Количественные аспекты.
Взаимоотношения между метаболитами и антиметаболитами обычно имеют конкурентный характер, т. е. если х молекулам метаболита противодействует у молекул аналога, то для достижения того же биологического эффекта 10х молекулам метаболита потребуется 10у молекул аналога.
Поскольку такие конкурентные реакции полностью- обратимы, антагонистическое действие, оказываемое у молекулами аналога на х молекул метаболита, может быть преодолена введением других х молекул метаболита и так далее. Конкурентные отношения этого типа наблюдаются между малоново» и янтарной кислотами, стрептоцидом (сульфаниламид) и пара- аминобензойной кислотой.Для каждой пары веществ существует свой специфический индекс ингибирования, определяемый как соотношение числа молекул аналога к числу молекул метаболита, при котором достигается 50% ингибирование. Это соотношение имеет разные значения для различных биологических видов, но она всегда одинаково для каждого данного вида. Оно характеризует относительное сродство аналога и метаболита к рецептору и, кроме того, отражает различия в способности двух веществ проникать к месту действия, особенно тогда, когда это место труднодоступно. Поэтому уровень ингибирования, вызванного аналогом, определяется двумя факторами: во-первых, относительным сродством аналога и метаболита к рецептору и, во-вторых, относительными количествами аналога и метаболита в месте реакции.
Соодство субстрата к ферменту характеризуется уравнением:
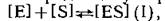 где [Е—концентрация фермента, [S]—концентрация субстрата, a [ES]—концентрация комплекса, который они образуют. Естественно, сродство лекарственного препарата, являющегося ингибитором (I) фермента, может быть описано аналогичным выражением и тогда константа ингибирования (Ki), т. е. константа диссоциации комплекса фермент — ингибитор будет представлена уравнением:
где [Е—концентрация фермента, [S]—концентрация субстрата, a [ES]—концентрация комплекса, который они образуют. Естественно, сродство лекарственного препарата, являющегося ингибитором (I) фермента, может быть описано аналогичным выражением и тогда константа ингибирования (Ki), т. е. константа диссоциации комплекса фермент — ингибитор будет представлена уравнением:

В отличие от ингибитора субстрат под действием фермента превращается в продукт (Р) и последовательность событий становится: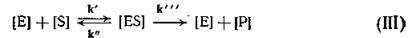
Соотношение констант скоростей к'/к" есть ничто иное, как константа Михаэлиса — Ментен (Км).
Она отражает диссоциацию комплекса фермент — субстрат [ES] на компоненты [Е] и [S] и может быть представлена равенством (IV), формально аналогичным приведенному выше для Ki(II):
22
Индекс ингибирования, характерный для каждой пары ингибитор — метаболит, — это соотношение Кі ■ Km — чем ниже его величина, тем эффективнее ингибитор.
Константа Михаэлиса, очевидно, обратно пропорциональна сродству фермента к субстрату и численно равна концентрации субстрата в тот момент, когда скорость реакции достигает половины максимальной. Размерность величин Кі и Кт выражается в г-моль/л. Однако это не истинные константы равновесия, а отношения констант скоростей прямой и обратной реакций фермента с субстратом или ингибитором. Наиболее удобно определять Кт графическим методом Lineweaver, Burk (1934), согласно которому строится график зависимости обратной величины начальной скорости образования комплекса [ES] от обратных величин концентрации субстрата. На графике должна получиться прямая линия, при этом точка пересечения полученной прямой с осью абсцисс соответствует величине Кт.
Поскольку интенсивные метаболические процессы в клетках гораздо чаще приводят к «стационарному» состоянию, чем к равновесному, целесообразно использовать уравнение Бриггса — Холдейна [Briggs — Haldane, 1925], которые показали, что совсем необязательно предполагать наличие равновесия между [Е] и [S]. Они вывели уравнение (V), формально аналогичное уравнению Михаэлиса — Ментен, но предполагающее условия стационарного состояния, а не равновесия.

Константа Кьь, имеющая более общий характер, чем предыдущая, является показателем степени одновременной диссоциации комплекса [ES], протекающей в двух противоположных направлениях. Если Кт заменить на Кьь, то индекс ингибирования становится равным соотношению Кі : Кьи- При проведении этих расчетов предполагается, что ни ингибитор, ни его комплекс с ферментом не разрушаются даже частично под действием других биологических веществ, которые могут присутствовать при реакции.
Молярный индекс ингибирования малоновой кислоты равен >/з [Thron, 1953]. Еще более эффективно ингибирует оксид углерода, когда замещает кислород в гемоглобине (1/210). Величины индекса ингибирования меньше единицы необычны, потому что очень редко аналог обладает большим сродством к природному рецептору, чем нормальный субстрат. Беспрецедентно высокий индекс ингибирования (1/10 000) для метотрексата (4.7) обусловлен, вероятно, тем, что самым основным атомом азота в молекуле этого ингибитора является N-1, в отличие от дигидрофолиевой кислоты, у которой таковым служит N-5. Очевидно, в этом заключена причина столь больших различий в соответствии метотрексата и дегидрофолиевой кислоты месту связывания их с рецептором (разд. 4.2 и 9.3.3). Такие индексы
ингибирования как у метотрексата (1/10 000)—предел обычно наблюдаемых для лекарственных веществ. Индексы ингибирования, величины которых лежат выше единицы, такие как, например, сульфаниламида по отношению к ПАБ (300/1) — другой предел, ниже которого не следует ожидать высокого терапевтического эффекта аналога.
Почему желательны минимальные величины индексов ингибирования для лекарственных препаратов? Потому что эволюция обусловила появление ферментов, использующих соответствующие природные субстраты с максимальной эффективностью. Другая причина относительно малой эффективности многих конкурентных ингибиторов — стационарное состояние живой клетки. Атакуемый фермент постоянно снабжается субстратом (от предыдущего фермента в цепи), а его продукт непрерывно удаляется последующим ферментом. Фермент, атакуемый ингибитором, обычно недостаточно насыщен субстратом и поэтому имеет большую резервную активность. При введении конкурентного ингибитора в стационарную систему целевой фермент сначала ингибируется, но по мере накопления нереагирующего нормального субстрата это ингибирование фермента прекращается. Таким образом, возникает новое стационарное состояние, в котором общий итог действия данного метаболического пути остается неизменным, поскольку концентрация субстрата для атакуемого фермента поддерживается на более высоком уровне с тем, чтобы уравновесить эффект ингибирования [Cleland, 1970]. В качестве примера можно привести различия между химией модельных систем, изучаемых в лаборатории, и более сложных систем, образовавшихся в живой клетке в процессе эволюции.
Еще по теме Количественные аспекты.:
- Количественные аспекты связывания металлов
- 2. Количественные ультразвуковые характеристики нормальной молочной железы в возрастном аспекте.
- 3.4.2. Анализ ассоциаций исследованных генов с количественными признаками туберкулеза
- Количественный подход
- 6. Наследственность и среда – основные факторы количественной изменчивости.
- Количественная (квантитативная) периметрия.
- Количественная оценка ионизирующих излучений. Основы дозиметрии
- Количественное сенсорное тестирование (KCT)
- 25. Количественные и качественные характеристики социальных отклонений.
- Количественные и качественные изменения лейкоцитов в крови
- Качественные и количественные нарушения фолликулогенеза в яичнике
- Количественные пробы
- Количественное определение суммы флавоноидов в препаратах (настое и настойке) полыни эстрагон
- Количественное определение содержания суммы флавоноидов в траве полыни эстрагон методом спектрофотометрии
- 1.4.3. Количественный анализ в КТ легких
- 26. Возможности количественной оценки здоровья.
- Результаты количественного SWOT- анализа
- Количественное исследование слуха камертонами,
- Количественное выражение вероятностных утверждений
- Определение количественных показателей гемодинамики печени.