Женская половая система
Органы размножения самок иксодовых и аргасовых клещей построены по одному плану (рис. 111—115, стр. 33). Значительные морфологические различия связаны только с характером функциональной дифференцировки влагалища.
Наибольшая расчлененность половой системы — в подсем. АтЫуоттіпае. Так, у самок Hyalomma asiaticum (рис. 116—128) она состоит из непарного яичника и пары яйцеводов, сливающихся в непарную матку. Последняя открывается в эктодермальное влагалище, разделенное на вестибулярный и цервикальный отделы. Последний сообщается с маткой с вентральной стороны мускулистой соединительной трубкой. Дорсо-каудально во влагалище открывается обширный пузыревидный семеприемник. На границе обоих отделов влагалища в него дорсально впадает пара придаточных половых желез. Кроме того, во время кровососания, как и у других Ixodidae, из эпителия вестибулярного отдела формируется лопастевидная придаточная половая железа (рис. 122).В подсем. Ixodinae обособленный семеприемник отсутствует и его функции выполняет расширенный цервикальный отдел (рис. 129). У Аг- gasidae в семеприемник превратилась сильно гппертрофпрованная матка и, кроме того, у них отсутствует лопастевидная железа (рис. 130).
Яичник. У голодных самок иксодовых клещей яичник представляет подковообразно изогнутую трубку. Задняя часть его лежит перпендикулярно к продольной оси тела сразу же позади ректального мешка и вентрально от дивертикулов кишечника. Медиально яичник образует небольшой, направленный вперед выгиб. Боковые части яичника продолжаются вперед и доходят, в зависимости от видовой принадлежности клещей, до уровня II—IV пары кокс, за которыми его дистальные концы делают короткий изгиб назад и открываются в яйцеводы. В подсем. Ixodinae относительные размеры яичника значительно меньше, чем у АтЫуоттіпае. Эти различия особенно выступают с начала кровососания, когда длина яичника увеличивается в несколько раз.
При этом он образует многочисленные изгибы и перед началом яйцекладки занимает большую часть полости тела, тогда как объем дивертикулов кишечника значительно уменьшается.У аргасовых клещей яичник значительно короче, чем у иксодовых, но толще и имеет форму вытянутого мешка, от концов которого отходятяйцеводы (рис.-ИЗ—115). Он также лежит в поперечном направлении позади ректального мешка, но не образует переднелатеральных рукавов. Поверхность яичника, в зависимости от стадии гонотрофического цикла, гладкая или же гроздевидная от массы выступающих над ней зрелых яиц. Исключение составляет лишь Argas vespertilionis (Roshdy, 1961а), у которого яичник парный и образован двумя гроздевидными группами ооцитов, лежащих позади ректального пузыря ближе к его боковым краям (рис. 114). У прошедших несколько гонотрофических циклов старых самок А. persicus яичник также часто выглядит парным, однако обе его половины связаны между собой тонкой полоской соединительной ткани и опорных эпителиальных клеток. Последнее объясняется исчерпанием запаса ооцитов в центральной части органа во время предшествующих яйцекладок. У молодых самок подобной парности яичника нам наблюдать не приходилось.
Яйцеводы. У иксодид яйцеводы представляют пару тонких трубок, отходящих от концов яичника и делающих несколько (чаще три) глубоких петлеобразных изгибов в передне-заднем направлении. Дистальные концы яйцеводов несут слабо выраженные расширения, гомологичные таковым аргазид (рис. 111—112). У АтЫуоттіпае сразу позади места соединения семеприемника с влагалищем яйцеводы соединяются в один непарный орган, называемый маткой. В нее телескопически впячивается соединительная трубка, связывающая матку с цервикальным отделом влагалища (рис. 388).
У Ixodinae яйцеводы также образуют петлеобразные изгибы, а на уровне границы цервикального и вестибулярного отделов влагалища заворачивают назад и вниз. Пройдя небольшое расстояние вдоль боковых стенок влагалища, они круто заворачивают вперед и открываются в матку.
Последняя связана соединительной трубкой с задневентральной частью цервикального отдела (рис. 129).У аргасовых клещей яйцеводы спирально извитые по всему своему ходу, а в их дистальных половинах постоянно наблюдаются ампуловидные расширения. Форма и величина последних зависят как от видовой принадлежности, так и от физиологического состояния клещей. Перед откладкой яиц эти расширения значительно толще, чем у голодных особей. Для аргазид характерны очень крупные размеры матки. Она имеет форму треугольного мешка с заднемедиальным вырезом и лежит между центральной частью средней кишки и вентральной стенкой тела. На дистальном конце перед впадением во влагалище матка обычно сильно сужается. Последний отдел называют шейкой матки (рис. 130).
Влагалище. Наиболее сложно устроенная часть половой системы клещей — влагалище, состоящее из вестибулярного и цервикального отделов. Первый представляет уплощенную трубку, идущую от наружного полового отверстия в заднедорсальном направлении к цервикальному отделу. У иксодид во время питания гиподерма вестибулярного отдела отделяется от кутикулярной выстилки, сильно разрастается и образует массивную лопастевидную придаточную железу (рис. 122).
Цервикальный отдел называют иногда вентральным клапаном (Yal- vac, 1939b) в связи с тем, что в нем соединяются все части половой системы. У АтЫуоттіпае он относительно невелик и с вентральной стороны продолжается в виде мускулистой соединительной трубки, изгибающейся назад и вдвинутой в матку. В заднедорсальную часть цервикального отдела открывается шейка семеприемника, а далее вперед на границе с вестибулярным отделом лежит пара трубчатых придаточных желез. Вентральная стенка вестибулярного отдела вдается в полость влагалища и расширяется на конце, напоминая своими очертаниями сапог, который образует клапан, способный попеременно замыкать доступ в вестибуляр-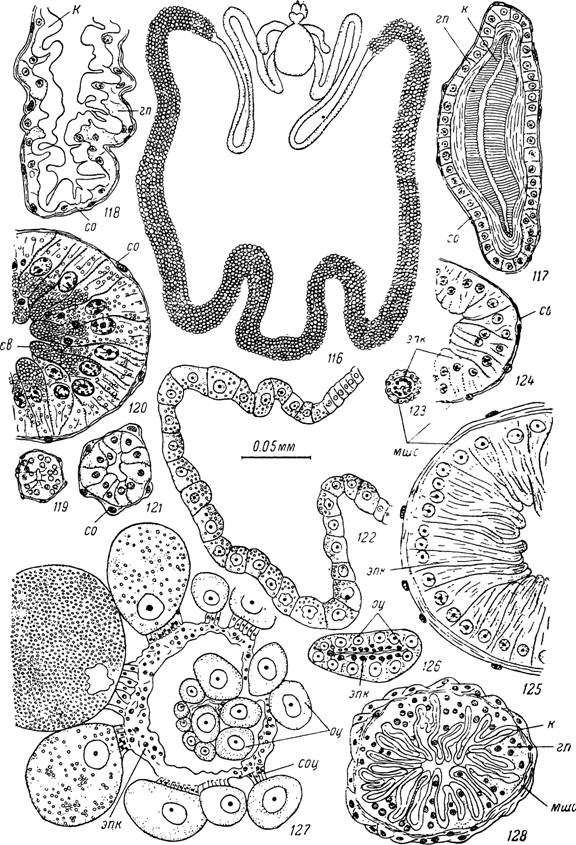
Рис. 116—128. Строение половой системы самки Hyalomma asiaticum.
116 — внешний вид органов размножения напитавшейся особи; поперечные срезы: 117 — вестибулярный и 118 — цервикальный отделы влагалища до начала питания; 119 — трубчатая придаточная железа голодного клеща; 120 — то же напитавшегося клеща; 121 — выводной проток трубчатой железы; 122 — лопастевидная придаточная железа напитавшейся особи; 123 — яйцевод до начала питания; 124 — проксимальный и 125 — дистальный участки яйцевода после питания; 126 — яичник голодного клеща; 127 — яичник напитавшегося клеща; 128 — соединительная трубка.
к — кутикулярная выстилка; гп — гиподерма; со — соединительнотканная оболочка; эпк — епителиальные клетки стенок яичника и яйцеводов; мшс — мышечный слой; оц — ооциты; соц — стебельки ооцитов; св — секреторные вакуоли.ный отдел или соединительную трубку (рис.
388). Семеприемник АтЫуот- тіпае представляет непарный тонкостенный мешок. Он лежит над маткой, а с его боков проходят изгибающиеся вниз парные яйцеводы. Передний конец его сильно сужен и называется шейкой семеприемника. У голодных неоплодотворенных особей стенки семеприемника собраны в многочисленные складки и его размеры относительно невелики. После оплодотворения он сильно растягивается сперматодозами и увеличивается в несколько раз (рис. 116).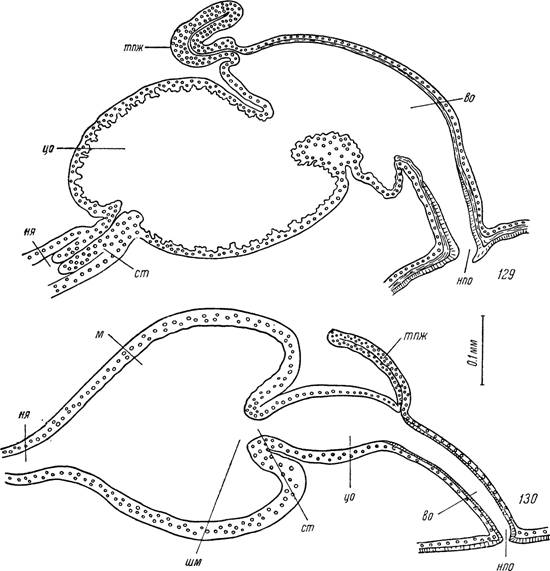
Рис. 129, 130. Сагиттальные срезы.
129 — выводные пути половой системы голодной самки Ixodes ricinus; ISO — то же Ornithodoros papillipes.
нпо — наружное половое отверстие; во — вестибулярный отдел влагалища; цо — цервикальный отдел влагалища; тпж — трубчатая придаточная железа; cm — соединительная трубка; м — матка; шм — шейка матки; ня — непарный яйцевод.
У Ixodinae цервикальный отдел служит семеприемником. Он имеет форму овального мешка, лежит параллельно продольной оси тела и расширяется каудально. С маткой цервикальный отдел соединяется короткой соединительной трубкой. У оплодотворенных особей последняя целиком прикрыта сильно растянутым цервикальным отделом (рис. 129).
У аргасовых клещей цервикальный отдел относительно короток и имеет боченковидную форму. Задний конец его вдвинут внутрь матки и образует соединительную трубку. Размеры цервикального отдела у ар- газид относительно постоянны и он увеличивается только в момент прохождения, яиц или сперматодоз (рис. 130).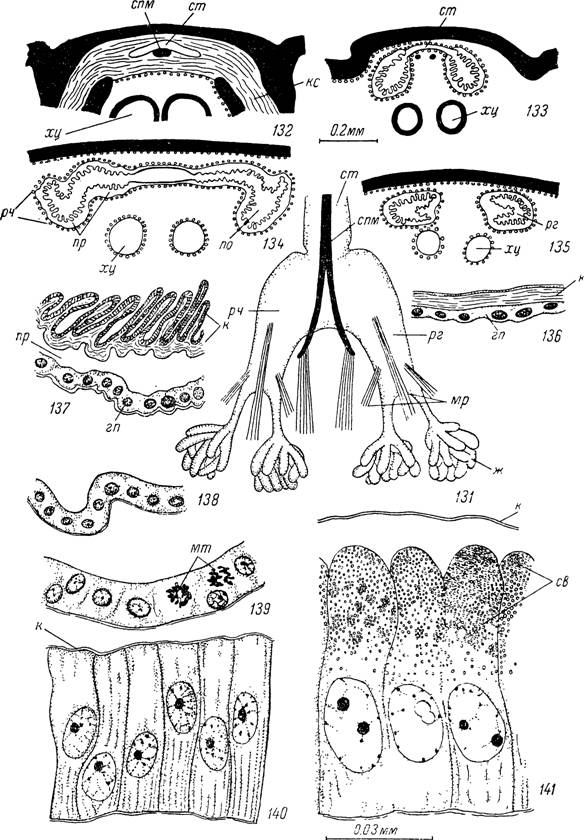
Рис. 131 —141. Орган Жене самки Hyalomma asiaticum.
131 — внешний вид; поперечные срезы на уровне: 132 — камеростомальной складки, 133 — сразу позади камеростомальной складки, 134 — расширенной части органа, 135 — рогов органа; поперечные срезы: 136 — участка стенки стебелька, 137 — расширенной части органа, 138 — железистой ткани рогов голодного, 139 — начавшего питание и 140 — заканчивающего питание клеща, 141 — то же в период яйцекладки.
cm — стебелек; рч — расширенная часть; рг — рога; ж — железы; спм — стержень для прикрепления мышц; мр — мускулы-ретракторы рогов; хц — хелицеры; тес — камеростомальная складка; к — кутикулярная выстилка; гп — гиподерма; по — полость органа; пр — пространство между гиподермой и кутикулярной выстилкой; мт — митотические деления гиподермальных клеток;
св — секреторные вакуоли.Орган Жене.
Важная роль в процессе яйцекладки принадлежит органу Жене (рис. 131—141). Он находится в передней части тела непосредственно под спинным щитком и представляет двух- или четырехлопастный двухслойный мешок, способный выворачиваться наружу через щелевидное отверстие (рис. 131). Последнее у иксодовых клещей расположено медиально в камеростомальной складке между передним краем спинного щитка и заднедорсальным краем основания гнатосомы (рис. 132). У арга- зид щелевидное отверстие находится в камеростомальном углублении.Орган Жене состоит из широкого дорсо-вентрально сплющенного стебелька, расширяющегося назад и расщепляющегося в задней части на два (.Argasidae, АтЫуоттіпаё) или четыре (Ixodinae) рога, и обособленных желез или складок железистой ткани. Отверстие и передняя часть стебелька окружены снаружи очень толстой эндокутикулой камеростомал ьной складки. Пройдя ее, орган сильно расширяется, а кутикулярная выстилка отходит от гиподермы (рис. 133). Внутренний кутикулярный мешок образован тонкими эпикутикулой и эндокутикулой, толщина которых уменьшается в направлении к рогам. В эпикутикуле, начиная с середины стебелька и далее назад, отсутствует поверхностный цементный слой. Между кутикулой и гиподермой находится обширное свободное пространство, заполненное жидкостью (рис. 134, 135).
Задняя часть гиподермального мешка представляет железистое образование. У голодных особей признаки секреторной активности отсут ствуют и дифференцировка желез органа Жене происходит во время кровососания.
У Hyalomma asiaticum существуют две пары желез — передняя и задняя. До начала питания это небольшие карманы складчатой гиподермы на концах рогов и несколько ближе к их основанию. Во время кровососания они вырастают в пучки пальцеобразных желез, сидящих на удлиненных протоках. Последние открываются во внутреннюю полость органа. У Ixodes ricinus на каждом из четырех протоков находится по одной трубчатой железе. У Ornithodoros papillipes, Alueonasus lahorensis и Argas persicus обособленные железы отсутствуют и секреторную функцию, как и у Ornithodoros moubata (Lees a. Beament, 1948), выполняет складчатая гиподерма на концах рогов.
От задней стенки и рогов органа Жене отходят пучки мышц-ретракторов, прикрепляющихся к спинному щитку. Гладкая мускулатура в стенках органа не обнаружена. Под органом Жене от заднего края основания гнатосомы проходит толстый кутикулярный стержень, вилкообразно расщепляющийся на конце (рис. 131). Он служит местом прикрепления ретракторов гнатосомы и при их сокращении, возможно, облегчает выворачивание органа.
Еще по теме Женская половая система:
- Женская половая система
- Женская половая система
- Опухоли женских половых органов
- Раздел 9. Опухоли женских половых органов
- Острой язвой женской половой области Чапина-Липшютца.
- ГЛАВА 11 Клиническая патология мужской и женской репродуктивных систем
- Эволюция женского организма. Структура и функция репродуктивной системы
- Мужская половая система
- Мужская половая система
- Половая система
- Мужская половая система
- Параграф пятый. Застой и загнивание молока в [женской] груди; растяжение иногда случающееся, и ушибы, поражающие [женскую] грудь
- 2.1. Эмбриогенез и дифференцировка органов репродуктивно-половой системы
- Игисинов Нурбек Сагинбекович. эпидемиология рака репродуктивной системы у женского населения Кыргызстана, 2007
- Клинические исследования ангиогенеза в опухолях женской репродуктивной системы и его роль в прогнозе заболевания
- Половая (гендерная) идентичность и половое формирование
- СПЭ потерпевших по делам о половой неприкосновенности и половой свободе личности