ГЛАВА XlV ОСНОВНЫЕ ПОЛОЖЕНИЯ УЧЕНИЯ О ПРИРОДНОЙ ОЧАГОВОСТИ БОЛЕЗНЕЙ
Е. Н. Павловский
В январе 1939 г. Е. Н. Павловским на Общем собрании Академии наук СССР был сделан доклад по проблеме эпидемиологической концепции о природе как об источнике заражения человека возбудителями различного характера, основанный на развернутых примерах четырех природноочаговых болезней разного характера (Й.
Г. Галузо, 1961).Учение о природной очаговости болезней имело ряд предпосылок, касавшихся разрозненных фактов, которые, взятые в отдельности, не давали связанной общей концепции, но хорошо укладывались в ее рамки, служащие надежным ориентиром при исследованиях некоторых болезней.
Предпосылками учения о природной очаговости болезней являлись следующие обстоятельства. В прошлом столетии, когда русские войска впервые вступали на территорию реки Мургаба в Туркмении, целые полки почти поголовно заболевали пендинской язвой (кожным лейшманиозом) при прохождении по территории пустыни и полупустыни. Тщетны были попытки ряда врачей выяснить причины массового развития этой «восточной язвы». Значительным шагом вперед было открытие П. Ф. Боровским в 1898 г. возбудителя этой болезни. Но долгое время оставалось загадкой, почему в зоне полупустыни и пустыни вступающие туда в определенное время люди заболевают кожным лейшманиозом, которым люди заражались не друг от друга, а от какого-то общего источника. Разгадка была достигнута позднее. Уже в советское время работами паразитологических экспедиций и лейшманиозной лаборатории Всесоюзного института экспериментальной медицины Н. И. Латышевым и А. Крюковой была выяснена роль грызунов песчанок и сусликов как источников заражения москитов (Phlebotonius) возбудителем кожного лейшманиоза и москитов как его переносчиков другим животным. По совокупности всех данных автором и была обоснована природ ная очаговость болезни Боровского для людей; по данным зарубежной литературы, можно с уверенностью утверждать, что и кала-азар (внутренностный лейшманиоз) также является в своей основе природноочагопой болезнью.
Другой предпосылкой было установление Д. К. Заболотным чумы у грызунов (тарбаганов) и выяснение рядом ученых роли блох в передаче чумных микробов, что ставило чуму в группу трансмиссивных болезней.
Советское время ознаменовалось весьма широким развитием экспеди цпонных исследований, в частности различного медицинского характера
Параллельно шло развитие сети специальных противочумных, протнво туляремийных учреждений и институтов разного медицинского профиля (институты эпидемиологии и гигиены, противомалярийные учреждения.
институты вакцин и сывороток и др.), которые в той или другой мере непосредственно или косвенно касались и проблем природной очаговости болезней. Все это приводило к мощному росту специальной литературы, укреплешпо значения этого учения, которое принимало не только общегосударственный масштаб, но и получило международное признание.
Показателями расширения значения этого учения являлись: преобразование Омского государственного научно-исследовательского института эпидемиологии, микробиологии JT гигиены в Институт по природноочаговым болезням с филиалом в Тюмени, открытие аналогичных отделов в ряде Институтов эпидемиологии и микробиологии, преобразование ранее бывшего отдела паразитологии и медицинской зоологии в отдел по природноочаговым инфекциям Института эпидемиологии и микробиологии имени Н. Ф. Гамалеи; активное участие кафедр некоторых медицинских институтов (биологии и инфекционных болезней в Кишиневе — Молдавская ССР) в разработке соответственных вопросов в пределах республики и др.
За рубежом Всемирная организация здравоохранения дважды обращалась к Советскому правительству с просьбой о проведении специальных курсов по природной очаговости болезней для медиков разных государств Европы, Азии, Африки, Америки в I960 и в 1962 гг. (Е. Н. Павловский, 1962). Был приведен также симпозиум советских ученых и ученых многих стран социалистического лагеря. За рубежом это учение энергично разрабатывают в Чехословакии, Польше, Болгарии, ГДР и в других государствах (Павлов, 1960; Е.
Н. Павловский, 1963).К настоящему времени природная очаговость установлена для следующих болезней.
Вирусные болезни: клещевой и японский (комариный) энцефалиты (Л. А. Зильбер, 1945; П. А. Петрищева, 1947; А. А. Смородинцев с сотр., 1941; Н. И. Гращенков, 1947; И. Б. Галант, 1955); лимфоцитарный хориоме- нингнт; желтая лихорадка; бешенство; орнитозы (пситтакоз и др.); весьма вероятно — лихорадка паппатачи; спирохетозы: клещевой возвратный тиф, лептоспирозы (В. В. Ананьин и В. В. Карасева, 1961); риккетсиозы (клещевой сыпной тиф, лихорадка Ку, японская речная лихорадка, кустарниковый тиф); особо опасные бактериальные инфекции (чума, туляремия) и другие бактериальные инфекции (сибирская язва, листереллез, эризипелоид, рожа свиней); геморрагические лихорадки; пефрозо-нефрит; некоторые микозы; протозойные болезни: * лейшманиозы (болезнь Боровского, кала-азар), трипаносомозы [сонная болезнь Африки, болезнь Чагаса в Южной Америке (С. А. Гоар, 1960, 1961), токсоплазмоз]; гельминтозы [описторхоз, дифплло- ботриоз, трихинеллез, бильгарциозы и многие другие (И. И. Галузо, 1961)]; акарозы (частично ■— чесотка); энтомозы (миазы от личинок вольфартовой мухи, larva migraπs от оводов) и другие болезни.
Как видно из этого неполного перечня природноочаговых болезней, последние имеют самых разнообразных возбудителей от вирусов до насекомых. Также разнообразны источники и пути заражения ими людей. Однако при таком этиологическом и эпидемиологическом разнообразии эти нозологические формы имеют нечто общее, позволяющее включать их в одну группу природноочаговых болезней, число которых увеличивается по мере применения общего биолого-медицинского критерия для их оценки.
Этим критерием является то, что источником заражения такими болезнями является природа, нетронутая деятельностью человека, или видоизмененная человеком. Возбудители некоторых болезней этой категории легко переходят из природы в непосредственную близость к человеку — в селения, в города, даже в отдельные комнаты жилья, на выпасные
луга, в искусственные водоемы (колодцы, пруды, случайные скопления воды) и в водоемы природные (ручьи, реки).
Природноочаговыми являются болезни о б л и г а т н о - тр а н с м неси в н ы е (Е. Н. Павловский, 1940), возбудители которых на современном этапе эволюции передаются исключительно специфическими для них переносчиками (чаще кровососущими клещами, и так же питающимися насекомыми, например для сезонных энцефалитов — клещи и комары), и ф а- к у л ь т а т и в н о - т р а н с м и с с и в н ы е болезни, возбудителями которых люди заражаются разными путями из внешней среды (вода, наземный инфицированный субстрат, непосредственное соприкосновение промыслового характера с добытыми животными, оказавшимися больными) и п р и посредничестве неспецифических переносчиков разного систематического положения (например, туляремия).
Если бывают бесспорные случаи заболеваний людей, впервые вступающих на территорию зоны нетронутой ранее природы, т. е. в безлюдную ранее местность (тайга, степь, полупустыня, пустыня и др.), заболеваний, необычных, возбудителями которых люди могут заражаться друг от друга и заболевать везде, то мысль обращается в другую сторону, хотя бы сначала для гипотетического истолкования причин необычных заболеваний. Создается рабочая гипотеза, которая должна послужить канвой для последующих поисковых работ в полевых условиях и для экспериментальных лабораторных исследований.
Такая рабочая гипотеза может строиться по следующей схеме: если в безлюдной ранее местности исключается возможность заражения человека от человека, то источник заболеваний следует искать среди животных фауны соответственной местности. Известны болезни животных, которыми человек также может заболеть, заражаясь непосредственно от больного животного или же при посредничестве живых агентов, которыми служат различные кровососущие клещи и насекомые, выполняющие роль переносчиков. В отсутствие людей они нападают с целью питания на животных-прокормителей, среди которых могут быть особи с циркуляцией в периферической крови возбудителя зоонозной болезни или микроба, вызывающего состояние носительства; в таком случае переносчик заражается им и при последующем акте питания на здоровом прокормителе такой «спонтанно зараженный переносчик передает имеющегося в нем возбудителя своему новому прокормителю и заражает его.
Зараженный прокормитель в свою очередь приходит в состояние, заражающее свежего переносчика, и т. д. в течение неограниченного времени (рис. 43).Все это происходит на фоне определенного географического ландшафта в его биотопах, служащих местом жизни или временного пребывания животных, между которыми непрерывно происходит передача возбудителя болезни от одной особи другой в процессе последовательного питания на них переносчиков возбудителя. Всем этим признакам и соответствует природный очаг болезни. Если неиммунный восприимчивый человек вступает на территорию такого очага в период его валентного (действующего) состояния, т. е. когда в очаге имеются голодные спонтанно инфицированные переносчики, то они незамедлительно нападают на него для питания его кровью и в процессе ее всасывания передают человеку возбудителя болезни, полученного от дикого больного животного или от бактерионосителя. Человек — прокормитель голодного переносчика, заражается от него возбудителем болезни и далее явно заболевает ею или при некоторых болезнях в нем протекает ее инаппарантная, или скрытая (латентная), форма инфекции, приводящая организм в состояние невосприимчивости
к новому заражению активной дозой вирулентного возбудителя того же вида (векторная вакцинация).
Описанную схему соотношений следует дать, пользуясь научными био логическими и медицинскими терминами.
Природным очагом болезни является участок определенного географического или в известной степени окультуренного ландшафта, в биотопах которого существует биоценоз, т. е. сочетание минимума организмов, нахо дящихся в межвидовых биоценотических связях, преимущественно характер- ра цепей питания; осуществление таких алиментарных связей обеспечивает непрерывность перехода возбудителя болезни (патоэргонт) от больного
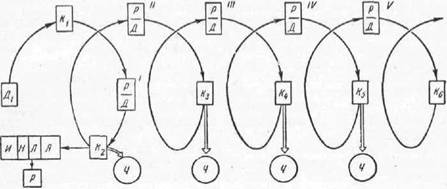
ее природном очаге.
Рис. 43. Схема циркуляции возбудителя (вируса) болезни р
Ді —исходный донор вируса; K1—переносчик вируса; — реципиент вируса, становящийся его донором; Kt — свежий переносчик вируса — клещ из популяции биоценоза; Ч —передача (вируса человеку); дальнейшего хода вирус от человека не получает (вирус попал в тупик); грансовариальная (Я — яйца.
Л — личинки) передача иируса самкой пирофорного клеща сс дочернему поколению и дальнейшая трансфазовая передача вируса (IJ — нимфа. И — имаго); P — реципиент; Кг—Ke — клещи-переносчики из состава популяции обеспечивающие передачу вируса свежим реципиентам биоценоза: J-V—развертывающиеся по спирали обороты (туры) в процессе циркуляции вируса в природном очаге болезни, обеспечивающие его суще ствование. Оригинал.животного или от носителя (животное донор возбудителя) через посредничество преимущественно кровопитающегося переносчика (вектора патоэргонта) к восприимчивому организму того же, что и донор, или другого вида (животное — восприемник или реципиент патоэргонта); реципиент через некоторое время приходит в состояние потенциального донора для новой партии переносчиков, которые передают возбудителя новым особям популяции реципиентов и т. д. Реципиентом может становиться и неимунный человек, вступающий на территорию природного очага болезни, когда в очаге в связи с сезо ном года имеются голодные переносчики. Нападая на человека как на богатый источник питания, переносчики передают ему возбудителя болезни, полученного ими от дикого донора; человек неминуемо заражается возбуди телем, что приводит его к явному заболеванию с разным исходом или у него развивается инаппарантная форма (скрытая, латентная форма) болезни с выработкой иммунитета к реинфекции достаточной дозой вирулентного возбудителя. В последнем случае естественно (спонтанно) зараженный век тор приобретает значение как бы живой вакцины.
Все эти процессы в природном очаге совершаются, если метеорологи ческие и другие факторы внешней среды не препятствуют такой передаче /циркуляции) возбудителя.
Природный очаг болезни существует все то время, пока осуществляется непрерывная циркуляция возбудителя болезни между животными донорами н реципиентами. Это время может исчисляться даже геологическими масштабами и эволюционное возникновение в природе очагов ряда болезней весьма вероятно приходилось на время, когда человек еще не появился на земле.
Происходя эволюционно от животных предков, люди могли получать от них в наследие и некоторые свойственные им болезни; процесс приручения животных приводил к тому, что донорами и реципиентами возбудителей болезни становились уже близкие к человеку домашние и сельскохозяйственные животные.
Видоизменения, которым подвергается природа под влиянием «хозяйственной» деятельности людей, могут приводить к противоположным результатам: а) при окультуривании местности человек настолько изменяет тот или другой географический ландшафт, что свойственные ему природные очаги болезней ослабляются или совсем исчезают вследствие разрушения структуры их биоценоза (деструкция) или же б) они все более и более приближаются к человеку, проникая даже в его жилье примитивной постройки; в) природные по своему существу очаги болезней могут даже возникать под влиянием бессознательной деятельности людей на пастбищах сельскохозяйственных животных (например, очаги бруцеллеза) или в местах их временного средоточия (например, очаги лептоспироза при загрязнении искусственно сооруженных прудов, которыми пользуются для водопоя скота, загрязняющего их лептоспирами со своими выделениями, попадающими в воду; то же при лагерном содержании свиней на берегах рек, вода которых инфицируется от загрязненных берегов и при купании людей в таких реках они заражаются лептоспирами).
Природные очаги болезней могут быть геоморфологически вполне опре д-е ленно ограниченными или п ед О ЛИ- МИТИ рован н ы м и; такова, например, нора грызуна или другого животного; но физические размеры их могут колебаться в чрезвычайных пределах — от поры сухопутной черепахи глубиной 30—40 см до норы дикобраза длиной более 10 м.
Норы животных бывают одиночные и сложные, как, например, городки песчанок; ходы их нор соединяются друг с другом и имеют много выходных отверстий.
Противоположным типом природных очагов являются очаги диффузного характера, не имеющие сколько-нибудь резко ограниченных границ, например лесная подстилка кедрово-широколиственных лесов или лесов листопадных. Правда, под слоем опавших листьев и хвои в почве могут быть норы мелких грызунов; но и самая толща лесной подстилки является местом нахождения иксодовых клещей, которые, насытившись и отпадая от своих прокормителей, находят в ней благоприятные места для пребывания на все время, необходимое для подготовки их к линянию на следующую фазу превращения. Но в лесной подстилке клещи распространены не равномерно, а мозаично. Так, переносчик вируса клещевого энцефалита Ixodes persulcatus в тайге скопляется по тропам, протоптанным животными к водопою; в стороне же от тропы клещей совсем может не быть; так же и вдоль тропы клещи распределяются по количеству далеко неравномерно.
Ixodes persulcatus обладает тонким обонянием, благодаря которому он занимает позицию в отношении места, где он может скорее прицепиться к проходящему мимо человеку или животному.
19 Заказ № 228
Природные очаги болезней имеют определенную общую структуру, определяемую сочетанием пентады (пятерки) факторов: 1) возбудитель болезни как таковой, 2) животное донор возбудителя, 3) для болезней трансмиссивных — переносчик возбудителя, 4) животное реципиент возбудителя и 5) факторы внешней среды, не препятствующие непрерывной передаче возбудителя через его переносчиков. Первые четыре компонента в своей совокупности образуют биоценоз определенного видового состава; но биоценоз природного очага болезни отличается от биоценоза в обычном зоологическом его понимании тем, что в состав его членов входит и возбудитель болезни, какой бы природы он ни был; в таком полном составе имеется совокупность организмов, которую следует обозначать особым термином биопатоценоз. Часть этого термина, обозначаемая слогами «пато», подчеркивает ту существенную особенность, что в этом термине учитывается абсолютная необходимость наличия возбудителя болезни даже и в тех случаях, когда он совсем не бывает во внешней среде и все время пребывает последовательно то в одном, то в другом организме. Такой биопатоценоз является замкнутым.
Классическим примером узко замкнутой циркуляции возбудителя болезни является трансмиссивная — ве природноочаговая протозойная болезнь — малярия. Переход ее возбудителя идет по короткой цепи:
донор — человек гаметоноситель, переносчик — самка комара рода Anopheles, реципиент — человек, получив через переносчика возбудителя малярии, заболевает малярией и со временем становится донором возбудителя для свежих (незараженных) комаров Anopheles и т. д.
Почти аналогичное видим и в соотношениях передачи вируса клещевого энцефалита его специфическими переносчиками, которыми являются иксо- довые клещи — Ixodes persulcatus, Dermacentor silvarum и Haemaphysalis conciπna, как это было показано экспедициями по сезонным энцефалитам 1938—1939 гг.
В дальнейшем число видов клещей-переносчиков возросло. Подтверждена роль упомянутых трех видов и прибавлен еще вид Haemaphysalis japonι- са douglasi) (Беликова и Татаринова, I960).
В отношении возможности выхода вируса клещевого энцефалита из организма больного вирусологи показали возможность обнаружения его в> носовой слизи; но данных о возможности заражения здоровых непосредственно от больных нет; наоборот, подчеркивают, что ни врачи, ни сестры, ни члены семьи не заражаются от больных; если же случится, что заболевают несколько членов семьи, то потому, что они заражались из какого-то общего источника.
В литературных данных чаще и чаще отмечают случаи энцефалита вследствие употребления сырого козьего молока. Такой «молочный энцефалит», по утверждению ряда вирусологов (А. А. Смородинцев, Ильенко и др.), является самостоятельной болезнью, хотя и общей группы с клещевым энцефалитом. Эпидемиологические различия их несомненны. Сравнительные эпидемиологические данные приводит В. Ф. Попов (1963) по наблюдениям в южных районах Кировской области. За четыре года (1959—1962) алиментарный путь передачи вируса клещевого энцефалита (это определение в дальнейшем изложении заменим словами «молочный энцефалит». — Е. Н. Павловский) был ведущим.
Трансмиссивный путь заражения наблюдался чаще среди населения деревень вследствие посещения ими леса по бытовым потребностям; контакт их с лесом был в 3 раза выше, чем в поселках рабочих и в поселках со смешанным населением.
Семейно-групповые заболевания молочным энцефалитом вследствие употребления сырого козьего молока наблюдались в рабочих и в смешанных поселках, где держали преимущественно коз. Высокий процент молочного энцефалита наблюдался у детей до 15-летнего возраста —• главных потребителей козьего молока.
По общему расчету иа 10 000 владельцев коз приходилось 377 заболеваний, но по районам число заболевших владельцев коз молочным энцефалитом колебалось: в одном районе заболевал один из 50 хозяев, в другом один из 26. Заболеваемость отмечалась главным образом среди местного населения.
Заболеваемость молочным энцефалитом начиналась и оканчивалась раньше, чем клещевым энцефалитом; это объяснялось выпасанием коз обычно на вырубках, где нападающая активность иксодовых клещей, по наблюдениям Земской и Суворовой, проявлялась примерно на 10 дней раньше, чем в высокоствольном лесу.
При молочном энцефалите (как и при клещевом) в ряде случаев развивались парезы и параличи; были даже смертные случаи. Соотношение клинически выраженных и инаппараптных форм колеблется с преобладанием последних. В 1960 г. одна клиническая форма приходилась на 33 инаппарантных, а в 1961 г. соотношения изменились до 1 на 77, а в 1962 — 1 па 66.
Хотя пути заражения людей клещевым и молочным энцефалитом существенно различны, все же и молочный энцефалит по своему существу является также клещевым, так как козы, в молоке которых обнаруживали вирус, могли получить его сами только от напавших на них вирофорных клещей; в процессе циркуляции в организме козы вирус из крови ее проникает в молочную железу и при доении C молоком выходит наружу. При употреблении некипяченого козьего молока вирус беспрепятственно попадает в желудочно-кишечный тракт.
Но есть и такие природноочаговые болезни, возбудители которых, как правило, выделяются из организма доноров во внешнюю среду, где разное и не редко продолжительное время сохраняют свои инфицирующие свойства; из внешней среды вирулентные возбудители разными путями проникают в организм человека и его заражают не трансмиссивно; но при всем разнообразии таких путей заражения параллельно бывает и трансмиссивный путь передачи возбудителя болезни его переносчиками человеку. Такой болезнью является туляремия.
Будучи открытой в США, она вскоре была обнаружена в СССР, где привлекла к себе большое внимание, что организационно выразилось в появлении специальных лабораторий и отделов в институтах медицинского профиля. Успешное развитие исследовательской и научно:практической работы советских и зарубежных ученых дредста- злено в книге «Туляремия» девяти авторов под ред. Н. Г. Олсуфьева и Г. П. Руднева.
Туляремией болеют многие животные разного систематического положения из классов птиц и млекопитающих, в частности и млекопитающие, имеющие промысловое значение. Большое значение в распространении туляремии имеет водяная крыса, в настоящее время называемая водяной полевкой. Она весьма чувствительна к заражению туляремийным микробом и в условиях эксперимента может быть заражена всего одной бактерией. Переносчиками возбудителя туляремии служат собственные эктопаразиты водяной полевки (вши, клещи, блохи) и летающие кровососущие двукрылые — комары, слепни, а также иксодовые клещи. Через них может заражаться и человек: трансмиссивные вспышки бывают комариные, слепневые. В связи с местами обитания водяной полевки и выплода комаров, отчасти и слепней, туляремия связана с поймами рек и с другими водоемами. Труп погибшей от туляремии водяной полевки, попадая в воду, заражает ее микробом туляремии; при стоянии голыми ногами в такой воде при рыбной ловле человек может заразиться этой болезнью. То же может быть и при бытовом использовании воды, например колодца, в который попал подохший от туляремии воробей. Но главное — это промысловые вспышки среди лиц, добывающих водяную полевку ради ее шкурки, имеющей хороший сбыт. Когда в руки промысловика попадают больные водяные полевки, то при снимании с них шкурок охотник или помогающие ему члены семьи легко заражаются туляремией. Бывают вспышки болезни совсем другого рода — сельскохозяйственные и в конечном счете совсем не связанные с водой. Туляремией
19*
заболевают и различные грызуны; больные животные выделяют возбудителя с мочой, загрязняют различные сельскохозяйственные объекты — скошенный хлеб, когда он лежит еще на полях. При собирании его руками работники могут заражаться туляремией. Еще легче это бывает, когда скошенный хлеб складывают в стога и ометы. В них легко поселяются и во множестве размножаются мелкие грызуны («мышиная напасть»). Когда среди них оказываются больныеособи, они загрязняют инфицированной мочой все окружающее, и при позднем обмолоте, когда поднимается зараженная пыль и люди ее вдыхают, появляются аспирационные сельскохозяйственные вспышки этой болезни. То же и при перекладке старых ометов. Бывают и алиментарные вспышки, когда убирают бобовые и инфицированные снаружи стручки поедают и дети, и работающие. В распространении туляремии имеют существенное значение и зайцы (например, в Чехословакии — Heyberger1Benda, Capek1 1956).
При обследовании одного пастбища в центральном районе Европейской части РСФСР Н. Г. Олсуфьев установил наличие на этом пастбище иксодо- вых, зараженных возбудителем туляремии клещей. Через 5 лет эго место было обследовано им вторично и снова были найдены клещи, спонтанно зараженные возбудителем туляремии. Значит наряду с дикими грызунами клещи и являются резервентами возбудителя туляремии.
Другая трансмиссивная болезнь — чума также относится к природноочаговым болезням потому, что ею болеют дикие грызуны: тарбаганы, суслики и другие грызуны. Тарбаганы и суслики являются промысловыми животными и люди, добывающие их для заготовки шкурок, подвергаются опасности заражения, когда в их руки попадают больные грызуны. Среди последних бывают целые чумные эпизоотии вследствие передачи возбудителя чумы через различных видов блох. Заражение чумой бывает и у грызунов, обитателей человеческого жилья, таких грызунов часто называют синантропами, но лучше их относить к группе «домовых обитателей».
Следует различать замкнутые биопатоценозы облигатно трансмиссивных природноочаговых болезней и смешанные или открытые биопатогеоцено- зы, которые характеризуются тем, что их компонент — возбудитель болезни (патоэргонт), не только циркулирует между донорами, переносчиками и реципиентами, но и закономерно переходит во внешнюю среду .(почву, воду и др.), где может существовать долгое время; поэтому люди могут заражаться такими факультативно-трансмиссивными болезнями и непосредственно из внешней среды. Структура природных очагов таких болезней более сложна, так как, кроме биокомпонентов, в структуру очага входят и абиотические компоненты неживой природы (почва, вода и др.).
Некоторые не трансмиссивные болезни также постепенно включаются в группу природноочаговых болезней. Таковы, например, лептоспирозы (заражение ими при купании в инфицированном искусственном и естественном водоеме). В Казахстане лептоспиры были выделены Крепкогорской из песчанок пустыни, а Я. А. Благодарным (1961) из сухопутных черепах Testudo Fiorsfieldi1а также из живущих в их норах клещей Ornithodoros tartakowskyi. Если для песчанок трудно установить какую-либо зависимость от водного фактора, то это совершенно невозможно сделать для спонтанно зараженных лептоспирами клещей Ornithodoros tartakowskyi. Ведь последние питаются только кровью своих хозяев-прокормителей; следовательно, и лептоспир они могут получать только от последних.
Эпидемиологические отличия трансмиссивных природноочаговых болезней во многом определяются биологическими особенностями клещей — переносчиков их возбудителей.
Для иксодовых клещей (семейство Ixodidae) существенное значение имеет количество хозяев-прокормителей, которых используют эти клещи в процессе своего жизненного цикла.
Так, Ixodes persulcatus, Ixodes ricinus являются треххозяинными клещами; личинки, нимфы, самцы и их самки питаются на трех различных хозяевах (млекопитающих или птицах). В каждой стадии жизненного цикла клещ (его личинка, нимфа и патовозрелая форма) сосет кровь по одному разу, но процесс сосания является продатжительным (до нескольких дней). Это определяет и передачу возбудителя клещом-переносчиком.
Если возбудителя болезни при сосании крови получает личинка клеща, то передавать возбудителя будет нимфа клеща, появляющаяся после линяния сытой личинки (сбрасывания хитиновой кожицы). После питания нимфа линяет на самца или на самку, которые сохраняют в себе вирус, бывший в нимфе; при питании половозрелых форм они и передают возбудителя живот- кому, кровь которого сосут; такова «трансфазовая передача» возбудителя. Она может быть и более короткой. Если зараженную кровь будет впервые сосать нимфа, то передавать полученного ею возбудителя будут после ее линяния самцы и самки.
Схематично изложенное можно представить в следующем виде:

Иксодовые клещи рода Dermacentor характерны тем, что их личинки нимфы питаются на мелких грызунах, а взрослые нападают на крупных животных и на человека. Поэтому схема циркуляции и передачи возбудителя иная:

Гаков путь трансмиссивной передачи клещевого риккетсноза (клещевого дынного тифа), который является природноочаговой болезнью, свойственной ландшафту степей Северной Азии. В степи люди заражаются также трансмиссивно весной от голодных самок Dermacentor (рис. 44).
В городских условиях люди могут заражать сами себя контаминативно. Iак и было с первым больным в Севастополе, которому А. Я. Алымов и поста- зил правильный диагноз. Это и было первым выявлением клещевого рик- ■ етсиоза в СССР.
Упомянутый больной лежал среди лета в больнице с диагнозом сыпного тифа. А. Я. Алымов заподозрил у него средиземноморскую сыпнотифозную лихорадку, рас- ^ своей собаки насосавшихся клещей. Сделать это, не раздавливая клещей, не всегда . дается, а при раздавливании пальцы загрязняются содержимым клеща. Если затем
касаться ими конъюнктивы глаза и слизистой оболочки носа, то имеющиеся в клеще риккетсии проникают через эти оболочки в организм и заражают его. Таков путь кон- таминативного самозараження клещевым риккетсиозом.
Открытие этой болезни на юге Крыма и затем в степи (рис. 45) под Красноярском экспедицией
Рис. 44. Скальный клещ солка Dermacentor — переносчик возбудителя клещевого риккетсиоза Северной Азии.
М. К. Кронтовской (Е. Н. Павловский. Н. В. Сергеев и Петрова-Пиоптковская, 1941) оказалось только началом. Последующими исследованиями многих авторов клещевой риккетсиоз был обнаружен в Центральной и Восточной Сибири, в Забайкалье, в Туве и Читинской области, в Хакасски, на Алтае, в Кемеровской и Новосибирской областях, в Казахстане, в Киргизии, в Биробиджане, в Хабаровском крае, на Дальнем Востоке, в Приморье. На севере он доходит до Якутска. При столь широком распространении клещевой сыпнотифозной лихорадки сменяются виды клещей ее переносчиков; кроме упомянутого выше Rhipicephalus sanguineus, в Северной Азии переносчиками являются Dermacentor Inarginatus1 D. pic- tus. D. Silvarum1в Киргизии и в Приморье— Haemaphysalis concinna. Последний вид клеща встречается на влажных кочкарных заболоченностях; клещ же Dermacentor—в степи, в степной части лесостепья; встречается и в таежной зоне на лесных полянах.

Рис. 45. Ландшафт степи Северной Азии — место заражения клещевым риккетсиозом.
Над изучением клещевого сыпного тифа в СССР работало много авторов, давая ему разные наименования: сибирский клещевой сыпной тиф, эндемичный тиф Сибири и Дальнего Востока, клещевой сыпной тиф Азии, Ixodo- Fickettsiosisasiatica (переносчиками возбудителя являются иксодовыеклещи), клещевой риккетсиоз Северной Азии, гамазовый риккетсиоз (переносчики - гама зовые кл егцй).
На одном из островов Южного Приморья Г. Сомову, М. Шапиро « А. Петрову пришлось столкнуться с естественной зараженностью риккетсиями гамазовых клещей — Hirstionyssus isabellinus и Haemolaelaps glas- gowi. Вопрос о передаче гамазовэго риккетсиоза нуждается в тщательном паразитологическом исследовании с учетом характера метаморфоза гамазовых клещей, их питания, не говоря уже о необходимости работы с отдельными и определенными видами этих клещей. Обычно же используют эмульсию смеси гамазовых клещей. Если при таком методе работы и выделяется возбудитель болезни, то это может быть лишь наводящим обстоятельством, требующим дальнейших исследований, так как заражение животного-реципиента в опыте с введением ему эмульсии из гамазовых клещей никак не моделирует ни того, что происходит в природе, ни способа, каким люди могут заражаться в конкретных условиях. Изучение клиники гамазового риккетсиоза продвигается вперед (И. Р. Дробинский, 1963).
В Японии распространена речная лихорадка, или краснотел ковы й сыпной тиф (переносчиками являются личинки некоторых видов краснотелко- вых клещей Trombidiidae); на юго-востоке Азии форма болезни с такими переносчиками называется кустарниковым тифом (scrub-typhus).
Иксодовые, гамазовые и краснотелковые клещи отличаются по своей биологии, распространению и роли как переносчиков возбудителей болезней, хотя они все, как и другие группы клещей, относятся к одному общему классу Acarina (тип членистоногих или Arthropoda).
Можно утверждать, что эпидемиологическое «лицо» трансмиссивных (и природноочаговых болезней) определяется именно биологическими особенностями их переносчиков (сезонность заболеваемости, ландшафты и другие места, в которых люди заражаются возбудителем, нозогеографня, меры неспецифической профилактики и др.). Поэтому паразитологическая точность методов их определения должна соблюдаться не в меньшей мере, чем вирусологические и микробиологические исследования.
Клещевые риккетсиозы являются природноочаговыми болезнями; природные очаги иксодовых риккегсиозов не переходят в непосредственную близость к человеку так же, как риккетсиоз, распространяемый краснотел- камн. Гамазовые клещи могут быть и в жилье человека и в служебных помещениях.
В отношении сезонности заболеваемости и места заражения трансмиссивными природноочаговыми болезнями можно высказать общее положение: заболевания могут быть только в то время, когда в природном очаге имеются голодные клещи-переносчики, и в тех местах, которые входят в границы ареала соответственного вида переносчика; но не в каждой точке внутри такого ареала непременно будут встречаться зараженные возбудителем передаваемой им болезни люди.
Клещевые риккетсиозы в общем имеют весенне-летнюю сезонность. IIa это время приходится наибольшее число заболеваний людей за весь эпидемический период (около 5 месяцев).
Клещи являются не только переносчиками, но и хранителями риккетсий в природе в межэпидемический период. Это их значение, помимо взаимной адаптации клещей и передаваемых ими возбудителей, зависит также и от формы передачи. Различают трансфазовую передачу, когда клещ, получивший возбудителя после превращения в следующую фазу метаморфоза (после линяния), нимфа от личинки; самка от нимфы (у иксодовых клещей) может передавать возбудителя.
Совершенство взаимного приспособления возбудителя болезни и его переносчика доказывается и другим способом —трансовариальной передачи.
Самка, так или иначе получившая возбудителя болезни, откладывает зараженные им яйца. Возбудитель нисколько не мешает эмбриональному развитию личинки переносчика и к моменту вылупления ее из яйца занимает в ней ворота выхода, т. е. достигает слюнных желез. Когда такая личинка — дочь инфицированной производительницы, впервые в своей жизни будет сосать кровь реципиента, она заразит его заключающимся в ней возбудителем; так же и дочерние нимфа и самка. Более того, трансовариальная передача распространяется и на внучатное поколение исходной инфицированной самки. Это в условиях опытов; возможно, что это не предел, но проведение таких опытов берет при треххозяинных клещах переносчика много лет.
Для клеща Dermacentor marginatus С. Пионтковская и О. Коршунова установили последовательную передачу риккетсий до четвертого нисходящего поколения, что продолжалось 5 лет и было сроком опытов, но не пределом передачи.
Следовательно, и для риккетсий клещи-переносчики являются действительными резервуарами возбудителя, который в них и зимует. Этим н объясняются ранние весенне-летние заболевания клещевым энцефалитом и клещевым риккетсиозом. Исходя из биологии иксодовых клещей, такое заключение было сделано теоретически автором и подтверждено обнаружением спонтанно зараженным вирусом клещевого энцефалита голодных самок Ixodes persulcatus, взятых в природе Дальнего Востока весной (Е. Н. Лев- кович и А. Н. Скрынник).
В. С. Ерофеев обнаружил естественную зараженность одной из партии 60 нимф того же вида, добытых 6 января 1963 г. из-под снега; вирус был выделен из клещей на белых мышах весом 8—10 г и из культуры тканей.
Известно, что некоторые микроорганизмы, поселившись в процессе эволюции в насекомых, клещах и в рыбах, становятся их постоянными симбионтами, потерявшими способность свободно существовать во внешней среде; они беспрерывно переходят из поколения в поколение своих «хозяев»; такие симбионты не являются паразитами, так как у глубоководных морских рыб они становятся частями их светвыделяющих органов («холодный свет»).
Нет ничего невероятного и в том, что патогенные вирусы, например вирусы клещевого энцефалита, взаимно сжившиеся с клещами-переносчиками, держатся в клещах не два поколения, а неограниченное, подобно симбионтам, число нисходящих генераций клещей.
Многие авторы делали попытки определения количества иксодовых клещей природного очага, спонтанно зараженных возбудителем клещевого энцефалита. Точные данные можно получить лишь при исследовании каждого клеща, но делать это практически невозможно (при сколько-нибудь крупных сборах) как по экономическим соображениям, так и по другим причинам (количество потребного персонала специалистов и подсобного персонала, затрата времени и др.). Поэтому исследователи вынуждены были применять условный метод.
Всю партию собранных клещей разбивали на одинаковые по количеству клещей группы (например, по 10—20—50 клещей) и каждую группу исследовали как одно целое, эквивалентное одному клещу, спонтанную зараженность определяли трансмиссивным или инокуляпионным методом. Процент зараженности обозначали в процентах по всем исследованным группам. Относительность получаемых данных очевидна. Ведь нет никакой гарантии в том, что соотношение зараженных и незараженных (будем в дальнейшем их называть стерильными) клещей во всех группах будет непременно одинаковым; неудобство заключается также и в том, что смешение сборов в природе клещей, собранных в разных биотопах, нивелирует возможность оценки эпидемиологического значения их биотопов для людей.
Следует иметь в виду, что результаты присасывания клещей-переносчиков бывают в разных местах и в разные годы явно различными.
Соотношение клинических и инаппарантных форм в одном и том же очаге по годам может существенно изменяться. Так, в одном из пунктов Кировской области в 1960 г. одна клиническая форма приходилась на 33 инаппарантных; в 1961 г. одна — на 76 инаппарантных и в 1962 г. одна — на 86,6 инаппарантных, что характеризует эпизоотологическую ситуацию соответственного года (В. Ф. Попов, 1963).
У населения, давно проживающего в местности, которой свойственны природные очаги клещевого энцефалита, происходит большее и большее заражение людей вирусом клещевого энцефалита, что чаще приводит к латентным и стертым формам клещевого энцефалита, но и при таких скрытых заболеваниях у многих лиц обнаруживаются такие признаки органического поражения нервной системы, как неравномерность глазных щелей, узость зрачков, ослабление реакции их на свет, асимметрия лицевой мускулатуры, односторонне ограниченная подвижность мягкого неба, отклонение языка, неравномерность рефлексов на конечностях, отсутствие брюшных рефлексов и др. (А. Шаповал, В. Коваленко, Сапельникова, 1963).
Так, при наблюдении в трех поселках Кемеровской области А. Шаповалом, В. Коваленко и Сапельниковой (1963) из 509 человек, к которым присасывались клещи, у 83 наблюдалось нарушение самочувствия в разных формах, у 45 человек было кратковременное повышение температуры; при неврологическом осмотре 458 человек были выявлены различные сочетания изменений обычно стойких неврологических симптомов; у 13 лиц развились клинические формы клещевого энцефалита. Следовательно, в природных очагах клещевого энцефалита население широко инфицируется, что приводит к развитию явных, стертых и латентных форм болезни.
При изучении природных очагов болезней приходится не только раскрывать их биопатоценологический состав, но и определять иммунологическую структуру.
Для очагов клещевого энцефалита в Томской области А. Явья (1963), применяя реакции связывания комплемента (PCK) и реакции подавления гемагглютинации ∣PΠΓA), получал у людей положительный эффект РПГА в 7,7—79,5% случаев, a PCK — в 28—46.4% случаев; у домашних животных при РПГА в 11,1—100% случаев, а при PCK — в 32—46% и у грызунов при РПГА — в 10,5—55,5% случаев. Совпадение положительных результатов PCK и РПГА колеблется в 7,1-—40%, что свидетельствовало о различной степени напряженности природных очагов клещевого энцефалита (1963).
Для определения количества людей, заражающихся в природных очагах, необходимо суммировать клинические и стертые случаи клещевого энцефалита, так как и последние возникают вследствие поступления вируса в организм человека.
Известно, что в сыворотке крови переболевших клещевым энцефалитом людей содержатся противотела к вирусу этой болезни. Реакцией нейтрализации сывороткой крови минимальных смертельных для белых мышей доз вируса пользуются для ретроспективной диагностики перенесенного заболевания, которое в свое время не было распознано.
Более того возможно обнаружение противотел к музейным штаммам вируса там, где явные заболевания еще не обнаруживались. Так, работа чехословацкой экспедиции в Албании установила наличие противотел в сыворотках крови к Западно-Нильскому энцефалиту, что говорит о наличии скрытых природных очагов данной болезни и о существовании этого вируса, неизвестного ранее для Европы.
Эпидемиологами комплексных экспедиций проводилось массовое обследование населения притаежных поселков на присутствие в сыворотке крови
противотел к вирусу энцефалита с опросом о перенесенной ранее «мозговой болезни». Оказалось, что у некоторых лиц, никогда не болевших энцефалитом, в сыворотке крови содержались противотела к вирусу этой болезни.
В связи с этим следует привести результаты экспериментальных работ Е. Н. Лев- кович и А. Н. Скрынник (1941) и автора совместно с В. Д. Соловьевым по циркуляции вируса энцефалита в клещах-переносчиках Haeniaphysalis Concinna (1941) и Ixodes persulcatus (1940). Оказалось, что часть белых мышей оставалась здоровой при введении им в мозг таких же доз взвеси из органов зараженного вирусом клеща, от которых большинство мышей в параллельном опыте погибало при классических проявлениях энцефалита.
Но при последующем введении в мозг незаболевшим белым мышам смертельной дозы вируса клещевого энцефалита эти мыши оставались здоровыми. Очевидно, введенная взвесь органов зараженного клеща иммунизировала часть мышей, которые ста новятся невосприимчивыми к смертельной дозе вируса.
Результаты этих опытов можно использовать и для объяснения сущно сти скрытой иммунизации населения там, где оно обычно контактирует с клещами. При обследовании сывороток крови на присутствие в них противотел, связывающих вирус клещевого энцефалита, были обнаружены такие противотела и у людей, которые не болели явной формой клещевого энцефалита. Очевидно, вирус от клещей они когда-то получали, но в дозе, недостаточной для заболевания. Такая доза и оказывала вакцинирующее действие на неболевших людей.
Можно полагать, что исход заражения определяется не только количеством передаваемого им клещами вируса. Причина различий в действии вируса на организм человека может быть скрыта и в качественных различиях штаммов вируса, могущих быть в клещах-переносчиках на одной и той же территории. Качественные различия штаммов вируса клещевого энцефалита могут зависеть от путей его предшествовавшей циркуляции в биоценозе, видовыми компонентами которого являются клещи-переносчики и различные позвоночные (млекопитающие и птицы), могущие содержать в крови вирус энцефалита. Для примера можно указать на следующие комбинации последовательного поступления штаммов вируса клещевого энцефалита в организмы клещей-переносчиков и в животных-реципиентов вируса в составе соответствующего биоценоза: бурундук — клещ Ixodes persulcatus; рябчик — клещ HaemaphysaIis coπciπna; волчонок — клещ и т. д.; клещ Ixodes persulcatus — еж амурский, клещ—полевка; клещ—дрозд и т. д.
Возникает вопрос, влияет ли на свойства вируса последовательность нахождения его в организме таких разных животных, как еж амурский, крот уссурийский, бурундук, рябчик, дрозд и др. при передаче его клещами- переносчиками разных родов. Так как последовательность передачи вируса энцефалита от животных-доноров животным реципиентам в природе варьирует, то и предшествовавшая «генеалогия» штаммов вируса оказывается различной; в результате штаммы вируса могут изменять свои свойства, в том числе и вирулентность для человека. «Генеалогия» штамма вируса — дело прошлое и восстановить ее ретроспективно нельзя, но считаться с возможностью влияния организма хозяина на находящийся в нем штамм вируса следует. При дальнейших экспериментах удастся установить, как качественно влияет штамм вируса на тот или иной вид хозяина, являющегося на некоторое время средой его обитания.
Не исключается влияние абиотических факторов внешней среды на биологические свойства переносчика и на находящегося в нем, как в пойки лотермном организме, возбудителя болезни.
Из факторов внешней среды прямое влияние оказывает температура; на примере клещей разные их виды проявляют в различных фазах своего
превращения далеко неодинаковую чувствительность. Как показывают опыты В. И. Алифанова (1963), температура воздуха оказывает влияние на продолжительность эмбрионального развития личинок в отложенных самкой клеща яиц, о чем можно судить по срокам появления личинок из яиц. отложенных самкой клеща. .
При температуре воздуха 4-22,9° личинки вылуплялись из оболочек яиц Ixodes persulcatus на 29,6 дня, а при температуре 4 15,9° этот срок увеличивался до 72,9 дня Сами личинки разных видов клещей неодинаково чувствительны к температуре окружающего воздуха; личинки Ixodes persulcatus могут оставаться живыми при —7°. тогда как личинки Dermacentor pictus погибают уже при понижении температуры до —I’.
Также по-разному влияет и влажность окружающей среды. Отложенные яйца Ixodes persulcatus крайне чувствительны к недостатку влаги и гибнут, тогда как в аналогичных условиях у клещей Dermacentor pictus и D. margiπatus гибель яйцекладок незначительна. Гибель или сохранность кладок естественно касается и возбудителя болезни, если он от самки пере носчика перешел в развившиеся в ее яичнике яйцо в порядке трансовариальной передачи возбудителя.
В настоящее время внимание медиков и биологов справедливо обращено на иксодовых клещей, которые играют бесспорно ведущую роль в лице видов — переносчиков вируса клещевого энцефалита и других болезней человека.
Только ли клещи принимают участие как переносчики в циркуляции возбудителя весенне-летнего энцефалита в природных и антропоургических очагах этой болезни? Во время виремии на людей, больных клещевым энцефалитом, нападают и комары, и наружные паразиты, в частности блохи. Несомненно, что при сосании крови бального находящийся в крови больного вирус поступает в желудок как комаров, так и блох. То же и в еще больше,м масштабе и в большем разнообразии совершается и в природе, когда на различных диких животных и на птиц, находящихся в состоянии виремии, нападает масса гнуса (летающие кровососущие двукрылые — комары, мошки, мокрецы, слепни) и различные неиксодовые клещи (гамазиды и др.). Хотя все эти кровососы и не являются переносчиками полученного ими виру са, все же следует выяснять хотя быслучайноеи крайне ограниченное участие их в поддержании циркуляции вируса в природных очагах болезни. Как указывалось выше, вирус, полученный в эксперименте комарами Aedes vexans и Aedes excrucians исчезает из них через 5 дней. За это время в их организме происходит закономерное снижение титра вируса (Е. Н. Левко вич и А. В. Гуцевич).
В. И. Ягодинскому и Б. И. Скворцову удалось в Хабаровском крае выделить вирус группы клещевого энцефалита из слепней Tabanus Huvi- corπis.
Нет ничего невероятного в том, что за короткое время переживания вируса в насекомых, не являющихся его переносчиками, такие насекомые могут принимать какое-то косвенное и случайное участие в частичной циркуляции вируса в его природном очаге.
3. Г. Кулакова (1962) предприняла исследование возможного значения блох мышевидных грызунов в циркуляции вируса клещевого энцефалита. Предпосылками к такой работе были случаи выделения вируса из блох грызунов и землероек в различных очагах клещевого энцефалита в Кемеровской области, в Приморье, в Беловежской пуще и экспериментальные данные, установившие способность разных родов и видов блох сохранять вирус клещевого энцефалита в течение различного времени (9 часов. 24 часа, 5 суток).
В своих первоначальных опытах 3. Г. Кулакова выяснила способность блох Ceratophyllus fasciatus и С. coπsimilis сохранять в себе вирус при температуре 4—25° в течение 6 ∙g∙ суток; а в течение первых суток после инфицирующего кормления блохи передавали вирус при кормлении здоровым мышам-сусликам (3. Г. Кулакова, 1962). Такая передача является, конечно, механической. Дальнейшие опыты привели к заключению, что в природных очагах клещевого энцефалита блохи могут играть некоторую роль в поддержании циркуляции вируса и способствовать созданию иммунной прослойки среди мышевидных грызунов особенно во время их размножения, с чем совпадает значительное возрастание численности блох в гнездах грызунов.
Такое заключение имеет значение для видов блох и грызунов, испытанных в эксперименте, но не может быть переносимо по аналогии на других кровососущих насекомых; каждая их группа должна исследоваться экспериментально, то же и для клещей разного систематического положения.
В Иркутске H. Н. Краминскому, P. Р. Живоляпнной, В. А. Перевозникову (1962) удалось выделить от парализованного голубя штамм вируса клещевого энцефалита и от гамазовых клещей Dermanyssus galIinae — штамм нейровнруса, который еще не идентифицирован.
Вирус клещевого энцефалита выделяли и другие авторы из смеси видов гамазовых клещей. Возможно, что и гамазовые клещи принимают некоторое участие в циркуляции вируса в природном очаге клещевого энцефалита. Кроме того, и роль самцов, и самок такого коренного переносчика вируса этой болезни, как Ixodes persulcatus, не одинакова.
Самцы Ixodes persulcatus так же, как и самка, нападают на теплокровных животных и на человека. Но самцы сосут кровь гораздо более короткое время, чем самка (до ll∕2часов), быстро вынимают из покровов погруженный в них хоботок и снова могут присасываться через несколько минут. Очевидно, им свойствен феномен прерывистого питания. Из самцов также удается выделить вирус клещевого энцефалита методами суспензии и кормлением на белых мышах. В. Н. Ягодинский (1962), который сообщает приведенные сведения, полагает, что приобретение населением иммунитета может зависеть именно от укусов самцов, которые при сосании крови вводят малые дозы вируса, недостаточные для вызывания болезни, но достаточно иммуногенные.
Исследования с клещами-переносчиками проводились обычно в теплое время года, когда они существуют и активны в открытой природе. Зимой же наступает «межиксодовый» период, когда клещей не удается обнаружить, кроме весьма редких случаев нападения их в декабре в пургу на охотников; кроме того, в более теплых по климату местах сытые самки Hyalomma зимуют на верблюдах в их шерсти.
При исследовании состояния нимфы клеща Ixodes, добытой из-под снега, была обнаружена естественная зараженность ее вирусом клещевого энцефалита, который зимует в клещах.
А. 3. Феоктистову (1962) удалось выделить вирус клещевого энцефалита из печени двух красно-серых полевок, отловленных 22 декабря, когда активных иксодовых клещей-перенос чи ков нет. А. 3. Феоктистов полагает, что в межиксодовый период возможна циркуляция возбудителя этой болезни и в зимний период, но через блох или гамазовых клещей.
Причины того, почему возбудитель болезни в одних случаях вызывает клиническую форму болезни, а в других приводит к скрытым заболеваниям, вероятно, различны. Изменения вирулентности вируса, количества и качества поступающего в организм человека штамма возбудителя имеют существенное значение. Исходя из теоретических соображений, нельзя обойти тот факт, что возбудитель болезни (как пример возьмем вирус клещевого энцефалита) в процессе своей циркуляции, обеспечивающей существование
природного очага, переходит при своей трансмиссивной передаче от донора через переносчика реципиенту через организмы не только разных видов млекопитающих, птиц и рептилий, но и кровососущих членистоногих. На каждой ступени своей циркуляции он попадает в какой-либо иной организм. Организм же, являющийся средой хотя бы и кратковременного обитания зируса, может оказывать на вирус разное влияние. Например, организм человека является для вируса тупиком. Попав в периферическую кровь, он некоторое время циркулирует р ней по телу (состояние виремии), в силу своей нейротропности он по нервной системе достигает головного мозга ■ поражает его разные части. Обратного выхода в кровеносную систему он не имеет. Поэтому человек-реципиент не может сделаться донором вируса для клеща-переносчика, и, кроме того, во время бывшей виремии клещ практически не мог получить вирус от человека и передать его дальше. Следовательно, соответственно штамм вируса не получает возможности выхода из человека-реципиента, который является для него тупиком. Но в период виремии на него могут нападать самки разных видов комаров; с кровью они -олучают и вирус; комар не делается переносчиком этого вируса, существующего в нем короткое время, по истечении которого уже нельзя обнаружить в нем следов вируса (Е. Н. Левкович и А. В. Гуцевич, 1941).
Если некоторые виды организмов оказывают в общем выражении губительное действие на вирус, то и организм заражающихся реципиентов столь различных, как птицы и млекопитающие, нельзя рассматривать как аналог химической пробирки, в которой известная реакция протекает с неизменной закономерностью. Нет ничего невероятного в том предположении, что на свойство вируса вызывать клиническое заболевание влияет организм животного, являющийсяхотя бы и временной средой обитания вируса, который передается клещами-переносчиками.
А. Н. Скрынник и Н. В. Рыжов (1941) в лабораторных экспериментах во время работ Дальневосточной экспедиции 1938 г. показали, что Dermacentor Silvarum также является переносчиком вируса клещевого энцефалита.
Однако О. Власова, Л. Гулимова, А. Опанасенко и М. Тюшнякова, работая з Юргинском районе Кузнецкой котловины (Кемеровская область) с клещами Dermacen- lor marginatus на открытых лугах, не могли выявить спонтанной зараженности их’ обычным методом заражения белых мышей. Позднее н.ми была взята смесь клещей Derma- centor pictus и D. marginatus; интрацеребральное заражение белых мышей клещевой эмульсией было безрезультатным при проверке на двух пассажах. Затем была взята двухсуточная культура клеток HeLa и заражена материалом от клещей. Через 6 дней последующего культивирования упомянутые исследователи заражали интероцереб- рально белых мышей тканевой жидкостью культуры. Мыши заболевали с параличами задних конечностей. Таким сложных! способом была выявлена спонтанная зараженность клещей рода Dermacentor, но осталось неизвестным, оба ли вида клещей имели в себе вирус, потребовавший для своего выявления применение такого особого метода, или вирусом обладал только один из видов (какой?) (1963).
Среди животных-реципиентов возбудителя болезни, относящихся к разным видам одного и того же рода, например Canis, одни не заболевают (собака — Canis faπιiliaris) клещевым энцефалитом, другие же (волчата — Canis lupus) гибнут от классической формы этой болезни при их лабораторном заражении. Но и на собаке могут встречаться вирофорные клещи Haema- phisalis Concinna. Это было обнаружено в Оборском леспромхозе на собаке, жившей при одной из лабораторий экспедиции 1938 г.
Некоторые животные-реципиенты одного и тодо же вида проявляют разную возрастную чувствительность к возбудителю, который открывается в некоторых случаях лишь при использовании для биологической реакции животных-сосунков.
Исходя из того, что возбудители природноочаговой трансмиссивной болезни циркулировали среди носителей в далеком прошлом, можно быть уверенным, что не только разные его штаммы, но и каждый штамм проходил не один и тот же путь сменяющихся в цепной последовательности узлов биоценотических связей, да еще подвергавшихся разным влияниям погодных и сезонных факторов внешней среды.
Учитывая растущее количество видов открываемых доноров и переносчиков клещевого энцефалита, можно привести теоретические примеры возможных цепей циркуляции вируса клещевого энцефалита:

Вся прошлая последовательность циркуляции вируса КЭ, или генеалогия штамма, несомненно не идентична у штаммов разных линий. Можно ли быть уверенным в том, что, проходя разные пути циркуляции, штаммы вируса остаются абсолютно неизменными в отношении своей вирулентности; и не от изменений ли ее (независимо от фактора дозировки) зависит проявление различных форм заболеваемости реципиента, который в свою очередь также не остается абсолютно неизменной средой, в которую поступает вирус на очередном этапе своей циркуляции?
«Генеалогия» вируса—это уже прошлый этап существования индивидуального штамма и раскрыть ее методов нет. Но можно попытаться в экспериментальном порядке получить примеры влияния проведения штам ма вируса через последовательно подбираемые систематически резко различные особи реципиентов, используя разные виды переносчиков, например по таким схемам:
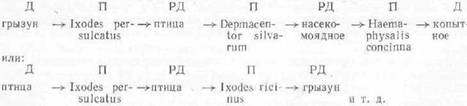
Опыты по примерному моделированию возможных цепей передачи вируса в природных очагах ставить нелегко, так как для их проведения необходимо обладать разнообразным ассортиментом экспериментальных животных, добытых из фауны природных очагов различных мест страны. Но такое положение в общем небезнадежно, так как многие научные и научно-практические учреждения широко размещены по территории государства.
Отом, что штаммы и бактериальных возбудителей природяоочаговых болезней неоднородны, можно судить по результатам выделения в 1960 и 1961 гг. в Забайкалье мных культур Л. А. Тимофеевым и Г. П. Апариным. Каждый штамм обладал индивидуальными, характерными для него особенностями в отношении вирулентности - J HMMVHoreHHbiM свойствам, по устойчивости к антибиотикам и в других отношениях 11962).
Степень восприимчивости реципиентов к возбудителям не является юстоянной; так, у грызунов, залегающих в зимнюю спячку, в период залегания резистентность повышается, благодаря чему возникают затяжные формы чумной инфекции. После весеннего пробуждения грызунов восприимчивость к их заражению повышается (Г. П. Апарин, 1963). По мнению Л. С. Малафеевой, особи грызуна одной и той же популяции обладают различной индивидуальной восприимчивостью и устойчивостью. У резистентных особей возникают затяжные формы чумы и такие особи обеспечивают длительное сохранение в природных очагах возбудителя чумы.
Кроме того, организм человека, млекопитающего, птицы в разных его органах (особенно кишечник) бывает заселен самыми разнообразными чуждыми для него микро- и микроорганизмами. Всю совокупность такого населения было предложено называть паразитоценозом (Е. Н. Павловский, 1934). Видовой состав последнего у одной и той же особи различен, что зависит от возраста, режима питания и других причин; у человека, в частности, и от факторов социального характера. В состав паразитоценоза, например пищеварительного тракта, входят в различных видовых сочетаниях спирохеты, риккетсии, бактерии, грибки, паразитические простейшие и паразитические черви. Многие виды из состава паразитоценоза безвредны для человека, но в его организм поступают и возбудители различных болезней, которые попадают в органы, заселенные сочленами паразитоценоза. Последние стоят в определенных межвидовых и внутривидовых соотношениях, причем весь паразитоценоз находится в состоянии взаимного влияния с организмом хозяина.
Характер соотношений между сочленами паразитоценоза применительно к влиянию его на хозяина троякий:
а) сочлены паразитоценоза находятся в индифферентных соотношениях друг с другом и безвредны для организма хозяина;
б) представители видов в составе паразитоценоза стоят в антагонистических отношениях с возбудителем болезни, попавшим в организм хозяина; вследствие этого затрудняется сосуществование патогенного для хозяина сочлена вида; при этом виды, безвредные для хозяина паразитоценоза, могут подавлять патогенное действие возбудителя болезни; в результате хозяин не заболевает болезнью, которую мог бы вызвать один возбудитель, действуя в одиночку;
в) возбудитель болезни, попавший в состав паразитоценоза хозяина, находит в нем таких сочленов, с которыми он не только уживается, но и проявляет с ними синергетическое действие или коакцию на организм хозяина (В. Г. Гнездилов, А. Ф. Тумка, A. AL Семенов, 1959; А. Г. Толстова, 1951; Е. Н. Павловский и В. Г. Гнездилов, 1953).
Особи паразитоценоза не обязательно должны иметь непосредственный контакт друг с другом; свое влияние на паразитоценоз и на восприимчивость организма хозяина к его действию оказывает центральная нервная система хозяина, поэтому влияние паразитоценоза на организм хозяина может быть рефлекторным.
Следствием таких разнообразных соотношений человек — обладатель паразитоценоза различного состава с включением возбудителя болезни, или заболевает типичной формой болезни, или же заболевание проявляется
с неясными симптомами недомогания, или он приобретает стертую, инаппа- рантную, латентную форму болезни; организм человека и животного может оставаться внешне здоровым, т. е. быть бессимптомным паразитоносителем, но заражающим выделяемым возбудителем внешнюю среду (почву, воду) и находящихся в ней здоровых людей.
При паразитировании круглых глистов (аскариды, анкилостомиды) лямблии в двенадцатиперстной кишке встречаются значительно реже, чем обычно, а при наличии ленточных червей они встречаются более часто (В. Г. Гнездилов). У детей до 4 лет, больных дизентерией Флекснера, лямблии встречаются почти в 2 раза чаще, чем у здоровых детей в той же местности; при дизентерии с высевом палочек Крузе — Зонне лямблии обнаруживались в 5 раз реже, а при паратифе (палочки Бреслау) их совсем не было (Полянская).
Очевидно, недостаточно ограничиваться только выделением от больного микроба — возбудителя болезни, необходимо в ряде случаев более широкое обследование больного с целью определения характера паразитоценоза. При этом можно будет учесть побочное влияние других сочленов паразитоценоза на характер заболевания. Об этом свидетельствует пример бездейственности сульфа н ил амидного лечения дезинтерии у людей, зараженных аскаридами, и положительный эффект лечения, полученный Чулковым, после изгнания последних.
Теорию паразитоценозов следует применять не только в клинике для уточнения диагноза и причин индивидуального течения болезни, но и при паразитолого-экологическом исследовании путей циркуляции возбудителя болезни в ее природном очаге, т. е. в донорах, переносчиках и в реципиентах возбудителя природноочаговой болезни. Другими словами, учение о природной очаговости болезней необходимо синтезировать с установками теории паразитоценозов.
Если биологические особенности переносчиков определяют «эпидемиологическое лицо» передаваемой ими болезни (сезонность, места и способ заражения, экстенсивность распространения болезни и др.), то самое открытие переносчика предопределяет меры профилактики соответственной болезни н борьбы с ней.
По своему существу меры профилактики разделяются на две группы: профилактика неспецифическая и профилактика специфическая.
Неспецифическая профилактика касается воздействия на переносчиков и на условия внешней среды, которые необходимы для их существования.
Предохраняя людей от трансмиссивных природноочаговых болезней, необходимо прежде всего поставить человека в такие условия, которые не давали бы возможности кровососущим переносчикам добраться до открытых частей тела или заползти под одежду и белье и присосаться к телу человека для питания его кровью. Среди массы нападающих переносчиков могут быть особи, естественно (спонтанно) зараженные возбудителем болезни, который передается человеку непосредственно при сосании его крови (т. е. инЬкулятивно) или же контаминативно.
Кроме мер индивидуальной профилактики, следует проводить и более общие мероприятия: засетчивание окон и дверей бараков, в которых живут временные рабочие; при палаточном жилье обрабатывают борта входа в палатку репелентами, отпугивающими клещей, или занавешивают вход в палатку портьерой из соответственного по размерам куска бредпевойдели, также пропитанной репелентами.
Самую территорию, которая должна быть занята под жилье в лесу, очищают от кустарника, снимают дерн; в степи ранней весной выжигают сухостой, при этом применяют огнеметы разных систем; в обжитых местах
нечерноземной полосы проводят мелиорацию пастбищ, применяя и методы воздействия арборицидами. .
Меры профилактики природноочаговых болезней в отношении обезвреживания территории, которой свойственны их очаги, различны и зависят от характера структуры таких очагов и более детально рассматриваются при описании самих болезней. Приведем лишь некоторые общие соображения. Борьба с грызунами имеет двоякое значение — в экономическом отношении как с вредителями сельскохозяйственных культур (например, суслики) и как с вредителями эпидемиологического значения (роль грызунов в чумных эпизоотиях и заболеваниях людей). Проведение обширных мероприятий по борьбе с такими «двойными» вредителями должно проводиться согласованными мерами с учетом интересов здравоохранения и сельского хозяйства.
Однако неизбежной остается' необходимость работы, постоянного или временного пребывания на территориях, имеющих природные очаги различных болезней. Обезвреживание таких территорий требует значительного времени; поэтому в данных случаях необходима специфическая профилактика, которая заключается в индивидуальной и особенно в массовой вакцинации людей, подвергающихся опасности заболевания природноочаговыми болезнями, если соответственные вакцины уже выработаны и могут быть применены в обиходе; для таких болезней, как чума, туляремия, холера, желтая лихорадка, вакцины уже имеются; для клещевого энцефалита они тоже есть, но научная разработка новых, более рентабельных в отношении использования вакцин продолжается.
Во всяком случае для профилактики природноочаговых болезней необходимо сочетание мероприятий как по неспецифической, так и по специфической профилактике.
Еще по теме ГЛАВА XlV ОСНОВНЫЕ ПОЛОЖЕНИЯ УЧЕНИЯ О ПРИРОДНОЙ ОЧАГОВОСТИ БОЛЕЗНЕЙ:
- Трансмиссивные природно-очаговые болезни
- 24. Основные положения учения В.Райха.
- 17. Основные положения учения Фрейда
- 28. Основные положения учения Перлса
- Природная очаговость клещевых инфекций
- Некоторые гипотезы о природной очаговости чумы
- Развитие методологических основ эпизоотологического обследования природных очагов чумы
- Оценка активности природных очагов чумы — основа их пространственной дифференциации.
- 22. Положения учения Альфреда Адлера
- Глава 2. Основные положения рационального питания
- Общие вопросы учения о патогенезе болезней
- Учения о причинах болезней в эпоху античности.
- Общие вопросы учения об этиологии болезней
- 8. Основные положения статьи Выготского «общие вопросы дефектологии» (основные тезисы и мысли по главам) (О. Леонович).
- 28 Характеристика основных категорий экоой психологии (мир природы, природная среда, биосфера, социогенез, антропоцентризм, экоцентризм, эко сознание).