительно несколькими генами.
Этот процесс является растянутым во времени и одновременно координируется экспрессией одних генов и регрессом других.Длительное время считалось, что гены, определяющие развитие бипотенциальной гонады в яичко, идентичны генам, которые кодируют H-Y-антиген, являющийся клеточно-поверхностным белком. Он способен оказывать непосредственное действие на дифференцировку первичной гонады в яичко. Однако тщательный анализ больных с различными нарушениями половой дифференцировки показал, что H-Y-антиген не идентичен Y-гену, контролирующему дифференцировку яичка. Еще W. K. Silvers установил, что H-Y-антиген локализуется на длинном плече Y-хромосомы и достаточно далеко от гена, определяющего развитие пола эмбриона. При детальном изучении карты хромосом у лиц с набором 46ХХ-половых хромосом при наличии яичек и мужского фенотипа установлено, что у них имеются участки последовательности Y-хромосомы на Х-хромосоме. На основании этого было высказано предположение, что на Y-хромосоме имеется участок последовательности, названный фактором, определяющим развитие яичка, который локализовался у этих больных в псевдоаутосомальной области. Ген, ответственный за развитие яичка, был идентифицирован D. C. Page [et al.] (1987), а клонированный ими участок хромосомы был назван ZFY. Указанная последовательность (ген ZFY) выявлялась у 46ХХ-фенотипичных мужчин, но отсутствовала у 46XY-фенотипичных женщин. Проведенные исследования позволяют считать, что ZFY является небольшой частью Y-хромосомы, отвечающей за дифференцировку плода. Однако у некоторых больных ZFY обнаруживали на аутосомах. Исследователи описали четырех мужчин с 46ХХ-набором половых хромосом, у которых наследуемый участок Y-хромосомы не включал ZFY (Palmer M. S. [et al.], 1990). Было установлено, что ZFY не является фактором, ответственным за развитие яичка. Почти одновременно другие ученые опубликовали уточняющие данные о локализации гена в области рY 53.3, ответственного за пол эмбриона, назвав его SRY (Sinclair A. H. [et al.], 1990).
По их мнению, SRY – истинный фактор, определяющий развитие яичка. Он экспрессируется только в яичках и отсутствует в легких и почках взрослого мужчины. Область ДНК, в которой локализуется SRY, ответственна также за кодирование двух ключевых ферментов, участвующих в дифференцировке первичной гонады по мужскому типу: 1) ароматазы Р450, контролирующей конверсию тестостерона в эстрадиол; 2) фактора или гормона, ингибирующего развитие протоков Мюллера, который вызывает регресс указанных протоков и способствует дифференцировке тестикул.Тем не менее SRY участвует в процессах половой дифференцировки в тесном взаимодействии с геном Z (McElreavey K. [et al.], 1993). Его функция в норме заключается в угнетении специфических мужских генов. В случае нормального мужского генотипа 46XY ген SRY продуцирует белок, угнетающий ген Z, и специфические мужские гены активируются. При нормальном женском генотипе 46ХХ и отсутствии SRY ген Z активируется и угнетает специфический мужской ген, что создает условия для развития по женскому типу. У 46ХY-мужчин с мутацией гена SRY, что сопровождается активацией гена Z с угнетением специфического мужского гена, происходит развитие по женскому типу. У 46ХХ-женщин при отсутствии у них SRY должна быть мутация гена Z, сопровождающаяся невозможностью его экспрессии, что способствует осуществлению дифференцировки по мужскому типу. Эта гипотеза объясняет генез развития женского фенотипа при 46XY. У этих лиц имеется интактный ген SRY и, вероятно, мутация гена Z.
Ген SRY локализуется на коротком плече 6-й хромосомы (центромерно к псевдоаутосомальной области Yp 11.3), а продукт этого гена комплексуется с различными участками ДНК. Ген SRY выполняет основную роль в половой дифференцировке и способен вызвать развитие обратного пола. Он экспрессируется в половой полоске только в определенное время эмбрионального развития, когда происходит образование тестикулярных зачатков. Мутация и делеция этого гена имеется у XY-мужчин.
Кроме того, ген SRY на уровне ДНК непосредственно регулирует активность других факторов транскрипции промоторной области гена ароматазы Р450, конвертирующей тестостерон в эстрадиол с угнетением этого процесса у эмбрионов мужского пола, а также ген антимюллерова гормона, ответственного за регрессию протоков Мюллера.
Несмотря на то что ген SRY принимает активное облигатное участие в превращении бипотенциальной гонады в яички, имеются данные, свидетельствующие о том, что одного указанного гена недостаточно для развития яичек (Ramos E. S. [et al.], 1996; Teebi A. S. [et al.], 1998). Имеются дополнительные области на Х-хромосоме и на аутосомах, участвующие в процессах трансформации первичной гонады в яичко. Помимо гена SRY, в указанных процессах половой дифференцировки мужской гонады важная роль принадлежит гену SOX9. К группе SOX-генов (SOX – гены, комплексирующиеся с областью SRY HMG) относят несколько генов (SOX1, SOX2, SOX3 и др.), участвующих в процессах половой дифференцировки первичной гонады. У человека ген SOX9 локализуется на хромосоме 17q 24 – 25 на участке, названном участком аутосомного изменения пола (SRA1), и кодирует фактор транскрипции, играющий важную роль в развитии яичка, в частности, отвечает за дифференцировку клеток Сертоли (Morais da Silva [et al.], 1996). Экспрессия гена SOX9 определяется в яичках в области семявыносящих канальцев на 18-й неделе эмбрионального развития вслед за экспрессией гена SRY. Исследования последних лет позволяют считать, что ген SRY активирует экспрессию гена SOX9, который прямо или опосредованно участвует в развитии клеток Сертоли, тогда как ген DAX1 репрессирует SOX9, угнетая развитие клеток Сертоли и являясь непременным условием трансформации первичной гонады в яичники.
Сравнительно недавно доказано участие гена Dhh и белка этого гена в процессах взаимодействия клеток Сертоли с герминальными клетками (Bitgood M. J. [et al.], 1996). Экспрессию гена Dhh выявляют в предшественниках клеток Сертоли вскоре после активирования гена SRY, и она продолжается во взрослом состоянии. Белок гена Dhh регулирует стадию митоза и мейоза в мужских герминальных клетках. Так, в период эмбриогенеза он контролирует пролиферацию герминальных клеток, а во взрослом состоянии – их созревание.
Об участии Х-хромосомы в дифференцировке первичной гонады свидетельствуют клинические данные не только об ингибировании развития яичек в выявлении Х-связанных рецессивных форм XY-гонадального дисгенеза, но и при удвоении так называемой DSS-области, локализованной на Х-хромосоме. Этот DSS-локус (дозочувствительный локус, ответственный за пол) локализуется в области 160 kb хромосомы (160 kilobase, или 160 тысяч пар нуклеотидов) Хp21 (Bardoni B. [et al.], 1994). При удвоении этого локуса наблюдают регресс яичек даже при наличии интактного гена SRY. Делеция этой области не влияет на образование яичка. Данные исследователей позволяют считать, что DSS-область не воздействует на состояние гена, ответственного за пол. Следует отметить, что A. Ion [et al.] (1996) идентифицировали у больного талассемией еще один сцепленный с Х-хромосомой ген – ХН2. Он участвовал в развитии яичек, мутация которых была выявлена у наблюдаемого ими пациента.
Установлено, что на хромосомах 2q (Slavotinek A. [et al.], 1999); 9p (Veitia R. [et al.], 1997), 10q (Wilkie A. O. [et al.], 1993) и 17q локализуются аутосомные гены, вовлеченные в развитие тестикул. Аутосомальные факторы ответственны за агонадию или синдром рудиментарных яичек, аплазию герминальных клеток и за развитие стрековых гонад у генетических женщин (Hiort O., Holterhus P., 2000).
Из перечисленных выше хромосом особое значение в процессах развития яичка принадлежит хромосоме 9р, на дистальном коротком плече которой локализуются области, участвующие в функции гена, ответственного за пол. Делеция этой области выявлена у больных с 46XY-гонадальным дисгенезом и обратным полом. У одного из таких пациентов заболевание сочеталось с гонадобластомой (McDonald M. T. [et al.], 1997). В последующем в области хромосомы 9р24.3 была идентифицирована локализация двух генов, участвующих в процессах становления пола (Raymond C. S. [et al.], 1999). Эти гены получили название DMRT1 и DMRT2. Окончательная роль и значение указанных генов в процессах половой дифференцировки устанавливаются, однако показано, что ген DMRT1 экспрессируется только в яичках взрослого человека.
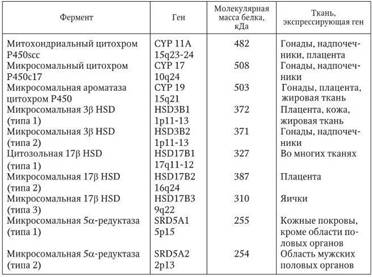
В норме уже к 8-й неделе эмбрионального развития яички способны секретировать половые гормоны, которые и определяют соответствующее развитие наружных половых органов. Дифференцировка наружных половых органов по мужскому типу происходит под влиянием тестостерона, который секретируется клетками Лейдига эмбриона. Последние образуются из интерстиция уже на 9-й неделе эмбрионального периода. Несколько позже эти клетки продуцируют тестостерон, уровень которого достигает максимального значения к 15 – 18-й неделе (Wilson J. D. [et al.], 1995). Уже к середине беременности содержание тестостерона в амниотической жидкости достоверно выше у плодов мужского пола, чем женского. Скорость и количество секретируемого тестостерона определяется активностью 3â-гидроксистероидной дегидрогеназы, которая в тестикулах эмбриона мужского пола в 50 раз выше чем в эмбрионе женского пола. Считается, что в течение первого триместра беременности до начала секреции ЛГ передней долей гипофиза плода образование тестостерона клетками Лейдига может находиться под контролем хорионического гонадотропина человека (ХГЧ), который вырабатывается плацентой. Активаторами тестостерона в тестикулах являются ИФР1 и ингибин, а ингибиторами – α-трансформирующий фактор роста (α-TGF), эпидермальный фактор роста (EGF), фактор роста фибробластов (FGF), активин, ИЛ1, кортиколиберин, вазопрессин, ангиотензин II. Ферменты, участвующие в биосинтезе андрогенов, приведены в табл. 2.1.
Таблица 2.1
Ферменты, участвующие в биосинтезе андрогенов
В клетках-мишенях (урогенитальный синус) должно быть достаточное количество рецепторов к тестостерону, наличием которых и объясняется чувствительность тканей к этому гормону. Гены, контролирующие синтез рецепторов к андрогенам, локализуются на Xq11-12-хромосоме. Ученые установили, что наличие у больных резистентности к андрогенам различной степени выраженности связано с мутациями гена рецептора к андрогенам (Lumbroso S. [et al.], 1992). Рецепторы к андрогенам, как и другие рецепторы к стероидным и тироидным гормонам, состоят из гормонсвязывающего домена, ДНК-связывающего домена и N-концевого домена. Большинство мутаций при андрогенрезистентных состояниях выявлены в гормонсвязывающем домене.
Следует отметить, что в клетках-мишенях (урогенитальный синус и наружные половые органы) дифференцировку урогенитального синуса в наружные половые органы по мужскому типу опосредует не сам тестостерон, а его производная – 5α-дигидротестостерон, который обладает большей биологической активностью. В цитозоле клеток-мишеней содержится фермент 5α-редуктаза (ген 5α-редуктазы расположен на 2-й хромосоме 2р13), конвертирующий тестостерон в дигидротестостерон (ДГТ), который является «чистым» андрогеном и не подвергается воздействию ароматазы и поэтому не может быть конвертирован в эстрогены. Ген 5α-редуктазы экспрессируется в период половой дифференцировки в тканях предстательной железы, урогенитального синуса и наружных половых органов. Выявляют две изоформы 5α-редуктазы. 5α-редуктаза типа 1 (экспрессируется в эпителиальных клетках и проявляет максимальную активность при рН 8,0) и 5α-редуктаза типа 2 (экспрессируется в мезенхимальных андрогенчувствительных и андрогеннечувствительных клетках и проявляет активность при рН 5,5). ДГТ комплексируется с высокоаффинным рецептором цитоплазмы, после чего гормонорецепторный комплекс перемещается в ядро и взаимодействует с местами связывания на хроматине ДНК, что активизирует определенные гены и стимулирует синтез белков. Цитозольный рецептор, связывающий андрогены, регулируется геном, расположенным на Х-хромосоме.
Наряду с тестостероном, большое значение в дифференцировке половых признаков имеет гормон, угнетающий развитие парамезонефрического протока (проток Мюллера), который секретируется клетками Сертоли эмбриона и под влиянием которого происходит инволюция протоков Мюллера, то есть ингибируется их дифференцировка в матку и маточные трубы. Антимюллеров гормон (АМГ) относится к семейству β-трансформирующего фактора роста, к которому также принадлежат активин, ингибин и другие факторы, участвующие в дифференцировке и росте клеток. У человека ген АМГ локализуется на 19-й хромосоме, и его секреция клетками Сертоли осуществляется от периода тестикулярной дифференцировки до пубертата. Примордиальные герминальные клетки являются предшественниками сперматогоний в яичке и овоцитов в яичнике. Дифференцировка бипотенциальной гонады в яичко начинается с процесса миграции примордиальных герминальных клеток на 7-й неделе развития (43 – 50-й день эмбрионального развития, когда длина плода составляет всего 13 – 20 мм). Происходит трансформация интерстициальных поддерживающих клеток в клетки Сертоли, которые затем агрегируют (начинают соединяться друг с другом) и образуют семявыносящие канальцы. Для дифференцировки первичной гонады в яичко необходима также нормальная функция клеток Сертоли, секретирующих антимюллеров гормон, который комплексируется с соответствующими рецепторами. Ген, кодирующий синтез рецепторов к АМГ, локализуется на 12-й хромосоме. При нормальном взаимодействии АМГ с его рецептором и последующих пострецепторных воздействиях у эмбриона мужского пола ингибируется развитие протоков Мюллера, и они подвергаются обратному развитию. Клетки Лейдига дифференцируются из мезенхимы уже на 9-й неделе внутриутробного развития и способны секретировать тестостерон, содержание которого предельно возрастает на 15 – 18-й неделе. Эти клетки имеют рецепторы, связывающие как ЛГ, так и хорионический гонадотропин, под влиянием которого осуществляется биосинтез тестостерона. Секреция тестостерона в период эмбрионального развития имеет прямую корреляцию с уровнем ХГЧ. Лишь позже гипофиз плода начинает секретировать ЛГ, обеспечивая вместе с ХГЧ нормальную секрецию тестостерона, дифференцировку урогенитального синуса, рост и развитие наружных мужских половых органов.
Таким образом, для нормального мужского фенотипа (рис. 2.8) необходимы интактная Y-хромосома с локализацией на ней генов SRY и SOX9, а также нормальная функция Х-хромосомы с наличием на ней гена, ответственного за рецепторы к андрогенам. Для последующей дифференцировки организма по мужскому типу необходимо не только нормальное образование клеток Лейдига и Сертоли (адекватная экспрессия SF1), но и секреция тестостерона – гормона, угнетающего развитие протока Мюллера. Кроме того, необходимо наличие в клетках-мишенях 5α-редуктазы, цитозольных рецепторов к андрогенам в протоках Вольфа, урогенитальном синусе и наружных половых органах, мембранных рецепторов, связывающих фактор, угнетающий развитие протока Мюллера, и, естественно, интактного андрогенрегулируемого гена транскрипции белков. Дефект в любом звене этого механизма приведет к нарушению половой дифференцировки и развитию организма по женскому типу. Необходимо отметить, что под влиянием АМГ к 10-й неделе происходит дегенерация мюллеровых протоков, а к 14-й из вольфовых протоков под контролем тестостерона формируются придатки яичек, семявыносящие протоки и семенные пузырьки (Шабалов Н. П., 2003).
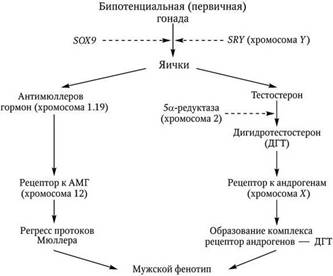
Рис. 2.8. Влияние генетических и гормональных факторов на дифференцировку яичек и формирование мужского фенотипа
Наружные половые органы формируются на 12 – 14-й неделе внутриутробного развития вместе с уретрой и дистальными отделами кишечника. Под влиянием ДГТ, образующегося из тестостерона в клетках полового бугорка и губно-мошоночных складок в присутствии фермента 5α-редуктазы, формируется губчатая часть мочеиспускательного канала и мошонка. Формирование полового члена и мошонки заканчивается к 16 – 17-й неделе гестации.
После 20-й недели происходит миграция яичек из забрюшинной области в мошонку, причем первый этап – опускание яичек до уровня внутреннего пахового кольца – контролирует АМГ, а второй этап – опускание яичек через паховый канал в мошонку – тестостерон. Миграция яичек завершается в основном к 35 – 36-й неделе, а к 38 – 40-й неделе у большинства новорожденных окончательно формируется паховый канал и прерывается сообщение между брюшной полостью и мошонкой.
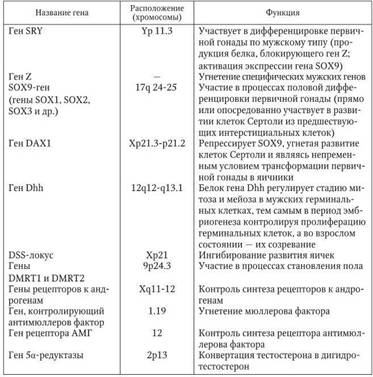
Обобщенные данные по генетическому контролю над дифференцировкой пола представлены в табл. 2.2.
Таблица 2.2
Обобщенные данные по основным участкам генома, контролирующим дифференцировку мужского пола

Еще по теме 2.1. Эмбриогенез и дифференцировка органов репродуктивно-половой системы:
- Органы репродуктивной фазы
- Глава 20 Опухоли половых органов удевочек
- структура и функционирование репродуктивной системы:
- Обработка наружных половых органов роженицы
- Оценка степени дифференцировки опухоли по системе градации Глисона
- Кандидоз наружных половых органов
- 3.4.1.3. ТЕМА: Ультразвуковая диагностика аномалий развития внутренних половых органов у женщин.
- Опухоли женских половых органов
- Опухоли мужских половых органов
- 1. Классификация аномалий развития внутренних половых органов у женщин.
- Эволюция женского организма. Структура и функция репродуктивной системы
- 3.8. Репродуктивная система.
- Раздел 9. Опухоли женских половых органов
- ГЛАВА 11 Клиническая патология мужской и женской репродуктивных систем
- Тубоовариальные образования как осложнения, связанные с инфекционными процессами половых органов. Основные понятия.
- Тубоовариальные образования как осложнения, связанные с инфекционными процессами половых органов. Основные понятия.
- Женская половая система
- Врожденные пороки почек, мочевыводящих путей и половых органов
- Женская половая система
- Женская половая система