Число жертв нападений блокированной блохи
В таком виде, как это звучит в заголовке данного подраздела, вопрос не ставился никогда. Тем не менее, сопутствующие ему проблемы рассматривались исследователями, но полнота этих исследований, как и их теоретическое обоснование, оказались весьма далекими от завершенности, поэтому есть смысл начать разговор об этом с самых общих рассуждений.
Есть достаточно много оснований полагать, что в природе вероятность «продуцирования» одним грызуном с бактериемией более одной блокированной блохи не всегда высока. Учитывая это, вряд ли найдутся возражения против того, что некоторая (возможно достаточно большая) часть блокированных блох должна заражать каждая более чем одного грызуна. В противном случае говорить о том, что при трансмиссивном эпизоотическом процессе идет нарастание числа зараженных носителей (хотя бы в отдельные, особо благоприятные для этого периоды) весьма проблематично. Поэтому существует необходимость выяснить, реальна ли вообще ситуация, когда среднее число носителей, заражаемых до состояния бактериемии одной блокированной блохой, превышает единицу. В тех редких случаях, когда ученые пытались получить подобные сведения, они не ставили вопрос перед собой столь остро. Очевидно, поэтому полученные в прошлом данные об этом так скудны и отрывочны. 7
Возможность заражения одной блохой нескольких грызунов должна быть оценена по двум позициям. Первая позиция (экологическая) характеризуется возможностью перехода блокированной блохи с одного зверька на другого в реальных условиях природного очага чумы, вторая (физиологическая) — вероятностью заражения каждого из животных, подвергшихся нападению инфицированного эктопаразита. В настоящей работе рассматривается только первая позиция. Как оказалось, вероятность многократной смены блохой своих прокормителей в природе в
208 количественном аспекте никем не изучалась. Причиной этому, на наш взгляд, служило не только и не столько отсутствие подходящих для этого методов, а главное — недооценка проблемы.
Известно достаточно большое число публикаций о «разносе» блох по поселениям различных видов грызунов, но приведенные в них данные не содержат информации о вероятности смены блохами своих прокормителей как о научном понятии или просто объекте исследования.То, что такая вероятность должна существовать, элементарно вытекает из образа жизни блох — временных (периодических) гнездовоноровых паразитов грызунов. Смена особей хозяев взрослыми насекомыми необходима для расселения вида (Кеннеди, 1978). Многократно нападая за свою жизнь на прокормителя, и столь же часто покидая его, блоха регулярно оказывается вне шерстного покрова хозяина, пережидая некоторое время в относительно стабильных гидротермических условиях его жилища. И нет никакой гарантии того, что в следующий раз она нападет на прежнего зверька. На этом этапе, когда объектом очередного нападения блохи может оказаться как старый, так и новый хозяин, и формируется «вероятность смены прокормителя». Буквально определение этой вероятности может быть сформулировано как отношение числа нападений со сменой прокормителя к общему их числу (не считая первого), совершаемых блохой в течение жизни или какого-либо ее отрезка. Разумеется, что для вычисления вероятности смены хозяина не обязательно следить за судьбой конкретной блохи всю ее жизнь, да это просто и не возможно. Но выяснить, какие факторы влияют на эту вероятность — задача выполнимая.
Для установления закономерностей смены прокормителей блохой были использованы те же графические модели, что и в подразделе 5.1.1. При этом к уже известной хронограмме условных нападений блох на грызунов добавили хронограмму присутствия в норе ее хозяина и других теплокровных «посетителей». Технология работы с графическими
209
моделями была следующей. На достаточно длинной (в несколько суток) шкале времени изображали цветными полосами чередующиеся между собой периоды нахождения в норе хозяина или посетителя, но каждые следующие сутки картина повторяла предыдущую. Это основано на высоком консерватизме суточной активности грызунов (Ердаков, 1984), которая меняется лишь по сезонам, поэтому периоды продолжительностью несколько суток вполне могут характеризоваться стандартным поведением зверьков.
На отдельной полоске бумаги изображали еще одну шкалу времени, на которой другими цветами выделяли периоды паразитирования блохи (tα) и промежутки между ними (Т - ta).Как уже говорилось, время однократного паразитирования и последующего отдыха незараженной блохи в норе или гнезде в сумме составляют один трофический цикл (7). Для блокированных блох понятие «трофический цикл» можно применить лишь условно, поскольку питаться в прямом смысле этого слова они не могут, хотя для них характерны многочисленные и более часто чередующиеся нападения с попытками кровососания при которых и может произойти заражение жертвы. Продолжительность жизни таких блох заметно сокращается и составляет в среднем 1-2 недели, редко превышая один месяц.На каждой хронограмме с тем или иным вариантом суточной активности теплокровных последовательно испытывалось несколько вариантов режима паразитирования блохи. Для этого на шкалу активности зверьков накладывали в случайном месте шкалу поведения насекомого. Смена хозяина могла осуществиться только в момент очередного нападения на прокормителя. Все зависело от того, в какой «норе» оказывалась блоха к этому моменту и какой зверек находился там в это время. «Двигаясь» вдоль шкалы, записывали, сколько нападений было со сменой и сколько без смены хозяина. Запись продолжали до тех пор, пока ситуация полностью не повторяла начальную, то есть когда блоха нападала на того же грызуна, в той же норе и в то же время суток.
210
Поскольку изготовленные варианты хронограмм были дискретными с «целочасовыми» показателями, возврат начальной ситуации происходил довольно быстро. Таким образом, вся количественная информация, заключенная между двумя идентичными состояниями системы, являлась одной генеральной совокупностью интересующих нас значений.
Самой простой изученной моделью была та, когда в некотором изолированном поселении имелось две норы с одним грызуном в каждой и одной блохой на двоих. Характер суточной активности грызунов был значительно идеализирован — в норе всегда присутствовал либо хозяин, либо посетитель.
Если в какой-то момент времени хозяин норы №1 выходил из нее, то сразу становился посетителем в норе №2, а хозяин второй норы в этот же момент становился посетителем в первой (происходил как бы обмен обитателями между норами). Возвращение хозяев в свои норы происходило так же одновременно. Варианты различались соотношением сроков нахождения зверьков «дома» и в «гостях». В этих вариантах каждые сутки были поделены на два сплошных отрезка, - тот, когда в каждой норе находился ее хозяин и другой, когда вместо хозяев в них были посетители. Именно на такую шкалу накладывали шкалу с трофическим циклом блохи. В данном случае предельное упрощение необходимо для обозначения «точки отсчета», четкое видение которой позволяет определить направление дальнейших исследований и программу усложнения моделей.Первоначально делались попытки проводить многократные испытания одних и тех же моделей, но с различными начальными точками совмещения шкал. Однако при этом всегда получали генеральную совокупность с одним и тем же количеством нападений и абсолютно совпадающим соотношением числа «смен» и «несмен», поэтому в дальнейшем мы ограничились только одним вариантом совмещения шкал. Каждой комбинации параметров соответствовали данные, состоящие из числа смен и несмен прокормителя, сумма которых принималась за
211 единицу, а отношение числа смен к этой сумме означало вероятность смены хозяина (Pm)при данных экологических параметрах.
В результате анализа всего массива данных было выяснено, что из показателей поведения блохи на смену хозяина оказывал влияние только интервал между периодами ее паразитирования (Т - ta),а варьирование продолжительностью ta(и, соответственно, 7) при неизменном T - taне влияло на вероятность перехода блохи на другого грызуна. При одних и тех же параметрах паразитирования блохи на исследуемую вероятность оказывало влияние соотношение периодов нахождения в норе хозяина или посетителя. Исходя из этого, была составлена таблица 15, графы которой соответствовали длительностям интервалов между периодами паразитирования (Т- ta),а горизонтальные строки — продолжительностям пребывания в норах грызуна-посетителя (tg)в течение одних суток.
Число граф этой таблицы могло быть сколь угодно большим, в зависимости от того, какой длительностью T - taмы пожелали бы ограничить испытания. Однако слишком большого числа граф не потребовалось, поскольку была обнаружена четкая цикличность в изменении вероятности смены прокормителя блохой.Максимальную величину эта вероятность могла приобрести всякий раз, когда интервалы между периодами паразитирования блохи были равны п + 0.5 суток, где п — целое натуральное число, включая 0. Если эти интервалы равнялись целому числу суток, то вероятность смены хозяина была равна 0 при любой суточной активности грызунов. Именно эти особенности обуславливают циклическую изменчивость изучаемой вероятности. Кроме того, число строк в таблице оказалось возможным сократить с 24 до 12, поскольку весь смысл статуса хозяина норы заключается в том, что он не может находиться в норе меньшее время, чем посетитель и наоборот.
Таблица 15. Зависимость вероятности смены прокормителя блохой от параметров паразитирования и суточной активности хозяев
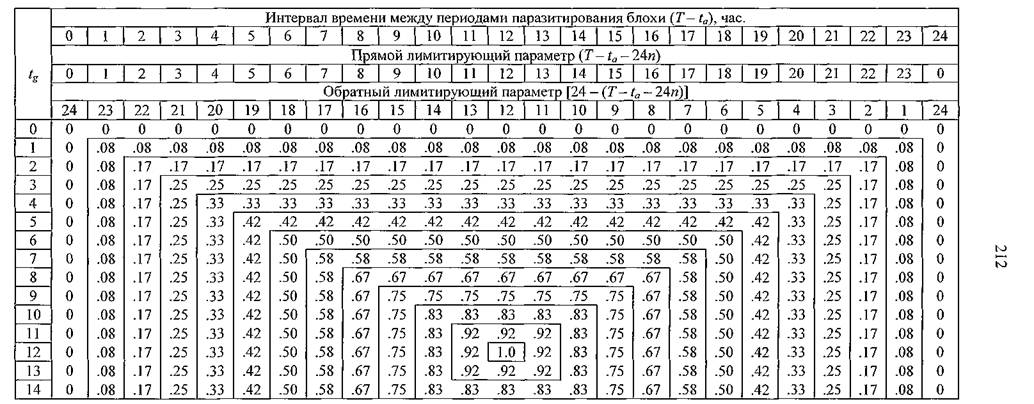
Примечание. Вероятности указаны только в знаках справа от запятой; tg— продолжительность нахождения в норе грызуна-посетителя; остальные обозначения в тексте.
213
Это обстоятельство нашло отражение в таблице 15 — при возрастании времени «визитов» вплоть до 12 часов, вероятность смены прокормителя плавно увеличивалась от 0 до 1, а при дальнейшем увеличении времени пребывания грызуна в чужой норе вплоть до 24 часов эта вероятность так же плавно снижалась до 0 (табл. 15). В этом случае происходила автоматическая смена статуса гостя на статус хозяина норы и наоборот. При том соотношении, когда каждый зверек одинаково долго пребывал в своей и чужой норе, и когда продолжительность T - taблохи равнялась п+ 0.5 суток, вероятность смены прокормителя становилась равной 1.0. Естественно, в природе эти крайние значения вероятности (0 и 1) никогда не могут быть реализованы благодаря лишь статистическому разбросу параметров в популяциях грызунов и блох.
Например, отклонение интервала между периодами паразитирования блох от среднего значения, равного целому числу суток, как в сторону уменьшения, так и увеличения, дает одинаковое повышение Pm.Снижение Pmпроисходит при отклонениях длительности указанного интервала от значения в п+ 0.5 суток.Анализ периодически изменяющейся вероятности перехода эктопаразитов с одного зверька на другого позволил вывести три ограничивающих эту вероятность параметра (L1; Ly L3). Как оказалось, любой из них, но только один - минимальный, определяет точное значение искомой вероятности, которая вычисляется путем деления удвоенного наименьшего параметра, измеренного в часах, на 24 (продолжительность суток): 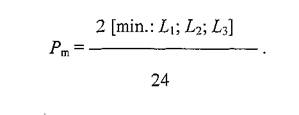
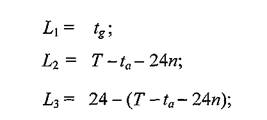 214 где tg— продолжительность нахождения в норе грызуна-пришельца, которая не может превышать таковую хозяина норы и в сумме с ней должна составлять 24 ч, п — целое число суток, укладывающихся в интервал времени T - ta.Второй и третий параметры, вытекающие из длительности одного непаразитического периода в трофическом цикле блохи, также дополняют друг друга до 24 ч. Наблюдаемый эффект минимального параметра можно сравнить с работой наиболее мелкого сита. Если просеивать песок сложного механического состава, используя несколько сит, различающихся размером ячеи и расположенных друг над другом, то преодолевать эту преграду смогут только частицы, проходящие через самое мелкое сито, каким бы по очереди оно не стояло.
214 где tg— продолжительность нахождения в норе грызуна-пришельца, которая не может превышать таковую хозяина норы и в сумме с ней должна составлять 24 ч, п — целое число суток, укладывающихся в интервал времени T - ta.Второй и третий параметры, вытекающие из длительности одного непаразитического периода в трофическом цикле блохи, также дополняют друг друга до 24 ч. Наблюдаемый эффект минимального параметра можно сравнить с работой наиболее мелкого сита. Если просеивать песок сложного механического состава, используя несколько сит, различающихся размером ячеи и расположенных друг над другом, то преодолевать эту преграду смогут только частицы, проходящие через самое мелкое сито, каким бы по очереди оно не стояло.
Таким образом, мы выяснили, что теоретическая вероятность смены прокормителя блохой зависит от двух величин: продолжительности ежесуточного пребывания грызунов в норах своих соседей (или «ничьих» норах) и продолжительности интервала между двумя периодами паразитирования блох. Подобная зависимость вполне понятна и на чисто описательном уровне — чем дольше зверек находится в чужой норе, тем выше вероятность того, что он оставит там «своих» блох и приобретет «чужих». Если интервал между двумя периодами паразитирования блохи равен целому числу суток, то, покинув прокормителя в конкретное время суток, она должна совершить новое нападение в такое же время через п суток. Скорее всего, в норе в этот момент окажется тот же самый зверек благодаря его стандартной суточной активности. Естественно, возможности поменять хозяина при этих обстоятельствах у блохи не будет никогда. Любое же отклонение от упомянутого интервала в п суток будет повышать шанс блохи напасть на другого зверька, но до определенного
215 предела, величина которого находится посередине между интервалами в п и п+ 1 (или п - 1) суток.
Следует заметить, что описанная выше зависимость носит функциональный характер, и это наглядно видно лишь на простой модели, лишенной всех модифицирующих факторов, накладывающихся на процесс смены хозяев. В природе эта зависимость, безусловно, существует. Однако две величины, формирующие вероятность смены прокормителя, зависят в свою очередь от многих природных явлений, что маскирует значение установленной зависимости в реальных поселениях зверьков.
Эти закономерности выявлены при испытании модели, представляющей систему из 2 нор, 2 грызунов и 1 блохи. Для выяснения того, сохранятся ли подобные закономерности при увеличении размеров системы, были проведены испытания других моделей, включающих до 7 нор с таким же количеством грызунов и до 24 эктопаразитов. Первая выявленная при этом закономерность заключается в отсутствии зависимости вероятности смены хозяина блохами от их числа. Фактически, то же самое было отмечено и в простых моделях, когда варьирование совмещением шкал поведения блохи и грызунов не влияло на генеральную совокупность из числа смен и несмен прокормителя. Точно также увеличение количества насекомых, нападающих в различные моменты времени на любых зверьков в созданной нами системе, приводило лишь к суммированию абсолютно идентичных генеральных совокупностей, что, естественно, не могло отразиться на среднем значении искомой вероятности. ,
При испытании следующих, более сложных моделей был сохранен принцип постоянного присутствия одного из грызунов, — либо хозяина, либо посетителя, в норе. Каждая нора характеризовалась тем, что ее хозяин находился в ней 12 и более часов, а остальные зверьки модельного поселения по очереди посещали эту и другие чужие норы в оставшееся время. Было испытано 2 варианта очередности посещения. В одном случае
216 каждые сутки в конкретной норе появлялся лишь один посетитель — сегодня один, завтра другой и т. д. В другом случае время, отведенное для посещений, ежесуточно делилось между всеми соседями. Совмещенную шкалу нападений при обоих вариантах посещений продлевали до абсолютного повторения первоначальной ситуации, то есть исследовалась вся генеральная совокупность случаев.
В результате анализа нескольких усложненных моделей было выяснено, что на вероятность смены прокормителя не оказывает влияния не только количество блох, но и число задействованных в системе грызунов и нор. Не имеет значения и характер очередности посещений нор посторонними зверьками. Решающими остаются все те же отмеченные ранее временные параметры: продолжительность пребывания в норе грызунов-посетителей (независимо от их количества) и интервал между периодами паразитирования блох.
Генеральная совокупность из числа смен и несмен прокормителя могла быть получена двумя способами. Первый описан выше и заключался в просмотре длинной шкалы времени между двумя повторившимися идентичными по всем характеристикам нападениями одной задействованной в модели блохи. Второй способ получения генеральной совокупности заключался в использовании большого числа блох, моменты первых нападений которых случайно распределялись в течение суток. Результаты второго нападения каждой блохи образовывали генеральную совокупность и давали максимально точное значение вероятности смены прокормителя, если число блох было равно или кратно 24. Анализ результатов испытаний модели, полученных этим способом, позволил также сделать вывод о том, что в природе интересующая нас вероятность, как и предполагалось, реализуется каждый раз по прошествии одного трофического цикла блохи.
Следует заметить, что отсутствие зависимости между вероятностью смены хозяина блохой и числом ее прокормителей, обитающих в
217
окрестностях, справедливо лишь при неизменном соотношении времен пребывания в норе хозяина и гостей. Однако само это соотношение не может не зависеть от плотности грызунов (и числа гостей) в поселении, но это уже другой вопрос, выходящий за рамки данного исследования.
Слабым местом изученных моделей является допущение о постоянном пребывании в норе теплокровных прокормителей блох. Известно, что грызуны определенное время проводят на поверхности земли, а в чужие норы заглядывают обычно на непродолжительный срок.
■ В связи с этим было рассмотрено некоторое количество моделей C перерывами в присутствии зверьков в норах. При этом были замечены следующие закономерности. Пребывание прокормителя с находящимися в его шерсти блохами вне норы заставляло дольше необходимого паразитировать этих насекомых и, естественно, удлиняло среднюю продолжительность однократного паразитирования блох в системе. Это же обстоятельство увеличивало продолжительность интервала между паразитическими фазами для блох, находящихся во временно пустующей норе. Чем дольше грызуны отсутствовали в норах, тем существенней увеличивались различные отрезки трофического цикла блох. Кроме того, наличие периодов, когда одни кровососы не могут покинуть прокормителя, а другим не на кого нападать, синхронизирует их активность между собой и привязывает моменты нападений на зверька и ухода с него ко времени его возвращения в нору. Принципиальным становится то обстоятельство, кто первым заходит в нору после отсутствия в ней теплокровных обитателей — хозяин или посетитель. Возвращение зверька после «прогулки» в свою же нору должно несколько снижать вероятность смены хозяина, а посещение сначала чужой норы и лишь после этого уход в свою нору — увеличивает эту вероятность. Указанные обстоятельства практически лишают нас перспективы выработки универсального и достаточно простого алгоритма вычислений искомой вероятности, учитывающего все возможные нюансы образа жизни
218 носителей и переносчиков возбудителя чумы. Однако анализ упрощенных моделей позволил наглядно представить, какие именно характеристики поведения хозяев и паразитов влияют на смену прокормителей. Кроме того, в резерве остается способ просмотра хронографической модели, но уже с реальными, измеренными в природных и лабораторных условиях, параметрами активности грызунов и блох.
Выявленные закономерности, обеспечивающие ту или иную вероятность смены хозяина блохой, позволяют максимально сузить круг вопросов, на которые нужно ответить при измерении необходимых параметров. По сути дела их имеется только два. Первый вопрос касается режима эксплуатации норы (городка или колонии) как ее исконными теплокровными обитателями, так и забегающими зверьками. При этом требуется знание среднестатистического режима, то есть наиболее характерного для обитаемой норы данного вида хозяина. Второй вопрос связан с параметрами паразитирования интересующих нас периодических (временных) эктопаразитов. Их измерение может быть осуществлено в лабораторных условиях и такие попытки применительно к блохам уже предпринимались (Новокрещенова, Солдаткин, Левошина, 1968; Ширанович, Чумакова, 1961; Медзыховский, 1971 а). Однако следует подчеркнуть, что важен хронометраж активности не только и не столько «здоровых», сколько блокированных блох, без чего окончательное вычисление коэффициента трансмиссивности чумы вряд ли осуществимо.
Итак, изучение хронографических моделей поведения грызунов и блох, прежде всего, открыло перспективу вычисления вероятности смены прокормителя паразитами. При этом выяснено, что изученная вероятность зависит не от численности грызунов и блох, а от суммы времени пребывания среднестатистического зверька в норах своих соседей того же или близких видов в течение суток и от средней продолжительности интервала между периодами паразитирования у конкретных видов блох (отдельно для интактных и блокированных особей). Эта вероятность в
219 природных условиях характеризует соотношение двух фиксированных состояний паразитарной системы «грызуны-блохи», разделенных промежутком времени, равным среднему трофическому циклу данного ’ вида блохи. Первое состояние, в котором конкретные экземпляры блох паразитируют на конкретных хозяевах, принимается за исходное. В следующем состоянии определенная часть насекомых, соответствующая вероятности смены хозяина, при повторном нападении паразитирует уже на других особях. При третьем нападении за исходное состояние принимается предыдущее (второе) паразитирование и т. д.
Из этого описания вытекает путь получения вероятности смены блохой прокормителя — наблюдение за мечеными эктопаразитами в природе. Для этого необходима разработка такого метода мечения блох, при котором насекомые могут получать индивидуальные номера и возможность неоднократно регистрироваться в процессе переходов с одного хозяина на другого. Такой метод был нами разработан и применен при исследованиях в природном очаге чумы в 1991-1994 гг. Результаты исследований с использованием метода индивидуального мечения блох представлены в разделе 5.2. Здесь же отметим, что независимо от того, каким способом измерена вероятность смены, она непосредственно участвует в формировании среднего числа грызунов, на которых может напасть блокированная блоха в природе.
Как явствует из определения только что упомянутого показателя, важно знать также общее число нападений, которые способна совершить блокированная блоха за оставшийся отрезок ее жизни. Вопрос о возможном числе нападений, как и о числе жертв, не изучался никем. Некоторые исследователи ежедневно пересаживали отдельные экземпляры или группы зараженных блох на новых зверьков, но это никак нельзя признать изучением поднятого вопроса. Почему, например, блохи пересаживались именно через сутки или двое и почему после пересадки они содержались на здоровом зверьке полчаса, час или другое, более
220 короткое или длительное время? Бесспорно, что эти временные интервалы выбраны из соображений удобства выполнения работы по пересадке блох экспериментатором. При этом практически ни в одной публикации не обнаружено даже попыток ответа на вопрос, — с какой частотой «блокированная» блоха сама совершала бы нападения на хозяина и уходы с него. Только в одной работе (Новокрещенова, Солдаткин, Левошина, 1968) проведено сравнение частот питания «блокированных» и «неблокированных» блох, показавшее практическую их идентичность. Признать это как окончательный вердикт трудно, к тому же и здесь не получено ответа на вопрос, сколько же раз в среднем за свою жизнь блокированная блоха может нападать на хозяина.
Исходя из знания среднего числа нападений, совершаемых блокированной блохой, и вероятности смены ею хозяина, можно было бы предположить, что среднее число разных зверьков, на которых может напасть такая блоха, должно быть равно произведению двух этих показателей. Однако это не совсем так. Для правильного определения среднего числа прокормителей, на которых может напасть инфицированная блоха в природе, необходимо учесть еще некоторые обстоятельства. Известно, что состояние бактериемии при чуме наступает у чувствительных к этой инфекции особей и практически однозначно свидетельствует о быстрой и неминуемой их гибели. Следовательно, блохи, питавшиеся кровью погибшего впоследствии зверька, не смогут напасть на него вновь, то есть остаться «не сменившими хозяина». Это означает, что первое после гибели хозяина нападение должно осуществляться на нового грызуна с вероятностью смены, равной или почти равной 1.0. Возможно, некоторые зараженные блохи могут остаться «невостребованными», если в ту нору, где они находятся, не заглянет ни один зверек вследствие, например, резкого снижения их численности, либо носитель с находящимися на нем блохами погибнет вдали от нор, где встреча паразита с другими прокормителями маловероятна. Однако есть
221
все основания считать, что осуществимость таких ситуаций пренебрегаемо мала.
Таким образом, можно смело утверждать, что после гибели носителя от чумы все или почти все блохи, питавшиеся на нем во время бактериемии, будут затем паразитировать на новых зверьках. Как известно из лабораторных экспериментов последующие после заражающего кормления «подкормки» являются необходимым, хоть и не единственным условием для образования чумного блока преджелудка у блох. При этом не важно, на одном зверьке или на нескольких осуществляется эта подкормка в природе. Число прокормителей будет иметь значение для трансмиссии лишь с момента образования блока. Можно считать, что первая с этого момента попытка питания блохи на здоровом зверьке обязательно реализуема, а последующие переходы блокированного насекомого на новых хозяев, осуществляемые с некоторой вероятностью, будут участвовать в формировании числа возможных реципиентов инфекции. Среднее число различных особей, подвергающихся нападению блокированной блохи (N^t),несколько больше простого произведения возможного числа ее нападений (А) и вероятности смены прокормителя (Pm)и выглядит так:
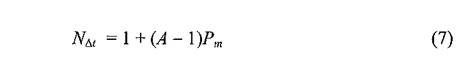
где N⅛ — среднее число особей, подвергшихся нападению; А — среднее число нападений блокированной блохи (A ≠ 0); Pm— вероятность смены прокормителя блохой.
Из этой формулы следует, что число «кусаемых» блохой грызунов не может быть меньше одного, даже при Pm,равной 0, а при Pm,равной 1.0, это число будет равно А. Если число потенциальных нападений блохи (А) разделить на число зверьков, подвергающихся нападению (Nf),мы узнаем — сколько раз в среднем блоха нападает на одного из них. Это число, особенно если оно дробное, само по себе мало что говорит о возможных
222 комбинациях количеств укусов, приходящихся на каждую жертву при нападениях. Крайние варианты комбинаций, это когда каждое нападение блокированной блохи совершается всякий раз на нового, не подвергавшегося ранее укусам этой блохи зверька, или когда все нападения совершаются только на одну особь прокормителя.
При анализе результатов подстановок различных значений в формулу (7) обнаруживается следующая закономерность: чем меньше различных зверьков подвергаются нападению инфекциозной блохи, тем большее число нападений (укусов) приходится на одного из них (при одном и том же А). Это существенно повышает шансы блохи передать этому зверьку инфекцию со всеми вытекающими отсюда последствиями, но сокращает число возможных разветвлений процесса. Это еще раз подтверждает то, что решающим для трансмиссии показателем является число активных нападений, совершаемых блокированной блохой за оставшийся период ее жизни, продолжительность которого в значительной мере определяет это число.
223
5.1.4.
Еще по теме Число жертв нападений блокированной блохи:
- Жертва и потерпевший, понятие и анализ
- Методика остеосинтеза блокированными стержнями.
- Подстерегание и нападение
- Число блох, питающихся на одном прокормителе в период бактериемии
- е) Число охваченных паллиативной помощью
- Блохи и их хозяева.
- БЛОХИ (OTP. SUCTORIA)
- Технические средства биологического нападения
- АБСОЛЮТНОЕ ЧИСЛО БОЛЬНЫХ С ВПЕРВЫЕ В ЖИЗНИ УСТАНОВЛЕННЫМ ДИАГНОЗОМ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ
- Блохи как переносчики чумы.
- Блохи и эндемический сыпной тиф.