УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
сопрозрачной эктоплазмы, в которую позднее вливается и эндоплазма (рис. 32, А). Слой эктоплазмы вариирует по толщине.
Стекловидная эктоплазма хорошо диференцирована от эндоплазмы даже в покойнохМ состоянии амебы.
На поверхности эктоплазмы иногда образуется довольно плотная пелликула.Движение дизентерийной амебы связано с энергичным цик лозом эндоплазмы, в которой развиваются токи самого разнообразного направления, принимая часто характер вращательных или вихреобразных движений; однако не всегда развитие бурных токов эндоплазмы ведет к передвижению самой амебы (Г. Эпштейн).
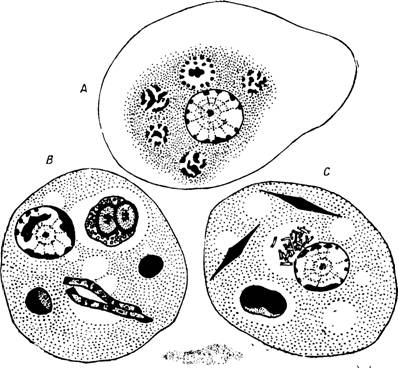
Рис. 31 —Дизентерийная амеба.
А — крупнозернистый распад фагоцитированных эритроцитов;
В — фагоцитоз эритроцита, эозинофила", бактерий; С — фагоцитоз эритроцита, бактерий, кристаллов Шарко — Лейдена.
По Г. Эпштейну. .
При обычном движении где-нибудь на поверхности амебы появляется вырост эктоплазмы, образуя более или менее широкую псевдоподию (лобоподию), которая выпячивается в одном направлении; в нее как бы переливается эндоплазма, благодаря чему амеба передвигается с места на место. Гиалиновые псевдоподии могут образовываться в нескольких участках поверхности амебы, но в своем дальнейшем развитии доминирует одна какая-нибудь псевдоподия, которая в своем движении увлекает эндоплазму; Бывают случаи движения амеб, у которых нет диференцировки на экто- и эндоплазму; такая форма движения бывает у экземпляров, только что выделенных из организма или взятых тотчас из культуры и наблюдаемых при температуре 37°. Наиболее активное движение наблюдается при температуре тела и при pH 6,5 (Е. Павлова); при комнатной температуре свежие экземпляры амеб из фекалий и из культур образуют гиалиновые лобоподии.
Эндоплазма мелкозерниста и часто заключает в себе пищевые вакуоли с содержимым.
Ядро лежит в эндоплазме. В живой Ent. histolytica оно видно хуже, чем у Ent. coli. Диаметр ядра колеблется от 4 до 7—8 у. Как правило, ядро одно, шарообразной формы. Оно одето тон-кой одноконтурной оболочкой, под которой лежит тонкий слой хрома- тиновых зернышек. Почти в самом центре ядра помещается маленькая хроматиновая кариозома, окруженная светлым ободком. Сеть линина расположена в общем радиально. Ядро в целом относится к категории «бедных хроматином (табл. I, рис. 31).
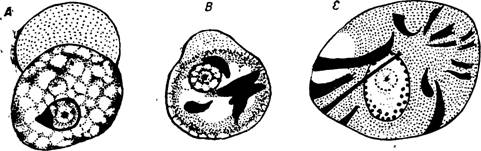
Рис. 32. А — Entamoeba histolytica форма minuta (предцистная стадия); В — она же с хроматоидными включениями; С — хрома- тоидные включения в типичной Entamoeba histolytica.
По Г, Эпштейну.
Дегенеративные процессы, затрагивающие амебу, касаются прежде всего тонкой структуры ее ядра. Периферийный слой хроматина рге- ряет правильность своего расположения и разбивается на неправильные скопления;[4] кариозома отходит ч от центра ядра, увеличивается в .размерах и др.
При специальной гистологической обработке в ядре Ent. histolytica можно найти эксцентрично лежащее зерно, которое, вероятно, является внутриядерным центральным аппаратом. Кроме всех этих образований в ядрах иногда находится по одному эксцентрично лежащему оксифильному ядрышку.
Деление дизентерийных амеб изучено недостаточно. Описываемые разнообразные формы амитотического деления на деле; быть может, являются стадиями дегенерации, а не действительного амитоза.
При кариокинетическом делении из хроматина' ядра формируется шесть хромозом.
Делящиеся амебы в содержимом кишечника встречаются исключительно редко. По Dobell, деление ядра у Е. histolytica является промежуточным по своему типу между настоящим кариокинезом и амитозом.
Ent. histolytica питается, кроме жидкой пищи (эндосмотическое питание — см. ниже), прежде всего эритроцитами, затем лейкоцитами (амеб, заглотивших красные кровяные тельца, называют э р и т р о ф а.- г а м и).
В среднем амеба заглатывает 6—8 эритроцитов, но бывает их и несколько десятков.
Фагоцитированные эритроциты перевариваются, совершенно не изменяясь в своей гомогенной структуре, они как бы «стаивают» с поверхности. Интересно, что в теле дизентерийной амебы не . удается обнаружить следов железа микрохимическими реакциями; одним из объяснений этого является предположение, что гемоглобин в процессе усвоения эритроцитов выделяется из тела амебы вовсе без разложения или последнее протекает в размерах, неуловимых. современными методами исследования (Courtelin).Способность амеб фагоцитировать эритроциты наблюдалась также и в культурах Е. histolytica, что было показано Беком и Доброглавом (Boeck, Dbrohlav, 1925) и другими авторами, (Dobell,. Frey, Meleney и др.). Способность амеб к эритрофагоцитозу связана с особенностями среды. Frey и Меіепеу показали, что свежая лошадиная сыворотка задерживает заглатывание эритроцитов амебами; в фильтратах амебной культуры с лошадиной сывороткой, нагретых до 70° С, эритрофагоцитоз наиболее энергичен; в чистом растворе Рингера он, наоборот, наиболее слаб. Наиболее выраженный фагоцитоз эритроцитов дизентерийной амебой наблюдается при pH от 5,6 до 6,5; сверх pH 7,8 этот процесс выпадает (Е. Павлова).
В эти же пределы укладываются данные Е. Хейсина.
Бактерии Ent. histolytica заглатывает в виде исключения и то, вероятно, в состоянии дегенерации. Заглоченные эритроциты в живой амебе просвечивают в виде округлых желтоватых телец; диаметр их по мере переваривания уменьшается (рис. 31—В; ЗО, 1—2). Потребляются также и другие клетки тела и кристаллы Шарко-Лейдена (рис. 31, С). Не редкость, когда в эндоплазме амебы совсем нет следов оформленных пищевых частиц.
Включения указанных выше сортов являются экзогенными, так как они поступают в амебу извне. Кроме них в протоплазме Ent histolytica бывают включения, формирующиеся в результате жизнедеятельности амебы; таковыми — эндогенными — включениями являются, например, окрашивающиеся ядерными красками хроматоидные тела в форме различных по величине зерен, глыбок, трабекул и др.
(рис. 32, С).2) Ent. histolytica forma minuta живет в просвете, в содержимом толстой кишки. Размеры амеб колеблются от 7 до 15 f* и несколько выше. Мельчайшие minuta описаны были в качестве особого вида — Ent. hartmanni Prowazek. Эктоплазма развита слабее, чем у Ent. histolytica forma magna, и скопляется главным образом в ложноножках (рис. 32 — А, В). Подвижность вариирует в зависимости от подготовленности амеб к инцистированию; во всяком случае, она меньше, чем у вегетативной формы. Перед инцистированием minuta вовсе теряет подвижность. Эндоплазма ее бывает сильно вакуолизована; предцистные амебы не заглатывают ни эритроцитов, ни каких-либо клеток; питание их комменсальное. В вакуолях иногда видны бактерии, которые, пови- димому, не являются главной пищей minuta. Перед самым инцистированием в эндоплазме появляются и увеличиваются в количестве хроматоидные или сидерофильные (т. е. окрашивающиеся железным гематоксилином) тельца (рис. 32, В), а также образуется гликогеновая вакуоля (проба иодом).
Ядро предцистной стадии в общем сходно с формой Ent. histolytica forma magna, хроматин у первой скопляется неравномерно под оболочкой ядра в более крупные массы. Предцистные формы бывают трудно отличимы или вовсе не отличимы от соответствующих форм Ent. coli. 'Мелкие расы minuta могут быть легко смешаны с Endolimax папа.
Значение встречающихся в подобных случаях амеб может быть выяснено многократными исследованиями одного и того же больного в течение хода болезни, что дает возможность проследить смену одной формы дизентерийной амебы другой ее формой и распознать типичные цисты.
Мелкие, просветные, предцистные формы дизентерийной амебы в толще тканей не живут и поэтому обнаруживаются в полужидких испражнениях; их появление совпадает с ослаблением острого течения болезни и началом выздоровления. В связи с таким переходным периодам, одновременно с предцистными амебами обычно встречаются вегетативные формы и даже цисты. Замена вегетативных форм предцистными происходит энергично при лечении амебной дизентерии эметином и др.
Рецидивы амебиаза сопряжены с повторным преобладанием крупных, паразитических форм над мелкими, просветными, комменсальными.3) Цисты Ent. histolytica круглые или слегка овоидные (рис. 30,
7— 11). Диаметр их колеблется в пределах в общем от 7 до 15,4 \ь. Величина цисты зависит от среды и состояния, при котором они измеряются. Истинные размеры получаются при измерениях в физиологическом растворе; почти такой же индиферентной в этом отношении средой является люголевский раствор. Цисты, фиксированные и заключенные в канадский бальзам, являются меньшими по своему размеру процентов на 10 сравнительно с нормой.
Двухконтурная оболочка цисты тонка и бесцветна. Толщина ее в цистах среднего размера достигает 0,5 р. В неокрашенных цистах ядра трудно различимы или вовсе не видны. Строение цисты зависит от степени ее зрелости. В извержениях обычно встречаются все стадии созревания цист. Молодые формы одноядерны (рис. 30, 7—9\ табл. III, рис. 5, 8), затем ядро делится надвое (двуядерная циста) (табл. III, рис. 6). Далее следует новое деление с образованием четырехъядерных цист, каковые и являются типичными для histolytica (рис. 30, 10; табл. III, рис. 7, 9).
Строение ядра у двуядерных и четырехъядерных цист обычно характеризуется «серповидным' скоплением» хроматиновых зерен под оболочкой ядра, каковая форма ядра типична для Е. minuta.
Кроме ядер, в цистах есть так называемые хроматоидные тела (рис. 30, 8; табл. III, рис. 2), имеющие вид палочек или столбиков с округлыми концами, окрашивающиеся железным гематоксилином; в виде сильно преломляющих свет телец, они заметны, и в неокрашенных цистах. В долго лежащих цистах они исчезают. Независимо от этого отмечают, что хроматоидные тела гораздо чаще встречаются у одно- и двуядерных цист, чем у четырехъядерных. Вторым включением в цистах Ent. histolytica является гликогеновая вакуоля (становится видной при действии люголевского раствора), имеющая ббльшие размеры в одноядерных цистах (рис. 30, 5).
Со временем гликоген потребляется инцистированной амебой, и циста лишается вакуоли.
Иногда ядра лежат парами у противоположных точек поверхности цисты. Цисты встречаются в оформленном (твердом) стуле носителей или в периоде внешне, здорового состояния болевшего.Встречаются различные виды атипичных цист; таковыми (Г. Эпштейн) являются цисты с перетяжками или неправильно контуриро- ванные, с несоразмерно крупными ядрами, с ядрами, различными по своим размерам, уродливыми по форме, иногда вовсе безъядерные и др. Дегенеративные цисты встречаются на всех стадиях зрелости.
Вышедшие с фекалиями наружу цисты продолжают созревать, о чем можно судить по увеличению количества четырехъядерных цист. В стуле пятичасовой давности Hegner наблюдал 82°/о одноядерных цист, 9°/о двуядерных и 9% четырехъядерных. Через 22 часа после дефекации в том же стуле оказалось 12% четырехъядерных цист и 29% двуядерных.
Ent. histolytica образует, по мнению ряда авторов, несколько рас, различаемых по средним размерам. В общем диаметр цист колеблется от 5 до 20 р-; отдельные расы также дают известные колебания, например, 7—9, 7—11, 10—14, И—15.
12—18 р* (Dobell и Jepps) (рис. 33).
Вопрос о расах дизентерийной амебы имеет существенное значение, ибо фигурирует и диаметрально противоположное мнение о наличии непатогенных мелких амеб с четырехъядерными цистами, которые при современной технике исследования морфологически неотличимы от цист дизентерийной амебы. В качестве

Рис. 33 —^ 1 у 2,3 — A, By Су D цисты четырех различных фас Entamoeba histolytica. По Dobell и Jepps.
основного довода унитаристы приводят данные Кливлэнда и Сандерса, получавших культуры от посеянных поодиночке цист (изменчивость в «чистых линиях» ИЛИ Ц клонах) и наблюдавших колебания пределов цист от 7 до А23 р., т. е. такую изменчивость, которая покрывает пределы вариаций цист, взятых в естественной обстановке. Отсюда делается вывод, что изменчивость цист дизентерийной амебы носит не расовый, а индивидуальный характер; и если нельзя говорить о наличии рас v Ent. histolytica, то тем менее оснований допускать существование видов непатогенных амеб с четырехъядерными цистами, характеризующимися теми или другими средними величинами.
Однако изменчивость в чистых линиях амеб, определяемая на культуральном материале, т. е. развивавшихся в искусственной среде, не может быть принята (без специальных доказательств) равнозначной вариациям амеб, обитающих в нормальном биотопе, которым является кишечник хозяина. Это положение дает право в ином свете рассматривать результаты биометрического изучения вариаций четырехъядерных цист (морфологически относимых оптом к Ent. histolytica), указывающих на существование групп особей, изменчивость которых колеблется вокруг различных средних величин.
Конечно, и одна биометрика, взятая сама по себе, не является еще ключом к практическому распознаванию различных видов амеб, которых одни соединяют в общий вид Ent. histolytica, а другие описывают в качестве самостоятельных видов Ent. tenufc, Ent. minutissima, Ent. dispar и др.; но в качестве опорного пункта для дальнейших исследований данные биометрики имеют бесспорное значение, особенно при сочетании с другими методами исследования и с результатами эпидемиологических обследований (см. ниже).
t
Инцистирование амеб происходит в просвете кишки; в толще тканей цисты никогда не образуются.
Оболочка цисты стойка к желудочному соку: растворяется она при действии трипсина; благодаря этому проглоченные с загрязненной пищей и водой цисты дизентерийной амебы вскрываются лишь в тонких кишках.
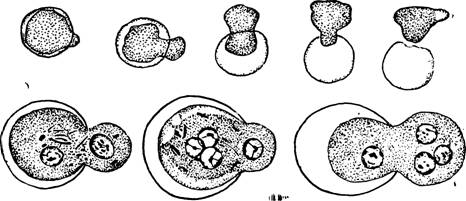
Рис. 34 — Эксцистирование Entamoeba histolytica.
Перед эксцистированием содержимое цисты начинает двигаться; в стенке цисты образуется маленькое отверстие, через которое амеба выходит наружу (рис. 34); далее быстро следует деление ядер, после чего восьмиядерная материнская амеба делится на восемь маленьких дочерних амеб; амебы переходят в толстые кишки, где и обосновываются. Повидимому, возможны случаи эксцистирования дизентерийных амеб в кишечнике того же самого хозяина.
Амебы могут инцистироваться и в культуре; стимулом являются присутствие в среде некоторых бактерий (например, В. mesentericus) или особенности состава питательной среды (например, прибавление рисовой муки). В культуре может происходить непрерывная смена процессов инцистирования и эксцистирования.
Инкубационный период при опытах Walker и Sellard на людях с дизентерийными амебами колебался от 20 до 95 дней, тогда как цисты были обнаружены в испражнениях дней через девять.
Стойкость цист к воздействиям внешних факторов и различных химикалий различна. Высушивание на воздухе их убивает очень быстро; во влажном кале при комнатной температуре цисты выживают около месяца; в разведенных водой faeces цисты можно обнаружить до шестой недели; в то же время хорошо отмытые цисты переживали в дестиллированной воде при температуре 12—22° С до семи месяцев. Температуру в 68° С в воде цисты выдерживают пять минут, а температуру ледника — свыше восьми дней.
.Этот срок в опытах Tsuchiga с промытыми цистами удлинялся до 35 дней, тогда как при комнатной температуре тот же автор наблюдал переживания непромытых цист в течение 3—6 дней, а промытых — в течение 7—9 дней; в фекалиях цисты жили всего 9—11 дней. Такие различия в результатах опытов зависят, видимо, как от вариаций свойств различных штаммов амеб, так и от сочетания влияния полностью неучитываемых факторов внешней среды.
По данным, добытым в Ленинграде, цисты в faeces при 37° С при доступе кислорода не живут больше суток; в тех же условиях при 13—17° С они выдерживают не менее 2 недель, а при 0° сохраняются больше месяца; при переменных температурах (средняя — ниже 0°,’ а минимальная —21° С) цисты выживают до 100 дней.
Некоторые дезинфицирующие вещества оказывают сильное действие на цисты. Крезол в разведении 1 : 250 убивает их в течение 5—15 минут, сулема 1 : 1000 — через 4 часа, 5°/о формалин не влияет на них в течение 5 дней. Цисты весьма чувствительны к действию слабых раз- ведений эметина (даже до 1 : 5 000 000).
Заключение о переживании или гибели цист производится на основании применения эозиновой пробы; если цисты окрашиваются эозином, значит, они погибли; неокрашивающиеся считаются живыми. Однако такое допущение может не всегда отражать действительное положение дела. Поэтому выживаемость была проверяема посредством посева бывших под опытом цист на искусственные питательные среды. Эти опыты дали результаты, в принципе совпадающие с эозиновой пробой. Следует отметить необходимость проверки сохранения патогенности амеб, развивающихся из цист, подвергавшихся различным внешним воздействиям; таковые воздействия могут затронуть также и патогенные свойства амеб при сохранении способности содержимого обработанных цист к размножению.
4) Дегенерирующие формы Ent. histolytica. В очень старых случаях амебной'дизентерии, а также у больных, подвергавшихся лечению,, наблюдаются1 резко дегенерирующие формы Ent. histolytica (табл. I, рис. 6, 7). Ядро у таких амеб теряет круглую форму: хроматин выходит из него и рассыпается глыбками в протоплазме (карио- рексис); в других амебах на первый план выступают в ядрах явления пикноза. В конце-концов ядро как таковое исчезает (рис. 35), а на поверхности амебы образуются почкообразные выросты, ошибочно принятые Шаудином за цисты.
Дегенерирующие формы амеб часто наблюдаются одновременно с нормальными предцистными амебами; очевидно, что в таких случаях дегенерация началась еще во время пребывания амеб в кишечнике.
Дегенеративные процессы в амебах начинаются также вскоре после выхода их из кишечника. Так как для диагностических целей приходится брать материал из faeces, то для исследования попадаются не нормальные амебы, а уже подвергнувшиеся той или другой степени дегенерации, которая прежде всего отражается на тонком строении ядра. Некоторыми исследователями такие дегенерирующие амебы были описаны в качестве новых видов (например, Ent. tetragena Viereck, Е. africana Hartm. и др.).
Дегенеративные изменения могут носить у амеб временный характер, и от них амебы могут оправляться. Например, при действии радия на культуры дизентерийной амебы наблюдаются изменения ядер в форме распада кариозомы, отхождения периферических зерен оболочки •ядра, вытягивания его, отделение безъядерных участков протоплазмы
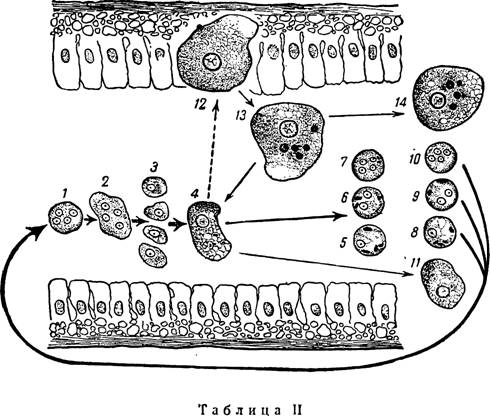
Схема жизненного цикла дизентерийной амебы (Entamoeba histolytia) в свете учения об адаптивных модификациях. Оригинал
Г нездилова
(/) Заглоченная 4-ядерная циста в кишечнике хозяина эксцистируется и дает (2—3) метацистические стадии (по Dobell, Cleveland a. Sanders и др.). Последние превращаются в обычные, вегетативные мелкие, просветно-комменсальные формы (4) —Е• histolytica forma minuta, которые являются основным звеном в жиз“ ненном цикле амебы; они образуют 1—2—4-ядерные цисты (5—6—7). Выделяясь с фекалиями во внешнюю среду, цисты (8—9—10) вновь попадают в организм хозяина, и цикл замыкается. Комменсализмом дизентерийной амебы объясняется обычное исторически сложившееся взаимоотношение ее с организмом хозяина, выражающееся в бессимптомном носителъстве паразита Под воздействием разнообразных факторов в организме хозяина, как в среде обитания паразита комменсала, может возникнуть комплекс услбвий, при котором мелкая, просветная форма (f- minuta) \4) реализует эволюционно вира* ботавшуюся у нее способность к образованию адаптивных модификаций в виде крупных, тканевых, патогенных эритроцито-фагоцитирующих форм (f. magna) (72), поселяющихся в тканях хозяина. Устанавливается новое соотношение с организмом хозяина, выражающееся в заболевали его амебной дизентерией. Крупные, тканевые формы (12), выпадая в просвет кишечника (75), выделяются с фекалиями во внешнюю среду (14) и гибнут, как и мелкие формы (7/), или же они (75) превращаются в исходные мелкие, просветные формы (4), которые образуют цисты (5—6—7)
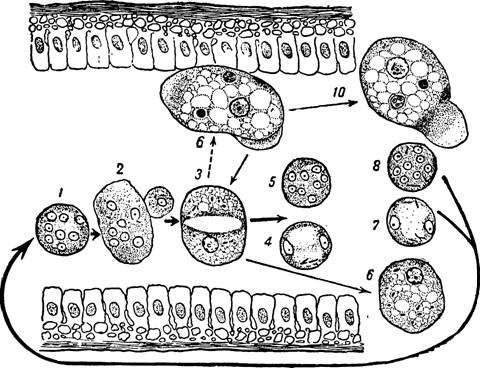
Таблица ПА
Схема жизненного цикла Entamoeba coli в свете учения об адаптивных модификациях. Оригинал Гнездилова
(/) Заглоченная 8-ядерная циста в кишеччике хозяина эксцистируется и дает метацистические стадии (2) (по Dobell), которые превращаются в обычные, вегетативные, комменсальные формы (3 , являющиеся основным звеном в жизненном цикле амебы и образующие 2—8-ядерные цисты (4—5). Последние (7—8), попадая во внешнюю среду, вновь заглатываются хозяином, и цикл замыкается. При определенных изменениях, возникающих в организме хозяина (бациллярная дизентерия, рак, полип кишечника и др.)» обычные, вегетативные Е. соіі (3) могут реагировать на возникшие изменения в среде их обитания образованием адаптивных модификаций—в виде крупных, фагоцитирующих эритроциты и лейкоциты форм (Р) (Е. coll forma magna). Последние описаны в лите >ятуре, как новый вид амеб Соипсіітаппіа lafleuri. Выделяясь со стулом больного во внешнюю среду, они гибнут (10) или же превращаются в исходные формы (3), которые и образуют цисты.
и др.; после прекращения опыта амебы возвращались в прежнее нормальное состояние (Нессет и Кофоид, 1928).
Цикл развития. Хотя изучению паразитических амеб посвящена огромная литература, все же некоторые вопросы соотношения форм Ent. histolytica и посейчас в глазах многих авторов остаются спорными, тем более что полностью жизненный цикл этих амеб еще не прослежен.
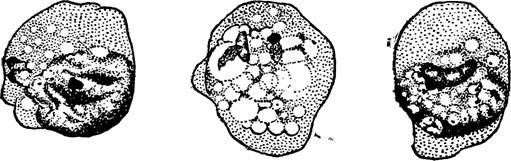
Рис. 35 — Дегенерирующие дизентерийные амебы с разными стадиями разрушения ядра. По Г. Эпштейну.
Заражение человека происходит проглатыванием цист; по взгляду одних авторов, выходящие из цист амебы размножаются делением, оставаясь просветными, комменсальными по питанию формами Ent. histolytica forma minuta. ‘После ряда поколений образуются предцистные формы, инцистирующиеся здесь же в толстой кишке. Цисты могут выделяться годами человеком без каких-либо симптомов кишечных заболеваний. При даче слабительного с жидким стулом выходит много Ent. histolytica forma minuta и предцистных форм.
При наличии особых условий Ent. histolytica forma minuta увеличивается в размерах, внедряется в стенку кишки и превращается в патогенную Ent. histolytica forma magna, которая размножается в инвазиро- ванных ею тканях (табл. II). Эти поглощающие эритроциты формы при недостатке питания делятся на более мелкие амебы и превращаются таким образом в форму minuta и далее инцистируются. С такой точки зрения Ent. histolytica forma magna не является непременным звеном Е жизненном цикле амеб формы minuta. По другим взглядам, Ent. histolytica обусловливает первую фазу инвазии: далее она превращается в Ent. minuta, которая долго живет в кишке, где инцистируется или снова превращается в форму histolytica, что сопряжено с появлением рецидива амебной дизентерии. Первый взгляд автору кажется более приемлемым.
Еще по теме УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ:
- ЗАКЛЮЧЕНИЕ
- Трипаносомы (Trypanosoma)
- ЛАБОРАТОРНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ ПАРАЗИТИЧЕСКИХ ЧЕРВЕЙ
- ПОСВЯЩАЕТСЯ
- ОБЪЕМ И СОДЕРЖАНИЕ ПАРАЗИТОЛОГИИ
- ОРГАНИЗМ КАК СРЕДА ОБИТАНИЯ
- ПОНЯТИЕ О ТРАНСМИССИВНЫХ БОЛЕЗНЯХ
- УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ (медицинская протозоология)
- УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
- УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
- УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
- ОГЛАВЛЕНИЕ
- ФОРМЫ МИАЗОВ У ЧЕЛОВЕКА
- Приложение IVНЕКОТОРЫЕ ДАННЫЕ ПО ЗАМЕНИТЕЛЯМ ИНСЕКТИЦИДОВ
- ОСНОВНЫЕ МЕТОДЫ ОБНАРУЖЕНИЯ И ИССЛЕДОВАНИЯ ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ КИШЕЧНИКА
- БАЛАНТИДИЙ — BA LA NTIDIUM COLI
- Раздел Vl МИКРОСКОПИЧЕСКАЯ ТЕХНИКА
- ИСТОРИЯ СТАНОВЛЕНИЯ И РАЗВИТИЯ ПАРАЗИТОЛОГИИ
- ТЕРМИНОЛОГИЧЕСКИЙ СЛОВАРЬ
- Общие вопросы учения об этиологии болезней