2.2. Изучение статуса метилирования CpG-островка гена /ЗА-адаптина в опухолях шейки матки и в клеточных линиях рака шейки матки.
Чтобы выяснить, как связана транскрипция гена /ЗА-адаптина с метилированием, мы исследовали статус CpG динуклеотидов, входящих в состав его CpG-островка, в клеточных линиях HeLa и SiHa.
Для этого мы использовали метил-чувствительный ПЦР со специфическими праймерами (МЧ-ПЦР), гибридизацию по Саузерну и определение нуклеотидной последовательности бисульфитно-модифицированной ДНК. Как и в случае СП-ПЦР первые два метода подразумевают обработку ДНК метилчувствительными ферментами. При этом исследуются CpG динуклеотиды, входящие в состав сайтов рестрикции МЧР. Наибольшее число сайтов рестрикции в составе CpG островка гена /ЗА-адаптина наблюдается для ферментов HpaII (10) и HhaI (8). Статус метилирования сайтов рестриктаз HhaI и SacII мы изучали с помощью МЧ-ПЦР, рестриктазы HpaII - блот-гибридизацией.Расположение сайтов рестрикции фермента HhaI и использованных в работе праймеров представлено на рисунке 17А. В случае применения праймеров 26-13 и 26-8, мы определяли статус метилирования CpG динуклеотидов в составе семи сайтов. Результат этого исследования представлен на рисунке 17Б. Отсутствие продуктов ПЦР при амплификации ДНК, обработанной рестриктазой HhaI, говорит о том, что хотя бы в одном из исследованных сайтов рестрикции CpG динуклеотид не подвергается
- 79 -
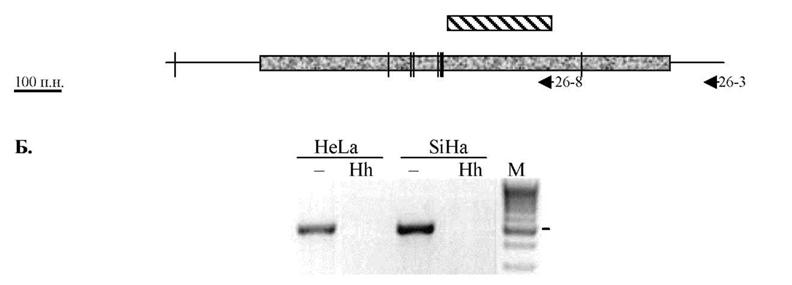 |
метилированию. Дробление первоначальной зоны исследований на более короткие отрезки с меньшим числом сайтов рестрикции (использовали комбинацию праймеров 26-14 и 26-8; 26-1 и 26-3) и, таким образом, уменьшение числа исследуемых за один раз CpG динуклеотидов ситуацию не изменило. Мы всегда наблюдали отсутствие продукта ПЦР, т.е. сайты рестриктазы Hhal оказывались рестрицированными. Таким образом, при исследовании статуса метилирования сайтов рестрикции фермента Hhal в границах CpG-островка гена вЗА-адаптина мы установили, что CpG динуклетиды, входящие в состав этих сайтов не метилированы в клеточных линиях HeLa и SiHa.
А. 26-13>- 26-14> 26-1>-
500пл. I -300пл.
Рисунок 17. Анализ статуса метилирования CpG динуклеотидов, входящих в состав сайтов рестрикции метилчувствительного фермента Hhal в пределах CpG-островка гена вЗА-адаптина, с помощью МЧ-ПЦР. А. Схематическое изображение расположения сайтов рестрикции фермента Hhal (вертикальная черта). Стрелками обозначены использованные праймеры; ? - CpG островок, И - 1 экзон. Б. Результат электрофореза в агарозном геле продуктов амплификации ДНК из клеточных линий с помощью праймеров 26-13 и 26-8. Рестрикция образцов ДНК Hhal (Hh) или без рестрикции (-). М - маркер 100 bp. Числами справа указаны размеры маркерных фрагментов.
Для изучения статус метилирования CpG динуклеотидов в сайтах рестриктазы HpaII мы использовали метод гибридизации ДНК по Саузерну. Здесь мы обрабатывали ДНК из клеточных линий метилчувствительной рестриктазой HpaII и, дополнительно, ферментом рестрикции Mspl, который является изошизомером HpaII, но не чувствителен к критическому для рестриктазы HpaII метилированию. При этом мы дополнительно исследовали
- 80 -
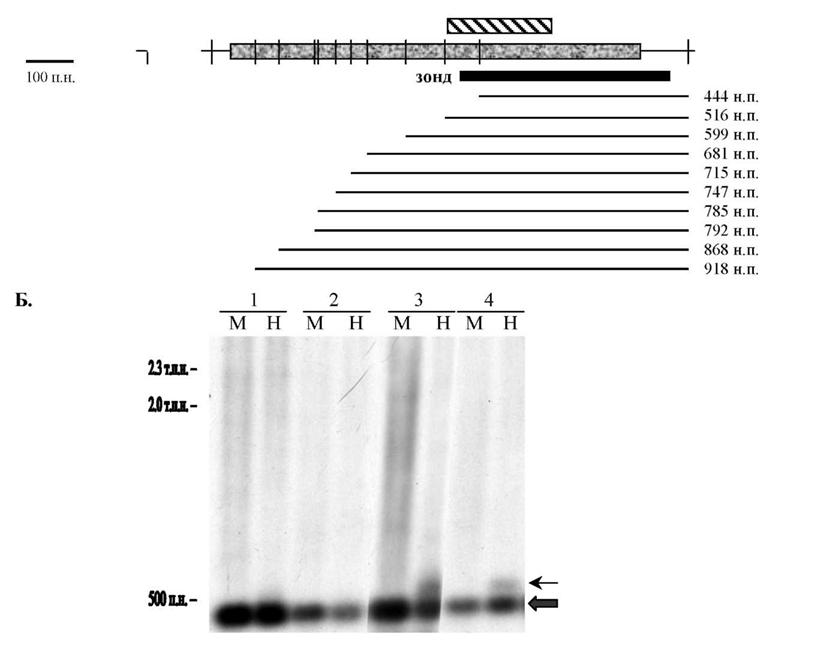 |
ДНК из клеток культур HeLa и SiHa, обработанных 5-азацитидином. Зонд для гибридизации представлял собой продукт ПЦР, который мы получили с помощью амплификации геномной ДНК с праймерами 26-1 и 26-3. На рисунке 18А представлено расположение сайтов рестрикции HpaII/MspI в исследуемом регионе гена и набор вариантов продуктов гибридизации,
А. I______
Рисунок 18. Анализ статуса метилирования CpG динуклеотидов, входящих в состав сайтов рестрикции метилчувствительного фермента HpaII в пределах CpG островка гена вЗА-адаптина, с помощью гибридизации по Саузерну. А. Схематическое изображение расположения сайтов рестрикции (вертикальная черта) по отношению к CpG островку -I | . Горизонтальными линиями обозначены варианты продуктов гибридизации. Число справа обозначает размер продукта.
И - 1 экзон. Б. Результат гибридизации культур клеток рака шейки матки до и после обработки 5-азацитидином. Числами вверху рисунка обозначены: 1 - Hela, 2 - Hela+5azaC, 3 - Siha, 4 - Siha+5azaC. Рестрикция образцов ДНК HpaII (H) или MspI (M). Числами слева указаны размеры маркерных фрагментов.которые будут наблюдаться при том или ином распределении метилирования сайтов рестриктазы HpaII. Рестрикция ферментом MspI является контролем, так как при этом образуется единственный продукт минимального размера 444 н.п. Результат гибридизации ДНК из клеточных культур представлен на рисунке 18Б. Видно, что в клетках HeLa при рестрикции как HpaII, так и MspI наблюдается один и тот же продукт (отмечен толстой стрелкой), что говорит об отсутствии метилирования сайтов HpaII в этих клетках. В случае клеточной линии SiHa возможно частичное метилирование одного-двух CpG динуклеотидов, так как при рестрикции ДНК ферментом HpaII отмечается дополнительная полоса большего размера, чем основной продукт (отмечен тонкой стрелкой). Таким образом, при изучении статуса метилирования CpG динуклеотидов в составе сайтов рестрикции фермента HpaII мы установили, что в клеточных линиях HeLa и SiHa большинство CpG динуклеотидов не метилировано.
CpG-островок гена вЗА-адаптина мы обнаружили с помощью СП-ПЦР по метилированию сайтов рестриктазы SacII. Поэтому представлялось возможным, что транскрипция гена вЗА-адаптина регулируется метилированием сайта какого-либо специфического транскрипционного фактора, содержащего сайт рестриктазы SacII. В последовательности CpG-островка гена вЗА-адаптина присутствуют два сайта рестрикции этого фермента, обозначенные нами как сайт 1 (S1) и сайт 2 (S2, рис. 19А). Для исследования статуса метилирования этих сайтов мы использовали праймеры 26-14 и 26-8 (S1), 26-1 и 26-3 (S2); рестрикцию ДНК ферментом SacII проводили двукратно. Результаты амплификации представлены на рисунке 19Б. В двух клеточных линиях при изучении статуса метилирования сайта 1 мы наблюдали присутствие продукта ПЦР, а при исследовании статуса метилирования сайта 2 продукт ПЦР отсутствовал в тех же образцах ДНК.
Однако, при исследовании статуса метилирования сайта 1 рестриктазы SacII в образцах нормы и опухоли шейки матки (амплификацию проводили с применением праймеров 26-7 и 26-8) выяснилось, что в шести из восьми- 82 -
образцов продукт ПЦР присутствовал не только в опухоли, но и в норме. Пример такого исследования представлен на рисунке 19В. В тоже время в случае сайта 2 продукта ПЦР для большинства этих образцов не было ни в норме, ни в опухоли. Такой результат может быть связан с одной стороны с метилированием этого рестрикционного сайта не только в опухоли, но и в нормальной ткани, а с другой стороны с тем, что данный сайт является труднорестрицируемым.
 |
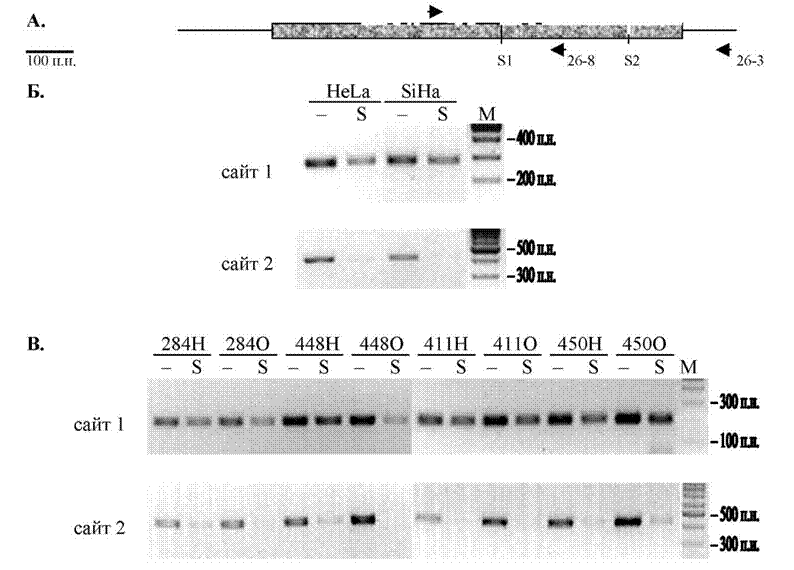 Рисунок 19. Анализ статуса метилирования CpG динуклеотидов, входящих в состав сайтов рестрикции метилчувствительного фермента SacII в пределах CpG островка гена вЗА-адаптина методом МЧ-ПЦР. А. Схематическое изображение расположения сайтов рестрикции фермента SacII (вертикальная черта). Стрелками обозначены использованные праймеры; ? - CpG островок, И - 1 экзон. Б. и В. Результат электрофореза в агарозном геле продуктов амплификации ДНК из клеточных линий (Б) и из образцов нормы и опухоли шейки матки (В). Рестрикция образцов ДНК SacII (S) или без рестрикции (-). М -маркер 100 bp. Числами вверху указаны номера образцов, где Н - ДНК из нормальной ткани, О - ДНК из опухоли шейки матки; числами справа - размеры маркерных фрагментов.
Рисунок 19. Анализ статуса метилирования CpG динуклеотидов, входящих в состав сайтов рестрикции метилчувствительного фермента SacII в пределах CpG островка гена вЗА-адаптина методом МЧ-ПЦР. А. Схематическое изображение расположения сайтов рестрикции фермента SacII (вертикальная черта). Стрелками обозначены использованные праймеры; ? - CpG островок, И - 1 экзон. Б. и В. Результат электрофореза в агарозном геле продуктов амплификации ДНК из клеточных линий (Б) и из образцов нормы и опухоли шейки матки (В). Рестрикция образцов ДНК SacII (S) или без рестрикции (-). М -маркер 100 bp. Числами вверху указаны номера образцов, где Н - ДНК из нормальной ткани, О - ДНК из опухоли шейки матки; числами справа - размеры маркерных фрагментов.
В связи с этим мы использовали бисульфитный сиквенс ДНК, который позволяет точно установить статус метилирования каждого CpG динуклеотида, и не ограничен рамками сайтов рестрикции. При обработке ДНК бисульфитом натрия в молекуле цитозина происходит замещение атома водорода на гидросульфитную группу по углероду в шестом положении. Эта модификация приводит к резкому возрастанию скорости дезаминирования и к превращению после десульфонации остатка цитозина в остаток урацила.
5-метилцитозин таким превращениям не подвергается. Поэтому, после амплификации обработанной ДНК в продукте ПЦР происходит замена неметилированного остатка цитозина на остаток тимина, в то время как метилированный остаток цитозина остается самим собой, т.е. остатком цитозина. Исследование статуса метилирования CpG-островка гена вЗА-адаптина таким способом мы проводили в клеточной линии HeLa и в образцах опухоли 286 и 289 (модифицированная бисульфитом натрия ДНК была любезно предоставлена сотрудником лаборатории молекулярной биологии вирусов Ивановой Т.А.). Амплификацию мы проводили полугнездовым способом с помощью трех праймеров, подобранных к модифицированной ДНК. На рисунке 20 схематически представлена исследуемая зона, а также приведены хроматограммы сиквенса участка CpG-островка длиной 86 н.п. в клетках HeLa и в образце опухоли 289 и немодифицированная нуклеотидная последовательность этого же участка. Видно, что в обоих случаях во всех шести приведенных CpG динуклеотидах остаток цитозина оказался замещенным на тимин, т.е. был неметилированным. Приведенный на рисунке сайт рестриктазы SacII представляет собой первый из двух расположенных в границах CpG-островка сайтов рестрикции этого фермента (сайт S1). По результатам применения МЧ-ПЦР данный сайт имел метилированный статус. Здесь же оба CpG динуклеотида, входящих в состав сайта, находятся в неметилированном состоянии. Таким образом, мы столкнулись с труднорестрицируемым сайтом рестриктазы SacII.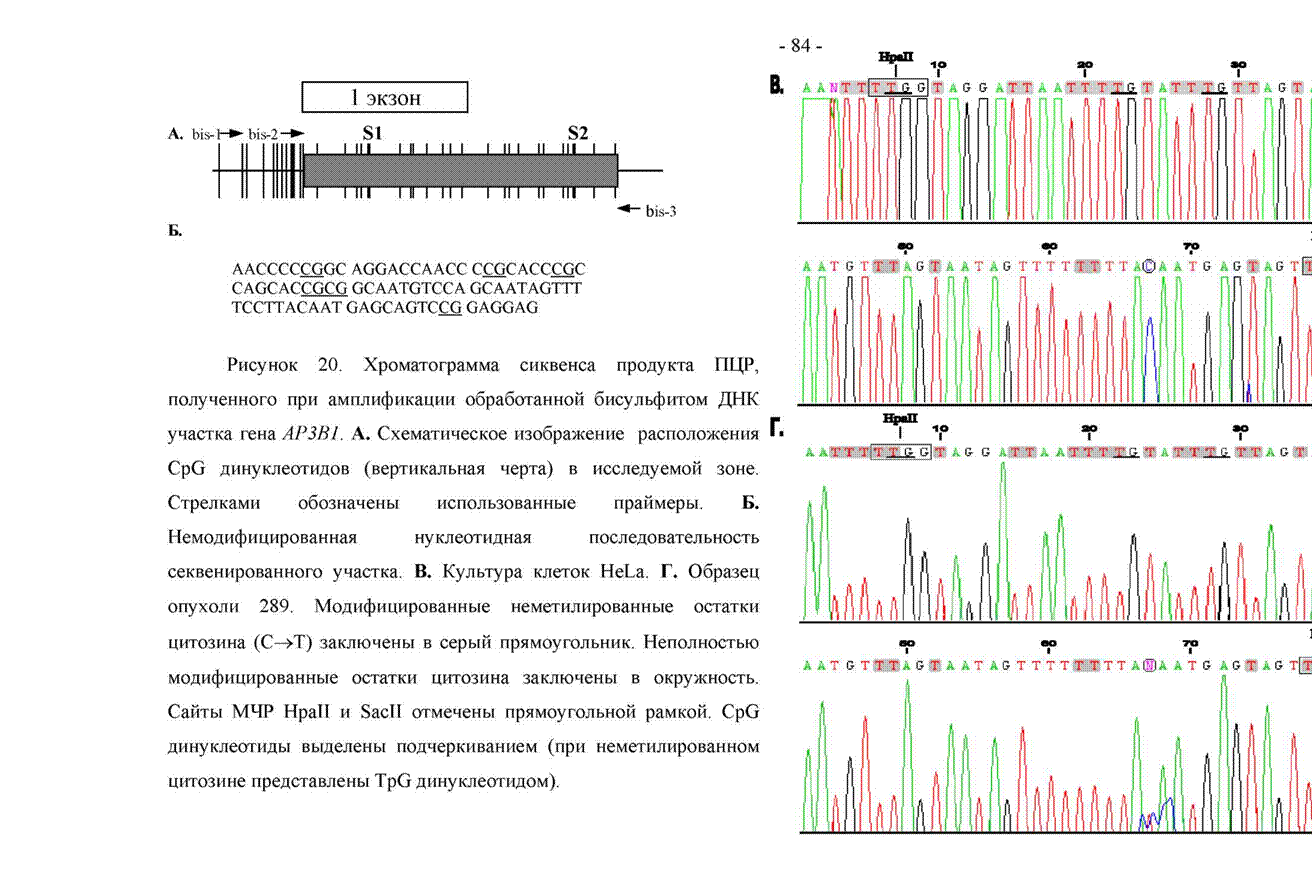
Обобщенные результаты, которые мы получили при изучении статуса метилирования участка CpG-островка гена |З3А-адаптина размером 411 н.п. с помощью бисульфитного сиквенса, выглядят следующим образом. Во всех трех случаях мы исследовали 26 CpG динуклеотидов. В клетках линии HeLa неметилированный статус имели 21 CpG динуклеотид. В образце опухоли 286 неметилированное число CpG динуклеотидов составило 23, в образце 289 неметилированными были 24 CpG динуклеотида.
При этом следует заметить, что, в противоположность неметилированным CpG динуклеотидам, статус метилирования остальных исследованных CpG динуклеотидов однозначно определить было невозможно, так как хроматографическая картина сиквенса этих CpG динуклеотидов и окружающих их районов не имела достаточной четкости и ясности. Исходя из результатов, полученных нами тремя разными методами, можно заключить, что CpG-островок гена |З3А-адаптина не подвергается существенному метилированию в клеточных линиях рака шейки матки и в плоскоклеточных карциномах шейки матки.Таким образом, с одной стороны экспрессию мРНК гена |З3А-адаптина можно реактивировать деметилирующим агентом в клеточных линиях рака шейки матки, а с другой стороны CpG-островок самого гена не подвергается аберрантному метилированию в этих же клеточных линиях. Эти данные позволяют сделать вывод о том, что экспрессия мРНК |З3А-адаптина подавлена в клетках HeLa и SiHa, и в одной из 15 опухолей шейки матки, но это подавление экспрессии не связано с метилированием исследованного района CpG-островка гена |З3А-адаптина.
- 86 -
Еще по теме 2.2. Изучение статуса метилирования CpG-островка гена /ЗА-адаптина в опухолях шейки матки и в клеточных линиях рака шейки матки.:
- 2.1. Исследование экспрессии мРНК гена вЗА-адаптина в опухолях шейки матки и в клеточных линиях рака шейки матки.
- 2. Исследование экспрессии и статуса метилирования гена р3А-адаптина при раке шейки матки.
- 1.З. Определение статуса метилирования CpG-островка З2 при раке шейки матки.
- 1. Поиск CpG-островков, гиперметилированных в опухолях шейки матки
- 1.4. Определение полного размера CpG-островка гена /ЗА-адаптина.
- 225.2. Лечение рака шейки матки IA стадии.
- ЦИТОЛОГИЧЕСКИЙ СКРИНИНГ РАКА ШЕЙКИ МАТКИ У ЖЕНЩИН 35 - 69 ЛЕТ В МОСКВЕ
- Возможности УЗКТ в стадировании рака шейки матки
- ПЕТРЕНКО АНАТОЛИЙ АНАТОЛЬЕВИЧ. Анализ метилирования ДНК при раке шейки матки, 2003
- Частота клинических проявлений начальных форм рака шейки матки
- Сравнительные результаты кольпоскопического и цитологического исследования у больных начальными формами рака шейки матки
- НЕПОСРЕДСТВЕННЫЕ РЕЗУЛЬТАТЫ КОМБИНИРОВАННОГО ЛЕЧЕНИЯ МНОЖЕСТВЕННЫХ МЕТАСТАЗОВ В ЛЕГКИЕ РАКА ШЕЙКИ МАТКИ
- РАК ШЕЙКИ МАТКИ
- Рак шейки матки
- Рак шейки матки