юМРХЛЕРЮАНКХРШ: НОПЕДЕКЕМХЕ, ОПНХЯУНФДЕМХЕ Х ЛЕУЮМХГЛ ДЕИЯРБХЪ
аХНКНЦХВЕЯЙХЕ ЩТТЕЙРШ ЛЕРЮАНКХРНБ (ЯСАЯРПЮРНБ ХКХ ЙН╜ТЕПЛЕМРНБ), МЮУНДЪЫХУЯЪ Б МЕГМЮВХРЕКЭМШУ ЙНКХВЕЯРБЮУ Б ЙКЕР╜ЙЕ ХКХ РЙЮМХ, ЛНЦСР АШРЭ ОНДЮБКЕМШ ДЕИЯРБХЕЛ ХУ ЮМЮКНЦНБ, МЮГШБЮЕЛШУ ЮМРХЛЕРЮАНКХРЮЛХ.
б ЛНКЕЙСКЕ ЙЮФДНЦН ХГРЮЙХУ ЮМЮКНЦНБ ХЛЕЕРЯЪ СВЮЯРНЙ, ОНДНАМШИ СВЮЯРЙС ЛЕРЮАНКХРЮ, НАЕЯОЕВХБЮЧЫЕЛС БГЮХЛНДЕИЯРБХЕ ЮМЮКНЦЮ Я ТЕПЛЕМРМШЛ АЕК╜ЙНЛ. дКЪ РНЦН ВРНАШ ЮМЮКНЦ АШК ЩТТЕЙРХБМШЛ, НМ ДНКФЕМ ХЛЕРЭ ЯУНДЯРБН Я ЛЕРЮАНКХРНЛ МЕ РНКЭЙН Б ПЮГЛЕПЮУ, МН Х Б ПЮЯ╜ОПЕДЕКЕМХХ ЩКЕЙРПНМНБ, РЮЙ ЙЮЙ ЮЙРХБМШЕ ЖЕМРПШ ТЕПЛЕМРНБ, ЙЮЙ ОПЮБХКН, БШЯНЙНОНКЪПМШ. юМРЮЦНМХГЛ ЛЕФДС ЮМЮКНЦНЛ Х ЛЕРЮАНКХРНЛ НАСЯКНБКЕМ РЕЛ, ВРН ЮМРХЛЕРЮАНКХР ГЮМХЛЮЕР Х АКНЙХПСЕР ЮЙРХБМШЕ СВЮЯРЙХ ТЕПЛЕМРЮ, НАШВМН ХЯОНКЭГСЕЛШЕ ЛЕРЮАНКХРНЛ (ПЮГД. 9.2).
уНРЪ ЛЕРЮАНКХР ЛНФЕР АШРЭ ОПЕБПЮЫЕМ Б ЮМЮКНЦ ХГЛЕМЕМХЕЛ УХЛХВЕЯЙНИ ЯРПСЙРСПШ ЛНКЕЙСКШ, ЩРН ХГЛЕМЕМХЕ МЕ ДНКФМН АШРЭ ГМЮВХРЕКЭМШЛ, ХМЮВЕ ЛЕРЮАНКХР ОНРЕПЪЕР АХНКНЦХВЕЯЙНЕ ДЕИЯРБХЕ, МН МЕ ОПХНАПЕРЕР ЮМРЮЦНМХЯРХВЕЯЙНЦН. рЮЙ ПЮГМННАПЮГ╜МШЕ ЛНДХТХЙЮЖХХ ЯРПСЙРСПШ РХЮЛХМЮ ОСРЕЛ ОПХЯНЕДХМЕМХЪ ДНОНКМХРЕКЭМНИ ХКХ НРЫЕОКЕМХЪ ХЛЕЧЫЕИЯЪ ЛЕРХКЭМНИ ЦПСООШ ЯМХФЮЧР ЕЦН ЮЙРХБМНЯРЭ ЙЮЙ БХРЮЛХМЮ, МН МЕ ДЕКЮЧР ЕЦН ЮМРЮ╜ЦНМХЯРНЛ (ЦКЮБЮ 2). б ЖЕКНЛ, ОНРЕПЪ ХКХ ОПХНАПЕРЕМХЕ ЛЕРХКЭ╜МНИ ЦПСООШ ЛНКЕЙСКНИ ЛЕРЮАНКХРЮ ≈ ЯКХЬЙНЛ АНКЭЬНЕ ХГЛЕМЕ╜МХЕ, ВРНАШ НМ ЯРЮК ЮМРЮЦНМХЯРНЛ, ОН ЙПЮИМЕИ ЛЕПЕ Б ЯКСВЮЕ МЕАНКЭЬХУ ЛНКЕЙСК, Б ЙНРНПШУ ХГЛЕМЕММШИ СВЮЯРНЙ НАШВМН МЮ╜УНДХРЯЪ БМСРПХ ХКХ НВЕМЭ АКХГЙН Й ЮЙРХБМНЛС СВЮЯРЙС ТЕПЛЕМ╜РЮ. жЕКЕЯННАПЮГМЕЕ ЯНУПЮМЪРЭ ЯРЕПХВЕЯЙХЕ ЯБНИЯРБЮ ЛЕРЮАНКХРЮ (ВРНАШ НАЕЯОЕВХРЭ ЯБЪГЭ Я ТЕПЛЕМРНЛ), МН БЮПЭХПНБЮРЭ ЕЦН ЩКЕЙРПНММШЕ ЯБНИЯРБЮ (ВРНАШ ЯДЕКЮРЭ МНБНЕ БЕЫЕЯРБН МЕ ОНД╜УНДЪЫХЛ ДКЪ ТЕПЛЕМРЮ Б ЙЮВЕЯРБЕ ЯСАЯРПЮРЮ).
мЕЙНРНПШЕ ЮМРЮЦНМХЯРШ ХЛЕЧР РЮЙСЧ ОПНЯРСЧ УХЛХВЕЯЙСЧ ОПХПНДС, ВРН ХУ ЯОНЯНАМНЯРЭ БШЯРСОЮРЭ Б ПНКХ ЮМЮКНЦНБ ЛЕ╜РЮАНКХРНБ ВЮЯРН НЯРЮЕРЯЪ МЕГЮЛЕВЕММНИ. мЮОПХЛЕП, НДМХ МЕНПЦЮ╜МХВЕЯЙХЕ ЙЮРХНМШ ЙНМЙСПХПСЧР Я ДПСЦХЛХ (ПЮГД. 9.2 Х 11.0). дЮФЕ ХНМ БНДНПНДЮ (ОН СРБЕПФДЕМХЧ лХРВЕККЮ[1] НДХМ ХГ МЮХ╜АНКЕЕ БЮФМШУ ЛЕРЮАНКХРНБ) СВЮЯРБСЕР Б ЙНМЙСПЕМЖХХ Я НПЦЮМХ╜ВЕЯЙХЛХ Х МЕНПЦЮМХВЕЯЙХЛХ ЙЮРХНМЮЛХ (ПЮГД.
10.3.1) [Mitchell, 1979]. щРХКНБШИ ЯОХПР, БУНДЪЫХИ Б ЯНЯРЮБ ЮКЙНЦНКЭМШУ МЮОХР╜ЙНБ, ДЕИЯРБСЕР ВЮЯРХВМН ГЮ ЯВЕР ЙНМЙСПЕМЖХХ Я БНДНИ (ОН ЙПЮИ╜МЕИ ЛЕПЕ ОПХ ПЮЯОПЕДЕКЕМХХ). нМ ЛНФЕР РЮЙФЕ ЯМХЛЮРЭ РНЙЯХ╜ВЕЯЙНЕ ДЕИЯРБХЕ ЛЕРХКНБНЦН ЯОХПРЮ, БШРЕЯМЪЪ ЕЦН Я НЙХЯКЪЧЫЕЦН ТЕПЛЕМРЮ [Roe, 1955].хГБЕЯРЕМ ПЪД ОПХЛЕПНБ ЙНМЙСПЕМЖХХ ЛЕФДС ОПНЯРШЛХ ЮМХН╜МЮЛХ. рЮЙ, ОЕПУКНПЮР- Х РХНЖХЮМЮР-ЮМХНМШ ХМЦХАХПСЧР МЮЙНО╜КЕМХЕ ИНДХД-ХНМЮ Б ЫХРНБХДМНИ ФЕКЕГЕ АЕГ ЯЙНКЭЙН-МХАСДЭ ГЮЛЕРМНЦН МЮПСЬЕМХЪ НЙХЯКХРЕКЭМНЦН БЙКЧВЕМХЪ ИНДХД-ХНМЮ Б РХПНЙЯХМ [Stanbury, Wyngaarden, 1952]. рЮЙХЛ ФЕ НАПЮГНЛ ЯОН╜ЯНАМНЯРЭ АЮЙРЕПХИ Nitrobacter НЙХЯКЪРЭ МХРПХРШ Б МХРПЮРШ ХМ╜ЦХАХПСЕРЯЪ ЖХЮМЮР- ХКХ ОЕПУКНПЮР-ЮМХНМЮЛХ: ЩТТЕЙР ОНЯКЕДМХУ КЕЦЙН ЯМХЛЮЕРЯЪ ОПНЯРНИ НРЛШБЙНИ [Lees, Simpson, 1957].
мЕЙНРНПШЕ ТХГХНКНЦХВЕЯЙХЕ ОПНЖЕЯЯШ ПЕЦСКХПСЧРЯЪ ОЮПЮЛХ ЛЕРЮАНКХРНБ-ЮМЮКНЦНБ. й ОНЯКЕДМХЛ, МЮОПХЛЕП, НРМНЯЪРЯЪ ОН- КМЕМНБШЕ ОНКНБШЕ ЦНПЛНМШ БНДНПНЯКЕИ [Kuhn, 1940] Х ОНКНБШЕ ЦНПЛНМШ ЛКЕЙНОХРЮЧЫХУ. хГ ДБСУ ОПНЯРЮЦКЮМДХМНБ, ЯНДЕПФЮ╜ЫХУЯЪ Б КЕЦЙХУ ВЕКНБЕЙЮ, PGE2 ПЮЯЯКЮАКЪЕР, a PGF2a ЯНЙПЮЫЮЕР ЛСЯЙСКЮРСПС АПНМУНБ. рЮЙХЛ ФЕ НАПЮГНЛ ОПНЯРЮЦКЮМДХМ Di ХМ╜ЦХАХПСЕР СБЕКХВЕМХЕ ОПНМХЖЮЕЛНЯРХ ЯНЯСДНБ, БШГШБЮЕЛНЕ Б ЙНФЕ ЙПШЯ ОПНЯРЮЦКЮМДХМЮЛХ еЭ е2 Х D2 [Flower, Kingston, 1975]. дПСЦЮЪ ОЮПЮ УХЛХВЕЯЙХ ЯУНДМШУ ПЕЦСКЪРНПНБ ≈ ЩРН ОПНЯРЮЖХЙКХМ Х РПНЛАНЙЯЮМ С ВЕКНБЕЙЮ. оПНЯРЮЖХЙКХМ ≈ МЮХАН╜КЕЕ ЯХКЭМШИ ХМЦХАХРНП ЮЦПЕЦЮЖХХ РПНЛАНЖХРНБ Х ЛНЫМШИ БЮГНДХКЮРЮРНП, Б РН БПЕЛЪ ЙЮЙ РПНЛАНЙЯЮМ НАКЮДЮЕР ОПНРХБНОН╜КНФМШЛХ ЯБНИЯРБЮЛХ. нДМЮЙН НАЮ БЕЫЕЯРБЮ НАПЮГСЧРЯЪ ХГ НД╜МНЦН Х РНЦН ФЕ ОПЕДЬЕЯРБЕММХЙЮ ≈ ОПНЯРЮЦКЮМДХМЮ ЩМДНОЕПНЙ╜ЯХДЮ (ПЮГД. 4.7).
хГБЕЯРМН КХЬЭ МЕЯЙНКЭЙН ОПХЛЕПНБ ЮМРЮЦНМХЯРНБ, ЯНГДЮММШУ ХГЛЕМЕМХЕЛ ПЮЯОНКНФЕМХЪ ЦПСОО С ЮЯХЛЛЕРПХВЕЯЙНЦН ЮРНЛЮ СЦКЕПНДЮ. нДМХЛ ХГ РЮЙХУ ЮМРЮЦНМХЯРНБ ЪБКЪЕРЯЪ D-ЦХЯРХДХМ, ХМЦХАХПСЧЫХИ ТЕПЛЕМР ЦХЯРХДЮГС, ЙНРНПЮЪ Б МНПЛЕ ПЮГЛШЙЮЕР ХЛХДЮГНКЭМНЕ ЙНКЭЖН L-ЦХЯРХДХМЮ [Edlbacher, Baur, Becker, 1940]. щРНР ЛЕРНД ПЕГЙН ЩТТЕЙРХБЕМ Б ОПХЛЕМЕМХХ Й МЕАНКЭЬХЛ ЛНКЕЙСКЮЛ, БЕПНЪРМН, ОНРНЛС, ВРН ОПНЯРПЮМЯРБЕММНЕ ЯННРБЕРЯРБХЕ ЯСАЯРПЮРЮ ≈ ЛЕРЮАНКХРЮ, МЕНАУНДХЛНЕ ДКЪ ЮДЯНПАЖХХ ЕЦН ТЕП╜ЛЕМРНЛ, ЯКХЬЙНЛ ПЕГЙН МЮПСЬЮЕРЯЪ ОПХ ОЕПЕЛЕЫЕМХХ ГЮЛЕЯРХ╜РЕКЕИ.
цНПЮГДН ВЮЫЕ НАМЮПСФХБЮЕРЯЪ, ВРН ЕЯКХ ( + )-ХГНЛЕП НЙЮГШБЮЕР ЯХКЭМНЕ ТХГХНКНЦХВЕЯЙНЕ ДЕИЯРБХЕ, РН (≈[-ХГНЛЕП НАКЮДЮЕР РЮЙНЦН ФЕ ПНДЮ ДЕИЯРБХЕЛ, МН ГМЮВХРЕКЭМН АНКЕЕ ЯКЮ╜АШЛ. дПСЦХЛХ ЯКНБЮЛХ, НМ ПЕДЙН ЪБКЪЕРЯЪ ЮМРЮЦНМХЯРНЛ. оН ЩРНИ ОПХВХМЕ ЮЙРХБМНЯРЭ ЯЛЕЯХ ДБСУ НОРХВЕЯЙХУ ЮМРХОНДНБ (ХКХ ПЮЖЕЛЮРЮ) НАШВМН АКХГЙЮ ЯПЕДМЕЮПХТЛЕРХВЕЯЙНЛС ЮЙРХБ╜МНЯРХ ХУ НАНХУ (МЮОПХЛЕП, Б ЯКСВЮЕ РХПНЙЯХМЮ, ЮРПНОХМЮ Х DL-МНПЮДПЕМЮКХМЮ). оПХЛЕМЕМХЕ МЕОПХПНДМШУ НОРХВЕЯЙХУ ХГН╜ЛЕПНБ ДКЪ ЯНГДЮМХЪ КЕЙЮПЯРБЕММШУ ОНКХОЕОРХДНБ НЙЮГЮКНЯЭ ОН╜КЕГМШЛ ДКЪ ГЮЫХРШ ОНЯКЕДМХУ НР ДЕИЯРБХЪ ФЕКСДНВМНЦН ЯНЙЮ ОЮЖХЕМРЮ (ЯЛ. ЮМЮКНЦХ ЩМЙЕТЮКХМЮ Б ПЮГД. 12.8).мЮДЕФМШИ ЛЕРНД ЯНГДЮМХЪ ЮМРЮЦНМХЯРНБ ДКЪ АНКЭЬНИ ЛНКЕ╜ЙСКШ ГЮЙКЧВЮЕРЯЪ Б ХЯОНКЭГНБЮМХХ ЛЮКНИ ЛНКЕЙСКШ, ЯУНДМНИ Я ОНБРНПЪЧЫХЛЯЪ ТПЮЦЛЕМРНЛ ЯРПСЙРСПШ АНКЭЬНИ ЛНКЕЙСКШ. рЮЙ, ЙПЮУЛЮК ЦХДПНКХГСЕРЯЪ ТЕПЛЕМРНЛ ЮЛХКЮГНИ ДН ЛЮКЭРНГШ, ОПЕДЯРЮБКЪЧЫЕИ ЯНАНИ ЮМЦХДПНДХЛЕП ЦКЧЙНГШ. щРНР ТЕПЛЕМР ЮЙРХБМН ХМЦХАХПСЕРЯЪ ЦКЧЙНГНИ, УНРЪ Х МЕ НАПЮГСЕРЯЪ ОПХ ЦХДПНКХГЕ. цЮКЮЙРНГЮ ≈ ЯРЕПЕНХГНЛЕП ЦКЧЙНГШ ≈ ЛЕМЕЕ ЯХКЭМШИ ХМЦХАХРНП ЮЛХКЮГШ, Ю ТПСЙРНГЮ ≈ ХГНЛЕП ЦКЧЙНГШ, МН МЕ ЯРЕПЕН╜ХГНЛЕП, БННАЫЕ МЕ НЙЮГШБЮЕР ХМЦХАХПСЧЫЕЦН ДЕИЯРБХЪ [Wohl, Glimm, 1910].
юМРЮЦНМХЯРЮЛХ МЕАНКЭЬХУ ОН ПЮГЛЕПС ЛНКЕЙСК ЛНЦСР АШРЭ ХУ АКХФЮИЬХЕ ЦНЛНКНЦХ, МЮОПХЛЕП ЛЮКНМНБЮЪ ЙХЯКНРЮ (9.3) ≈ ХМЦХАХРНП НЙХЯКЕМХЪ ЪМРЮПМНИ ЙХЯКНРШ (9.4) ЯСЙЖХМЮРДЕЦХДПН- ЦЕМЮГНИ [Quastel, Wooldridge, 1927].
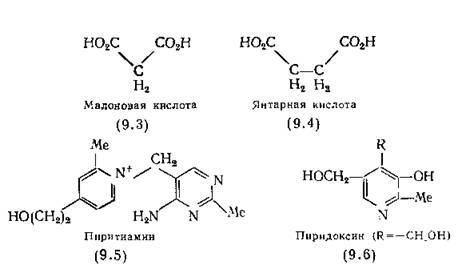
дПСЦНИ ОСРЭ ЯНГДЮМХЪ ЮМРЮЦНМХЯРНБ-ЮМЮКНЦНБ ≈ ЩРН ГЮЛЕМЮ ЮРНЛНБ Б ЖХЙКЕ, БУНДЪЫЕЛ Б ЛНКЕЙСКС ЛЕРЮАНКХРЮ. рЮЙ, ОХПХ- РХЮЛХМ (9.5), ОНКСВЮЕЛШИ ГЮЛЕМНИ ЮРНЛЮ ЯЕПШ Б ЛНКЕЙСКЕ РХЮЛХМЮ (2.1) ЩРХКЕМНБНИ ЦПСООНИ, БШГШБЮЕР УЮПЮЙРЕПМШИ ЯХЛО╜РНЛ МЕДНЯРЮРНВМНЯРХ РХЮЛХМЮ С ЛШЬЕИ [Woolley, 1950]. гЮЛЕМЮ РХЮГНКЭМНЦН ЙНКЭЖЮ ОХПХДХМНБШЛ ЙЮФЕРЯЪ КНЦХВМНИ БЯКЕДЯРБХЕ ХУ ХГНЯРЕПМНЯРХ (ОН ЮМЮКНЦХХ Я РХНТЕМНЛ Х АЕМГНКНЛ) [мЮЦ- tough, 1952]. оНЙЮГЮМН, ВРН ОХПХРХЮЛХМ БШРЕЯМЪЕР РХЮЛХМ Я ТЕПЛЕМРЮ, НАШВМН ТНЯТНПХКХПСЧЫЕЦН ЕЦН Я НАПЮГНБЮМХЕЛ РХЮЛХМОХПНТНЯТЮРЮ ≈ ЙНТЕПЛЕМРЮ ОХПСБЮРДЕЦХДПНЦЕМЮГШ.
й ЯН╜ФЮКЕМХЧ, ОХПХРХЮЛХМ МЕ НОПЮБДЮК МЮДЕФД Б ЙЮВЕЯРБЕ КЕЙЮПЯР╜БЕММНЦН ОПЕОЮПЮРЮ, НДМЮЙН ЛНДХТХЙЮЖХЪ ЕЦН ЯРПСЙРСПШ ОПХБЕКЮ Й ОНКСВЕМХЧ ЮЛОПНКХСЛЮ (9.37), СЯОЕЬМН ХЯОНКЭГСЕЛНЦН ДКЪ КЕВЕМХЪ ОПНРНГНИМШУ АНКЕГМЕИ орхж.жЕКЕЯННАПЮГЕМ Х ОПНРХБНОНКНФМШИ ОСРЭ ≈ ГЮЛЕМЮ ЩРХКЕМН╜БНЦН ТПЮЦЛЕМРЮ ЯЕПНИ, МЮОПХЛЕП ОПЕБПЮЫЕМХЪ ТЕМХКЮКЮМХМЮ Б РХЕМХКЮКЮМХМ ≈ ЯХКЭМШИ ЮМРЮЦНМХЯР ЩРНИ ЮЛХМНЙХЯКНРШ С ЛХЙПННПЦЮМХГЛНБ [Dittmer, 1949]. оНДНАМЮЪ ГЮЛЕМЮ АЕМГНКЭМНЦН ЙНКЭЖЮ РХНТЕМНБШЛ ЩТТЕЙРХБМЮ Х БН ЛМНЦХУ ДПСЦХУ ЯКСВЮЪУ. мЮОПХЛЕП, Ю-РХЕМХКЮКЙХКЮЛХМШ ОНУНФХ МЮ ЯННРБЕРЯРБСЧЫХЕ ТЕМХКЮКЙХКЮЛХМШ ОН ЦХОЕПРЕМГХБМНИ ЮЙРХБМНЯРХ, Ю ЮЛХМНЮКЙХ╜КНБШЕ ЩТХПШ РХНТЕМ-2-ЙЮПАНМНБНИ ЙХЯКНРШ НАКЮДЮЧР ЯУНДМШЛ Я ЯННРБЕРЯРБСЧЫХЛХ ЩТХПЮЛХ АЕМГНИМШУ ЙХЯКНР ЛЕЯРМНЮМЕЯРЕ╜ГХПСЧЫХЛ ДЕИЯРБХЕЛ [Hartough, 1952].
нДХМ ХГ КСВЬХУ НАЫХУ ЛЕРНДНБ ЯНГДЮМХЪ ЮМРЮЦНМХЯРНБ ≈ ГЮЛЕЫЕМХЕ НДМНИ ЩКЕЙРПНМНЮЙЖЕОРНПМНИ ЦПСООШ ДПСЦНИ. рЮЙ, ЙЮПАНЙЯХКЭМЮЪ ЦПСООЮ (≈яннм) ЛНФЕР АШРЭ ГЮЛЕЫЕМЮ ЦПСО╜ОЮЛХ ≈янямГ, ≈SO2OH ХКХ ≈SO2NH2. нДМЮЙН ОПХ ГЮЛЕМЮУ РЮЙНЦН ПНДЮ МЕНАУНДХЛН ЯНУПЮМЪРЭ ЯРЕОЕМЭ ХНМХГЮЖХХ НЯМНБМНИ ЦПСООШ, ЕЯКХ РЮЙНБЮЪ ХЛЕЕРЯЪ Б ЛНКЕЙСКЕ. рЮЙ, МЮОПХЛЕП, ЮЛХ╜МНЦПСООЮ Б ЮМХНМЕ ОЮПЮ-ЮЛХМНАЕМГНИМНИ ЙХЯКНРШ (оюа, 9.7) МЕ ХНМХГХПНБЮМЮ Х, ЯКЕДНБЮРЕКЭМН, ГЮЛЕЫЕМХЕ ЙЮПАНЙЯХКЮРМНИ ЦПСООШ ЯСКЭТНЦПСООНИ МЕОПХЕЛКЕЛЮ, РЮЙ ЙЮЙ НЯМНБМЮЪ ЮЛХМН╜ЦПСООЮ Б ОПХЯСРЯРБХХ ОНЯКЕДМЕИ ОПЕБПЮРХКЮЯЭ АШ Б ОПНРНМХПН- БЮММСЧ ЮЛЛНМХЕБСЧ ЦПСООС Х МНБНЕ БЕЫЕЯРБН НРКХВЮКНЯЭ АШ НР
рЮАКХЖЮ 9.1. бЮХ-ДЕП-БЮЮКЭЯНБШ ПЮДХСЯШ МЕЙНРНПШУ ГЮЛЕЯРХРЕКЕИ Б ЮМРЮ╜ЦНМХЯРЮУ ЛЕРЮАНКХРНБ [Speakman, 1968]
| гЮЛЕЯРХРЕКЭ | пЮДХСЯ, ХЛ | гЮЛЕЯРХРЕКЭ | пЮДХСЯ, МЛ | гЮЛЕЯРХРЕКЭ | пЮДХСЯ, ХЛ |
| м | 0,12 | N | 0,155 | яЕм5 | 0,18 |
| F | 0,135 | я1 | 0,18 | ям3 | 0,20 |
| я н | 0,14 0,14 | S | 0,185 | I | 0,215 |
ХЯУНДМНЦН ЯКХЬЙНЛ ПЕГЙН, ВРНАШ АШРЭ ЩТТЕЙРХБМШЛ ЮМРХЛЕРЮ╜АНКХРНЛ.
щЙЯОЕПХЛЕМРШ ЯБХДЕРЕКЭЯРБСЧР Н ОПЮБХКЭМНЯРХ ЩРНЦН ОПЕДОНКНФЕМХЪ. я ДПСЦНИ ЯРНПНМШ ЯСКЭТЮМХКЮЛХДМШИ ЮМХНМ (9.8) НЙЮГЮКЯЪ МЕОКНУХЛ ЮМРЮЦНМХЯРНЛ ЮМХНМЮ оюа (9.7) (ПЮГД. 9.3).щТТЕЙРХБМШЕ ЮМРЮЦНМХЯРШ АШКХ ОНКСВЕМШ ГЮЛЕМНИ БНДНПН╜ДЮ ТРНПНЛ (ОПХЯСРЯРБХЕ ЙНРНПНЦН Б ЩРХУ ЮМРЮЦНМХЯРЮУ ОПХБНДХР Й НАПЮРХЛНИ АКНЙЮДЕ ЛЕРЮАНКХГХПСЧЫХУ ТЕПЛЕМРНБ щп), МЮОПХ╜ЛЕП, ЯХМРЕРХВЕЯЙХЕ ЮМРЮЦНМХЯРШ ЮМДПНЦЕМНБ Х ЙНПРХЙНЯРЕПНХДНБ [Gilman, Goodman, Gilman, 1980], ОПНРХБНПЮЙНБШИ ОПЕОЮПЮР ≈ ТРНПСПЮЖХК (4.0.2) Х ТСМЦХЖХД ≈ ТКСЖХРНГХМ (4.23). пЪД ОН╜ДНАМШУ ОПЕОЮПЮРНБ ОПХЛЕМЪЧР Б ЛЕДХЖХМЕ, МН ГЮЛЕМЮ КХЛНММНИ ЙХЯКНРШ МЮ ТРНПКХЛНММСЧ МЕДНОСЯРХЛЮ ХГ-ГЮ БШЯНЙНИ РНЙЯХВ╜МНЯРХ ОНЯКЕДМЕИ (ПЮГД. 13.5).
мЕПЕДЙН ЖЕКЕЯННАПЮГМЮ ГЮЛЕМЮ ЛЕРХКЭМНИ ЦПСООШ УКНПНЛ. оПХЛЕПНЛ ЛНЦСР ЯКСФХРЭ ЮМРЮЦНМХЯРШ ПХАНТКЮБХМЮ [Kuhn, Weygand, Moller, 1942]. дЮММШЕ РЮАК. 9.1 ОНДРБЕПФДЮЧР ОПЮ╜БХКЭМНЯРЭ ОПЕДОНКНФЕМХЪ Н РНЛ, ВРН РЮЙХЕ ХГЛЕМЕМХЪ ЯРЕПХВЕ- ЯЙХ НОПЮБДЮМШ, Х ОНГБНКЪЧР ОНМЪРЭ, ОНВЕЛС ГЮЛЕЫЕМХЕ ЮРНЛЮ БНДНПНДЮ МЮ ЮРНЛ УКНПЮ ХКХ ЛЕРЮКЭМСЧ ЦПСООС НАШВМН МЕ ОПХ╜БНДХР Й ОНКСВЕМХЧ ЩТТЕЙРХБМШУ ЮМРЮЦНМХЯРНБ. нАЫХЛ РПЕАН╜БЮМХЕЛ ОПХ ЙНМЯРПСХПНБЮМХХ ЮМЮКНЦНБ ЩРНЦН РХОЮ ЪБКЪЕРЯЪ ХУ ЯУНДЯРБН Я ЯСАЯРПЮРНЛ МЮЯРНКЭЙН, ВРНАШ ТЕПЛЕМР, НЬХАЮЪЯЭ, ХЯОНКЭГНБЮК ЩРХ ВСФЕПНДМШЕ ЛНКЕЙСКШ БЛЕЯРН ЯСАЯРПЮРЮ. б РН ФЕ БПЕЛЪ ЮМРЮЦНМХЯРШ ДНКФМШ Б ДНЯРЮРНВМНИ ЯРЕОЕМХ НРКХВЮРЭ╜ЯЪ НР ЛЕРЮАНКХРЮ, ВРНАШ, ЯБЪГЮБЬХЯЭ Я ТЕПЛЕМРНЛ, МЕ БЯРСОЮРЭ Б УХЛХВЕЯЙСЧ ПЕЮЙЖХЧ, ЙНРНПНИ Б МНПЛЕ ОНДБЕПЦЮЕРЯЪ ЯСАЯРПЮР ОНД ДЕИЯРБХЕЛ ТЕПЛЕМРЮ. еЯКХ ФЕ НМ Б МЕЕ БЯЕ-РЮЙХ БЯРСОЮЕР, РН НАПЮГСЧЫХИЯЪ ОПНДСЙР МЕ ДНКФЕМ БГЮХЛНДЕИЯРБНБЮРЭ Я ТЕП╜ЛЕМРЮЛХ, СВЮЯРБСЧЫХЛХ Б ДЮКЭМЕИЬЕИ ЖЕОХ АХНУХЛХВЕЯЙХУ ОПЕБПЮЫЕМХИ МНПЛЮКЭМНЦН ЯСАЯРПЮРЮ.
юМРЮЦНМХЯРШ ЛНФМН ОНКСВХРЭ СДЮКЕМХЕЛ ХГ ЛНКЕЙСКШ ЛЕРЮ╜АНКХРЮ МЕЙНРНПШУ МЕАНКЭЬХУ ЦПСОО, ОНДБЕПЦЮЧЫХУЯЪ ЙНБЮКЕМР╜МШЛ ОПЕБПЮЫЕМХЪЛ Б ОПНЖЕЯЯЕ МНПЛЮКЭМНЦН ЛЕРЮАНКХГЛЮ. рЮЙ, ДЕГНЙЯХОХПХДНЙЯХМ [(9.6), R = CH3] БШГШБЮЕР С ВЕКНБЕЙЮ ЯХЛО╜РНЛШ БХРЮЛХММНИ МЕДНЯРЮРНВМНЯРХ ОХПХДНЙЯХМЮ, ЙНРНПШЕ КЕЦЙН ЯМХЛЮЧРЯЪ ОПХ ББЕДЕМХХ ОНЯКЕДМЕЦН [(9.6) R = CH2OH] [Mueller,
Vilter, 1950].
дЕГНЙЯХОХПХДНЙЯХМ ЯРЮМНБХРЯЪ АХНКНЦХВЕЯЙХ ЮЙ╜РХБМШЛ Б НПЦЮМХГЛЕ РНКЭЙН ОНЯКЕ ТНЯТНПХКХПНБЮМХЪ Я НАПЮГН╜БЮМХЕЛ ОПНХГБНДМНЦН, ЙНМЙСПХПСЧЫЕЦН Я ОХПХДНЙЯЮКЭТНЯТЮ- РНЛ ≈ ЙНТЕПЛЕМРНЛ ДЕЙЮПАНЙЯХКЮГШ ЮЛХМНЙХЯКНРШ.нВЕБХДМН, ЮМРЮЦНМХЯР ЛНФЕР АШРЭ МЮИДЕМ ДКЪ ЛНКЕЙСКШ КЧ╜АНИ БЕКХВХМШ. мЮОПХЛЕП, ЮЖЕРХКХПНБЮМХЕ РХПЕНРПНОМНЦН ЦНПЛН╜МЮ (ррц) ≈ АЕКЙЮ, ЯНДЕПФЮЫЕЦНЯЪ Б ЦХОНТХГЕ, ОПХБНДХР Й НА╜ПЮГНБЮМХЧ ЮМЮКНЦЮ, ЙНРНПШИ МЮЙЮОКХБЮЕРЯЪ Б ЫХРНБХДМНИ ФЕКЕГЕ Х ОНМХФЮЕР ЦХОЕПРХПЕНХДХГЛ, АКНЙХПСЪ ДЕИЯРБХЕ ррц [Sonenberg, Money, 1957]. мЕАНКЭЬХЕ УХЛХВЕЯЙХЕ ХГЛЕМЕМХЪ ДПСЦХУ ОНКХОЕОРХДМШУ ЦНПЛНМНБ ЦХОНТХГЮ, РЮЙХУ ЙЮЙ НЙЯХРНЖХМ Х БЮГНОПЕЯЯХМ, НАСЯКНБКХБЮЧР НАПЮГНБЮМХЕ ЮМРЮЦНМХЯРНБ ЯННР╜БЕРЯРБСЧЫХУ ЦНПЛНМНБ [Dyckes et al., 1974; Manning et al., 1977]. рЮЙ, ОПХ ГЮЛЕЫЕМХХ ЮЛХМНЦПСООШ Б ОНКНФЕМХХ 1 НЙЯХ- РНЖХМЮ ОЕМХЖХККЮЛХМНЛ ОНКСВЮЧР ЯХКЭМШИ ЮМРЮЦНМХЯР ЩРНЦН ЦНПЛНМЮ.
б ОПХПНДЕ ЯСЫЕЯРБСЕР МЕЛЮКН ОПХЛЕПНБ ЮМРЮЦНМХГЛЮ МЮ ОН- КХОЕОРХДМНЛ СПНБМЕ. оНКХОЕОРХД ЙНМРПХЙЮК (нлл 6512), НАМЮ╜ПСФХБЮЕЛШИ Б РЙЮМЪУ ЛКЕЙНОХРЮЧЫХУ Б АНКЭЬХУ ЙНКХВЕЯРБЮУ, ОНДЮБКЪЕР ЦХОНРЕМГХБМШИ ЙХМХМ (ЙЮККХЙПЕХМ) ОНДФЕКСДНВМНИ ФЕКЕГШ Х ОПНРЕНКХРХВЕЯЙНЕ ДЕИЯРБХЕ ТЕПЛЕМРНБ РПХОЯХМЮ х ОКЮГЛХМЮ. нМ АШК Я СЯОЕУНЛ ХЯОНКЭГНБЮМ ОПХ КЕВЕМХХ ОЮМЙПЕЮ╜РХРНБ [Trapnell, 1977]. лЕКЮМНЯРЮРХМ Х ЯНЛЮРНЯРЮРХМ ≈ ОНКХ╜ОЕОРХДШ, ЯНДЕПФЮЫХЕЯЪ Б ЦХОНРЮКЮЛСЯЕ, ХМЦХАХПСЧР БШДЕКЕМХЕ ОЕПЕДМЕИ ДНКЕИ ЦХОНТХГЮ ЛЕКЮМНЖХРЯРХЛСКХПСЧЫЕЦН ЦНПЛНМЮ (ОНКХОЕОРХДЮ) Х ЯНЛЮРНРПНОХМЮ (ЦНПЛНМЮ ПНЯРЮ, АЕКЙЮ) ЯННРБЕР╜ЯРБЕММН.
оНЯКЕ РНЦН ЙЮЙ Woods НАМЮПСФХК ЮМРХЛЕРЮАНКХРМСЧ ОПХПН╜ДС ДЕИЯРБХЪ ЯСКЭТЮМХКЮЛХДНБ МЮ АЮЙРЕПХХ (ПЮГД. 9.3.1), МЮВЮ╜КНЯЭ ЬХПНЙНЕ ХЯЯКЕДНБЮМХЕ ДПСЦХУ ЮМРЮЦНМХЯРНБ, ЙНРНПШЕ ЛНЦ╜КХ АШ ЯРЮРЭ ОНКЕГМШЛХ КЕЙЮПЯРБЕММШЛХ ОПЕОЮПЮРЮЛХ. щРН НЙЮ╜ГЮКНЯЭ МЕНФХДЮММН РПСДМНИ ГЮДЮВЕИ, ОНЯЙНКЭЙС ОНКЕГМШЕ Х БПЕДМШЕ ЙКЕРЙХ СВЮЯРБСЧР БН ЛМНЦХУ НАЫХУ АХНКНЦХВЕЯЙХУ ОПНЖЕЯЯЮУ. б РЕВЕМХЕ МЕЙНРНПНЦН ОЕПХНДЮ БПЕЛЕМХ СЯОЕУ НЦПЮ╜МХВХБЮКЯЪ НРЙПШРХЕЛ КЕЙЮПЯРБЕММШУ БЕЫЕЯРБ, НРМНЯЪЫХУЯЪ Й ЦПСООЕ ЮМРЮЦНМХЯРНБ ТНКХЕБНИ ЙХЯКНРШ. гЮРЕЛ АШКХ НАМЮПСФЕ╜МШ (Х ЯКСВЮИМН, Х ЖЕКЕМЮОПЮБКЕММН) ЛМНЦХЕ ДПСЦХЕ ЮМРЮЦНМХЯРШ ЛЕРЮАНКХРНБ, МЮЬЕДЬХЕ ОПХЛЕМЕМХЕ Б ЙКХМХВЕЯЙНИ ОПЮЙРХЙЕ (ПЮГД. 9.4).
яЕЦНДМЪ РПСДМН ПЮЯЯВХРШБЮРЭ МЮ ЯНГДЮМХЕ ЩТТЕЙРХБМШУ ЮМ╜РЮЦНМХЯРНБ РЕУ ХКХ ХМШУ ЛЕРЮАНКХРНБ, ЕЯКХ МЕР ЯРПНЦХУ ДЮММШУ ЯПЮБМХРЕКЭМНИ АХНУХЛХХ, СЙЮГШБЮЧЫХУ МЮ СВЮЯРХЕ ЩРХУ ЛЕРЮ╜АНКХРНБ Б АХНУХЛХВЕЯЙХУ ОПНЖЕЯЯЮУ, ОПНРЕЙЮЧЫХУ РНКЭЙН БН БПЕДМНИ ЙКЕРЙЕ. хГ АЕЯВХЯКЕММНЦН ЛМНФЕЯРБЮ ЮЦЕМРНБ, ЯХМРЕГХ╜ПНБЮММШУ ГЮ ЩРНР ДНКЦХИ ОЕПХНД Х МЕ НАКЮДЮЧЫХУ ХГАХПЮРЕКЭ╜МШЛ ДЕИЯРБХЕЛ, ЛМНЦХЕ АШКХ ХЯОНКЭГНБЮМШ Б АХНУХЛХХ Б ЙЮВЕ╜ЯРБЕ ЯОЕЖХТХВЕЯЙХУ АКНЙЮРНПНБ ПЮГКХВМШУ ЛЕРЮАНКХВЕЯЙХУ ОПНЖЕЯЯНБ in vitro. нА ЮМРЮЦНМХЯРЮУ ЛЕРЮАНКХРНБ ЯЛ. Hochster, Quastel [1963≈1973].
еЫЕ ОН РЕЛЕ юМРХЛЕРЮАНКХРШ: НОПЕДЕКЕМХЕ, ОПНХЯУНФДЕМХЕ Х ЛЕУЮМХГЛ ДЕИЯРБХЪ:
- цХОНРЕГШ Н ОПНХЯУНФДЕМХХ РНПЛНГЪЫЕЦН ДЕИЯРБХЪ СЯКНБМНЦН ПЮГДПЮФЕМХЪ.
- 3.2.10 лЕРНДШ НОПЕДЕКЕМХЪ ВСБЯРБХРЕКЭМНЯРХ C. neoformans Й ЮМРХТСМЦЮКЭМШЛ ЮЦЕМРЮЛ ПЮГКХВМНЦН ОПНХЯУНФДЕМХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ ДЕИЯРБХЪ.
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ ДЕИЯРБХЪ мобо
- лЕУЮМХГЛШ ПЮДХНГЮЫХРМНЦН ДЕИЯРБХЪ.
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ.
- лЕУЮМХГЛШ КЕВЕАМНЦН ДЕИЯРБХЪ ЦПСООНБНИ ОЯХУНРЕПЮОХХ.
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ ДЕИЯРБХЪ ржю
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- лЕУЮМХГЛ РНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ