Женская половая система
Анатомические и цитологичекие особенности женской половой системы достаточно подробно описаны для иксодин (Roshdy, 1969; Таежный клещ..., 1985) и амблиомин (Brinton, Oliver, 1971а, 1971b; Атлас..., 1979; Kakuda et al., 1995b).
Половая система этих подсемейств имеет сходный план строения, за исключением небольших отличий в анатомии выводных отделов. Сходным образом у всех ик- содид протекают и процессы оогенеза. Женская половая система состоит из непарного яичника, пары яйцеводов, сливающихся в непарный яйцевод, или матку, соединительной трубки и влагалища (рис. 9; табл. XXXI). С влагалищем связаны пара трубчатых придаточных желез, лопастевидная железа, а у амблиомин также семяприемник. В процессах яйцекладки принимает участие орган Женэ.Яичник представляет собой подковообразную трубку, дистальные концы которой открываются в яйцеводы. Он лежит вентрально от дивертикулов кишечника, а его задняя часть расположена перпендикулярно к продольной оси тела над ректальным пузырем. Боковые части яичника продолжаются вперед и доходят до уровня III—IV пары кокс. Снаружи яичник окружен соединительнотканной оболочкой, состоящей из нескольких фибриллярных слоев и неклеточного материала. У голодной самки на поперечном разрезе яичник округлый, с щелевидным центральным просветом. Его стенки образованы генеративными и интерстициальными клетками. Генеративные клетки недавно перелинявшей самки представлены оого- ниями и ооцитами I порядка на разных этапах периода малого роста (I стадия развития). Они округлой или овальной формы, диаметром 10—15 мкм, с крупными ядрами и плотной цитоплазмой. Интерстициальные клетки лежат между генеративными и ограничивают просвет яичника.
У недавно перелинявших особей I. persulcatus (стадия послелиночного дораз- вития) еще продолжаются митотические деления интерстициальных клеток и оого- ниев, но значительная часть ооцитов успевает завершить I стадию развития.
У голодающих самок часть ооцитов успевает вступить в начало цитоплазматической стадии большого роста (И стадия развития), и они слегка выступают над поверхностью яичника.Стимулом к возобновлению процессов оогенеза служит начало питания клеща (рис. 22; табл. XXXII, 7). Уже через 1—2 сут прикрепления значительная часть ооцитов заканчивает II стадию развития и достигает диаметра в 60—80 мкм. На стадии цитоплазматического роста в ооците завершается формирование клеточных органоидов, почти отсутствовавших на стадии малого роста. В цитоплазме возрастает количество митохондрий, появляются многочисленные удлиненные цистерны гранулярного эндоплазматического ретикулума и комплексы Гольджи. Диаметр ядра достигает 20—25 мкм, а сильно увеличившееся в размерах ядрышко приобретает сложную структуру; сформированный на этой стадии рибонуклеопротеиновый материал выводится в цитоплазму через поры ядерной оболочки и образует около- ядерные скопления (табл. XXXII, 5). Проникновение питательных веществ в развивающиеся ооциты облегчается развитием особой структуры — ножки. Она образована из видоизмененных интерстициальных клеток (табл. XXXII, 2). В зоне соединения этих клеток с ооцитом с обеих сторон формируются хорошо развитые микроворсинки, что служит признаком активного транспорта веществ.
Следующий период большого роста ооцитов (III—IV стадии) связан с образованием желточных гранул (трофоплазмати%еский рост). Стимулом для его начала служит осеменение клещей, и в его отсутствие эта стадия у питающихся самок не наступает (Coons et al., 1989). Формирование предшественника желтка — вителлина происходит в специализированных желточных клетках кишечника и клетках жирового тела. Предшественники желтка поступают из гемолимфы через легко проницаемую для белковых молекул соединительнотканную оболочку яичника. Внутрь развивающегося ооцита вителлогенин проникает путем эндоцитоза межд\ микроворсинками на его поверхности. В цитоплазме яйцеклеток в этот период появляется большое количество мелких гранул, которые, вероятно, являются желточными.
К концу III стадии развития диаметр ооцитов достигает 200—250, ядер — 40—50 мкм.
На IV стадии развития ооцитов доминируют процессы экзогенного накопления желтка. Экзогенные и эндогенные компоненты желтка сливаются в крупные желточные шары диаметром 20—30 мкм. Они вместе с многочисленными каплями липидов заполняют большую часть цитоплазмы и частично маскируют ядро. На этой стадии заканчивается рост ооцитов, достигающих диаметра около 700 мкм.Поглощение белков из гемолимфы прекращается с формированием наружной желточной оболочки ооцита (V стадия развития), неправильно называемой хорионом. Желточная оболочка имеет кутикулярную природу и образуется исключительно самим ооцитом. Фрагменты оболочки закладываются между микроворсинками в непосредственном с ними контакте (табл. XXXII, 4). В конце вителлогенеза ооцит соединяется с клетками ножки десмосомоподобными структурами. После окончательного формирования желточной оболочки яйцо под действием натяжения соединительнотканной оболочки яичника раздвигает клетки ножки и выталкивается в полость яичника.
Развитие ооцитов происходит неодновременно, чем обусловлен, в частности, многодневный период откладки яиц. У голодной самки вдоль всего яичника проходит так называемая продольная борозда. Последняя состоит из оогониев и ооцитов, остановившихся на I стадии _рдзвития, тогда как на поверхности яичника уже выступают многочисленные яйцеклетки, достигающие конца II стадии развития (табл. XXXII, 7). Во время питания различия в темпах развития между ооцитами внутри продольной борозды и на поверхности яичника увеличиваются. В результате у полностью напитавшейся самки (табл. XXXI, 2) в борозде заключены ооциты I и II стадий, тогда как на поверхности яичника находятся ооциты, завершающие вителлогенез (III—IV стадии).
С начала яйцекладки (табл. XXXI, 3) по мере расходования запаса наиболее продвинутых в развитии зрелых ооцитов на поверхности яичника ускоряется раз-
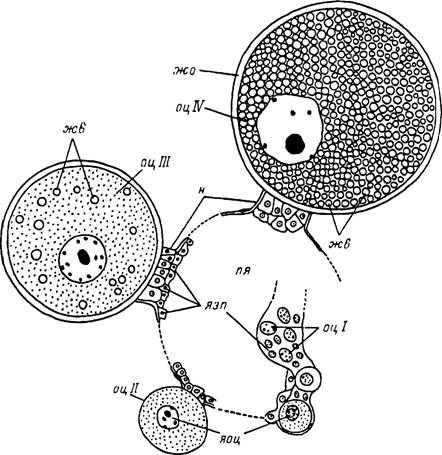
Рис. 22. Развитие яйцеклеток в яичнике питающейся самки иксодового клеща.
же — желточные включения; жо — желточная оболочка; н — ножка; оц 1 — ооцит I стадии внутри продольной борозды стенки яичника; оц II— ооцит II стадии в начале периода большого роста; оц III — ооцит III стадии в начале заполнения цитоплазмы желточными включениями; оц IV—ооцит IV стадии со сформировавшейся желточной оболочкой; пя — полость яичника; яоц — ядро ооцита; лэп — ядро эпителиальной клетки стенки яичника.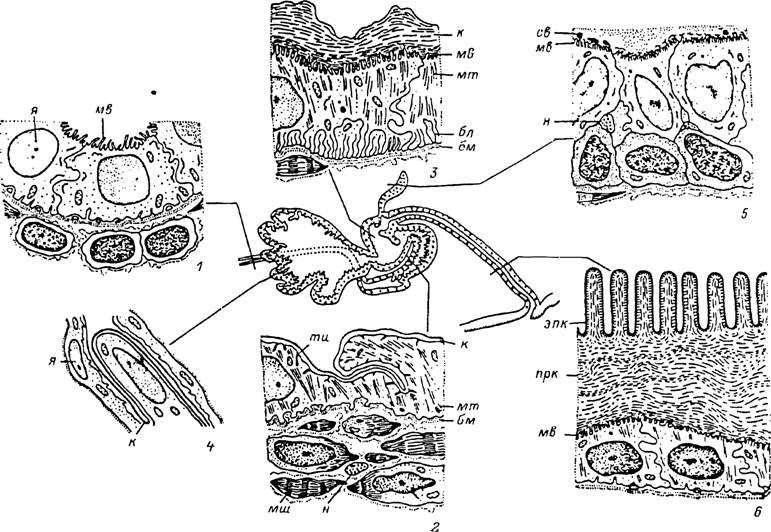
Рис.
23. Строение выводящих путей половой системы самки Hyalomma asiaticum.1 — яйцевод; 2 — соединительная трубка; 3 — цервикальный отдел влагалища; 4 — семяприемник; 5 — придаточная железа; 6 — вестибулярный отдел влагалища, бл — базальный лабиринт; бм — базальная мембрана; к — кутикулярная выстилка; мв — микроворсинки; мт — микротрубочки; мш — мышечные волокна; н — нерв; прк — прокутикула; св — секреторные включения; ти — трубчатая инвагинация; эпк — эпикутикула; я — ядро.зитие ооцитов продольной борозды. Последняя постепенно расправляется, и заключенные в ней яйцеклетки занимают поверхностное положение, завершают большой рост и овулируют. У заканчивающих яйцекладку и погибающих самок в гтенках яичника остаются лишь единичные ооциты I—IV стадий, по тем или иным причинам остановившиеся в своем развитии. Яичник сильно уменьшается, і на поверхности сморщившейся соединительнотканной оболочки от закончивших развитие ооцитов сохраняются многочисленные пенькообразные выросты — остатки ножек (табл. XXXI, 4).
У голодающих самок внутри яичника в ооцитах одновременно протекают как митотические, так и мейотические процессы. Ооциты, образующиеся при размножении оогониев, минуя период интеркинеза, сразу же переходят в профазу первого деления созревания. Однако ядра развивающихся ооцитов достигают лишь стадии іиплотены, а основная их масса остается у голодной самки на более ранних стадиях мейоза. Уже в начале диплотенной стадии биваленты деспирализуются и з дальнейшем с трудом обнаруживаются в ядре по отдельным гетерохроматиновым ♦часткам.
На II стадии развития в сильно Увеличившихся ядрах ооцитов хромосомы по- лрежнему находятся в деспирализованном состоянии. Переход на стадию диакинеза наблюдается только в конце развития ооцита перед самой овуляцией. Хромосомы зновь образуют характерные диакинетические биваленты, ядерная оболочка растворяется, а биваленты тесно смыкаются и формируют одно компактное тело, называемое кариосферой, сохраняющееся до оплодотворения яйцеклетки.
Оплодот- зорение (сингамия) происходит после овуляции ооцита во время его движения по ійиеводам или даже в полости яичника, куда также проникают спермии. Слияние мужского и женского ядер происходит уже после откладки яйца. Таким образом, оба периода — большого роста и отложения желтка протекают в ооцитах I порядка.Яйцеводы представлены парой тонких трубок, которые отходят от концов зичника вперед, а затем на уровне передней трети влагалища круто поворачивают назад и впадают в заднебоковые углы общего яйцевода, или матки (рис. 23). Диаметр яйцеводов у голодных самок составляет 15—30, а перед впадением в матку — 25—50 мкм. На поперечном разрезе они почти круглые, с небольшим центральным просветом. Стенки яйцеводов построены из слоя столбчатых эпителиальных клеток высотой 7—10 мкм, окруженных снаружи соединительнотканной оболочкой.
С началом питания наблюдаются многочисленные деления эпителиальных клеток и увеличение их размеров до 30—50 мкм в дистальных и 50—90 мкм в проксимальных участках яйцеводов; соответственно диаметр яйцеводов увеличивается до 60—70 и 100—150 мкм. Апикальные концы клеток, особенно в дистальных частях, приобретают сложную конфигурацию и вдаются в просвет. На их поверхности густо расположены микроворсинки, а базальные концы клеток образуют лабиринт. В цитоплазме формируются многочисленные цистерны эндоплазматического ретикулума и комплексы Гольджи. Клетки стенок яйцевода приобретают железистую функцию. В апикальных концах клеток и в просвете яйцевода появляются гранулы секрета полисахаридной природы. Между апикальными концами клеток в полости яйцеводов оплодотворенных питающихся самок обнаруживаются многочисленные спермии. Часть спермиев фагоцитируется или каким-то другим способом попадает внутрь клеток стенок яйцеводов (Соколов, 1956), где они подвергаются лизису в многочисленных фаголизосомах. В наружной соединительнотканной оболочке яйцеводов питающихся самок дифференцируется хорошо развитый мышечный слой. Последний осуществляет перистальтические сокращения ійцеводов, вследствие которых яйца передвигаются к влагалищу.
Матка иксодин представляет собой треугольный мешок, вершина которого направлена вперед и вдвинута в заднедорсальную часть влагалища, с которым она сообщается соединительной трубкой (рис.
9). Гистологическое строение матки то же, что и у яйцеводов (табл. XXXIII, 2). Эпителиальные клетки ее стенок уплощенные, а вблизи впадения яйцеводов, в заднедорсальной части органа, — столбчатые. Снаружи матка окружена соединительнотканной оболочкой с тонким мышечным сплетением. Ее полость у оплодотворенных особей содержит плотную массу спермиев, проникающих из влагалища. У амблиомин матка меньших размеров, чем у иксодин, и расположена вентрально по отношению к влагалищу (рис. 23).Соединительная трубка эктодермального происхождения представляет продолжение влагалища и у иксодин глубоко вворачивается в его внутреннюю полость. Снаружи она окружена мощно развитым слоем мускулатуры, а изнутри выстлана гиподермальными клетками, которые образуют глубокие складки, расправляющиеся при прохождении яиц. Апикальная поверхность этих клеток покрыта тонкой складчатой кутикулой (табл. XXXIII, 3).
Влагалище состоит из цервикального и вестибулярного отделов. У иксодин обширный мешковидный цервикальный отдел вместе с маткой выполняет функции семяприемника (рис. 9). В полости вестибулярного отдела (табл. XXXIII, 1) заключены целые сперматодозы, их пустые оболочки, а в заднем конце вблизи от соединительной трубки может лежать масса из свободных спермиофоров. В передней части цервикального отдела, частично прикрывающей у оплодотворенных особей вестибулярный отдел, может находиться скопление слизистого материала. Снаружи цервикальный отдел окружен мышечным сплетением, а изнутри выстлан гиподермальными клетками, несущими железистую функцию. Апикальные концы клеток покрыты микроворсинками, а базальные формируют лабиринт. Поверхность клеток покрыта складчатой кутикулярной выстилкой (табл. XXXIII, 2).
У Я. asiaticum и других амблиомин цервикальный отдел небольших размеров. От него в заднедорсальном направлении отходит тонкостенный мешок, называемый семяприемником. Стенки последнего собраны в глубокие складки, снаружи выстланные тонкой кутикулой. После поступления в семяприемник сперматофора стенки семяприемника растягиваются и складки расправляются. Клетки стенок увеличиваются в размерах и становятся секреторными. Именно в семяприемнике амблиомин проспермии завершают свой спермиогенез и превращаются в зрелые спермии.
Вестибулярный отдел. Короткий трубчатый вестибулярный отдел отходит почти вертикально от наружного полового отверстия и сообщается с вентральной стороны с цервикальным отделом. В месте их соединения формируется кутикулярный, сильно складчатый вырост, возможно, выполняющий клапанную функцию. Кутикулярная выстилка влагалища собрана в складки, отличается значительной толщиной и напоминает кутикулу покровов тела, продолжением которой она является (табл. XXXIII, 7). Снаружи вестибулярный отдел окружен мышечным слоем. При откладке яйца стенки вестибулярного отдела частично выворачиваются наружу и он выполняет функции яйцеклада.
Придаточные железы. С началом питайия гиподерма вестибулярного отдела отходит от кутикулы, приобретает железистую функцию и образует многочисленные складки лопастевидной придаточной железы. Рост железы осуществляется за счет интенсивного деления клеток. В результате перед началом яйцекладки кутикулярная трубка вестибулярного отдела соединяется с гиподермой лишь на границе с половой щелью и цервикальным отделом, а на большем протяжении она окружена обширной полостью вновь образовавшейся железы (табл. XXXIII, 3). Клетки этой железы достигают высоты 40—80 мкм, и в их апикальной части появляются многочисленные эозинофильные и липоидные секреторные вакуоли. Подобный же секреторный материал накапливается и в полости железы.
Трубчатая придаточная железа у голодных самок иксодид представлена парой коротких, слегка извитых трубок диаметром 20—40 мкм, открывающихся в заднебоковые участки вестибулярного отдела. Наиболее детально строение этих органов описано у Я. longicornis (Kakuda et al., 1994). Стенки железы построены из нескольких рядов плотно прилегающих друг к другу овальных ядер будущих железистых клеток, разделенных тонкими прослойками цитоплазмы. Просвет железы на этой стадии развития не выражен.
Во время питания трубчатые железы увеличиваются в размерах в несколько раз, и у готовых к яйцекладке самок их диаметр достигает 200 мкм. Клетки, и в первую очередь граничащие с просветом железы, начинают выполнять секреторные функции. Они приобретают пирамидальную форму высотой в 70—80 мкм. Ядра их значительно увеличиваются, а в цитоплазме хорошо развиты органоиды, ответственные за образование секрета. Апикальные концы клеток, выступающие в просвет, покрыты многочисленными невысокими микроворсинками, а цитоплазма богата секреторными включениями белковой природы (табл. XXXIII, 4). Между клетками железы имеются нервные окончания, контролирующие секреторную активность органа.
Во время яйцекладки полость вестибулярного отдела влагалища бывает заполнена слизистой массой, вероятно представляющей секрет трубчатых и лопастевидных придаточных желез. В процессе яйцекладки этот секрет, как и липоидный секрет органа Женэ, обволакивает*“Іяйца снаружи водонепроницаемым слоем и, кроме того, обеспечивает склеивание яиц между собой.
Орган Женэ, хотя анатомически и не связан с женским половым аппаратом, играет важную роль в процессе откладки яиц. Он находится в передней части тела непосредственно под спинным щитком и представляет у иксодин четырехлопастный, а у амблиомин двухлопастный двухслойный мешок, способный в момент откладки яиц выворачиваться наружу через щелевидное отверстие в камеростомал ьной складке. Вворачивание органа осуществляют пучки мышц-ретракторов, которые прикрепляются к спинному щитку.
Внутренняя полость органа выстлана тонкой кутикулой, способной выворачиваться наружу. Между кутикулой и гиподермой заключено обширное свободное пространство, заполненное жидкостью. У голодных особей признаки секреторной активности отсутствуют. Собственно железы дифференцируются только во время питания в виде пальчатых выростов концов рогов. Секрет поступает в пространство между гиподермой и кутикулой и далее при откладке яиц проникает на поверхность выворачивающихся рогов. В момент соприкосновения с рогами органа Женэ на поверхность яйца изливается и секрет желез поровых полей.
Еще по теме Женская половая система:
- ТЕМА № 1 ОРГАНИЗАЦИЯ РАБОТЫ ЖЕНСКОЙ КОНСУЛЬТАЦИИ ДИСПАНСЕРНОЕ НАБЛЮДЕНИЕ БЕРЕМЕННЫХ
- Эволюция женского организма. Структура и функция репродуктивной системы
- Классификация заменителей женского молока
- Женская половая система
- Мужская половая система
- Введение ...................................................................................................................... 5
- Иксодовые клещи.
- Женская половая система
- ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАЗМНОЖЕНИЯУ САМОК
- Введение
- Цепень вооруженный, или свиной солитер (Taenia solium)
- Лентец широкий (Diphyllobothrium latum)
- Тараканы
- КЛАСС СОСАЛЬЩИКИ (TREMATODA)
- Возбудитель энтеробиоза
- Половая (гендерная) идентичность и половое формирование
- 2.2.1. Профилактические осмотры женского населения и выявление гинекологических заболеваний.
- Содержание
- Женская половая система
- Нарушение функций половых желез