СТАДИЯ АКТИВНОСТИ ОСОБЕННОСТИ ПОВЕДЕНИЯ
Окончание стадии послелиночного доразвития проявляется в приобретении клещами способности к нападению на хозяев или, как часто говорят, в возникновении у них агрессивности. На предшествующей стадии, как уже было отмечено, клещи обычно не реагируют на хозяев и даже в условиях эксперимента прикрепление и начало кровососания у подобных особей можно было наблюдать только в единичных случаях и после нескольких дней нахождения на теле позвоночного.
В связи со значительным биологическим разнообразием группы Іхо- doidea у клещей различают три основных типа нападения на позвоночных животных.
У подавляющего большинства аргасовых клещей и гнездовоноровых видов иксодид нападение осуществляется в замкнутом пространстве норы, пещеры или других сходных местообитаний. По наблюдениям над поведением Ornithodoros papillipes (Павловский, 1960; Поспелова- Штром, 1953; Петрищева и Жмаева, 1962), клещи наползают на хозяина, ориентируясь с помощью высокоразвитых органов обоняния (хеморецепторы пальп и органов Галлера). В частности, в последнее время установлено, что аргасовые (О. coriaceus) и иксодовые клещи (Dermacentor оссі- dentalis и Ixodes pacificus) возбуждаются и движутся в направлении источника углекислоты (Garcia, 1962). Не исключено также участие термо- и механорецепторов, реагирующих на тепловой градиент воздуха и сотрясения субстрата, хотя специальные исследования в этом направлении не проводились. Фоторецепция не может обеспечивать ориентации в условиях темноты нор и пещер, чем можно объяснить полную редукцию глаз у аргасовых клещей и в пределах подсем. Ixodinae. В то же время у клещей сохраняется общая световая чувствительность, проявляющаяся у Ornithodoros papillipes в реакции отрицательного фототаксиса (Павловский, 1950; Неуймин, 1954). Соответственно с характером нападения способности норовых видов к активным перемещениям весьма ограничены и распространение их осуществляется пассивно, на теле хозяев во время миграции последних. Тем не менее в отдельных случаях, ориентируясь на запах хозяев, клещи могут выходить в ночное время из норы и нападать на животных и человека, находящихся вблизи от входа в нору (Петрищева и Шмаева, 1962).Большинство видов иксодовых клещей с пастбищным типом паразитизма пассивно ожидает встречи с подвижными хозяевами, располагаясь в определенных ярусах растительности. Наиболее детально подстерегающий тип нападения изучен у клещей Ixodes persulcatus и /. ricinus (Миронов, 1939; Померанцев и Сердюкова, 1948; Хейсин, 1953; Бабенко и Хижинский, 1964), личинок Boophilus microplus (Wilkinson, 1953) и имаго Dermacentor pictus (Олсуфьев, 1953). Готовые к нападению имаго Ixodes persulcatus поднимаются на травянистую растительность (особенно стебли злаков) или кусты. Наибольшее количество клещей сидит на высоте 25—50 см от поверхности почвы и лишь немногие поднимаются до 1 м. Клещи размещаются вблизи концов стеблей и ветвей в вертикальном положении, а на листьях — параллельно их поверхности с нижней или верхней стороны. При приближении хозяина клещ принимает характерную позу «активного ожидания». Передняя пара ног, наиболее богато снабженная органами чувств, при этом выставлена вперед, вторая и третья пары обхватывают стебель снизу, а четвертая пара отведена назад и цепляется за растение сверху (рис. 6). При отсутствии раздражителей клещи находятся в «пассивной позе» и передние ноги поджаты.
Встреча и прикрепление к хозяину обеспечиваются тремя типами поведенческих реакций (Lees, 1948) 1) реакции ориентации определяют
нахождение клещей в определенных ярусах растительности и занятие ими выгодной позиции для подстерегания; 2) реакции непосредственного реагирования на приближение хозяина, связанные с принятием «активной позы»; 3) реакции, связанные с прицеплением к хозяину и выбором на его теле места для присасывания. Приближение хозяина определяется по раздражению механорецепторов (неожиданные колебания растительности), рецепторов общей световой чувствительности (внезапное затенение), а главное по запаху и теплу, исходящим от хозяина.
При длительном воздействии двух последних раздражителей с расстояния в несколько метров возможно и активное переползание клещей в направлении хозяина.Радиус активных горизонтальных перемещений клещей с подстерегающим типом нападения, по данным наших опытов с Ixodes persulcatus и Rhipicephalus turanicus (Балашов, 1958а, 19596) и наблюдений над поведением Ixodes ricinus (Lees a. Milne, 1951), оказался весьма ограниченным.
Для выяснения подвижности имаго I. persulcatus в кедрово-широколиственных лесах Южного Приморья отлавливались партии голодных самцов и самок, метились быстро сохнущей водонесмываемой белой краской и выпускались в определенных пунктах тех же стаций. Самки выпускались в подстилку партиями по, 500 особей на расстоянии 10, 25 и 50 мот тропы, по которой постоянно ходили люди. В общей сложности было выпущено 7500 клещей в сроки между 1 и 10 июня 1956 г. Через месяц вся территория в радиусе 50 м от мест выпуска тщательно несколько раз облавливалась флажком.
Результаты опытов сведены в табл. 3. Из нее видно, что количество выловленных меченых самок было очень велико и достигало 37.07% от общего числа выпущенных. Меченые клещи через месяц после начала опыта в основной массе оставались в радиусе 5 м от точки выпуска. Плотность их постепенно снижалась от центра к периферии. На расстояние свыше 5 м переместилось всего 106 самок (3.45%). Несколько клещей бьшо поймано в 10—40 м от места выпуска, но остается неясным, прошли ли они это расстояние активно или же были пассивно перенесены каким- либо зверем или птицей.
«Притягивающее» действие тропы проявлялось только на расстоянии до 10 м. В сторону тропы здесь переместилось 560 особей (22.4% от общего числа выпущенных), а от тропы только 222 (8.88%). Еще важнее, что количество клещей, переползших в сторону тропы больше чем на 5 м, составляло 75 особей (3.0%), а от тропы — 3 (0.12 %). Таким образом, в данном случае приходится констатировать направленное, хотя и очень медленное, активное движение клещей в сторону тропы.
Привлекающим фактором в данном случае, вероятно, является запах хозяина. Интересно, что в опытах Б. И. Померанцева и Г. В. Сердюковой (1948) при выпуске клещей в 5 м от тропы, по визуальной оценке, большая часть их переместилась к самой тропе. Вероятно, чем интенсивнее раздражающий стимул, тем сильнее активное перемещение в его сторону.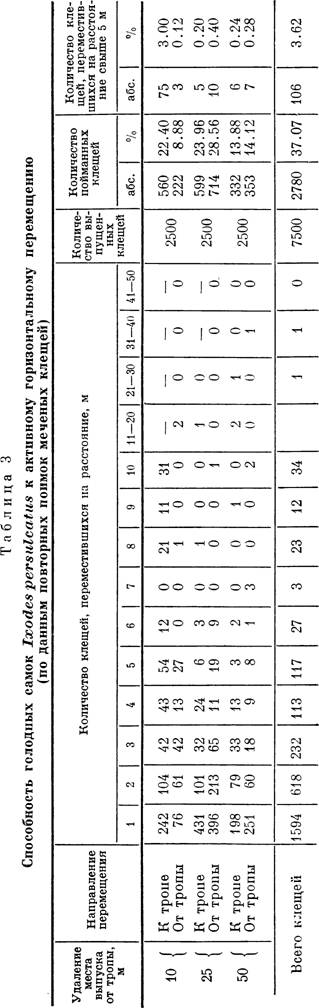
При удалении точки выпуска на 25 и 50 м от тропы картина меняется. Клещи расползаются более или менее равномерно по всем направлениям от центра. Количество особей, переместившихся дальше чем на 10 м, очень невелико.
Сходные данные были получены и для клещей R. turanicus. В фисташковом редколесье Бад- хызского заповедника (Туркменская ССР) были отловлены на флажок и повторно выпущены после мечения сухой жирорастворимой краской 1000 самцов и самок. Отлов и выпуск клещей осуществлялись в типичных для вида биотопах и в сезон их максимальной активности — в апреле месяце 1958 г. Сбор меченых клещей проводился через 5, 10 и 15 суток после выпуска (табл. 4). Возврат клещей был достаточно высок и достигал 50—80%. Основная масса клещей (до 80 %) была поймана в радиусе не бо-Подвижность голодных Rhipicephalus turanicus (по данным повторно пойманных меченых клещей)
| Номер опыт ной пло щадки | Время расползания, в сутках | Пол | Количество клещей, переместившихся на расстояние, м | Всего выпу щено | Всего поймано | |||||||||||
| 0-1 | 1-2 | 2-3 | 3-4 | 4-5 | 5-6 | 6-7 | 7-8 | 8-9 | 9—10 | абс. | 7о | |||||
| Самцы | 55 | 10 | 5 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 130 | 71 | 54.6 | |||
| Самки | 86 | 27 | 6 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 170 | 122 | 71.7 | |||
| 1 | 5 | |||||||||||||||
| Всего | 141 | 37 | 11 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 300 | 193 | 64.3 | |||
| Самцы | 102 | 16 | 5 | 1 | 9 | 2 | 0 | 1 | 3 | 0 | 200 | 139 | 69.5 | |||
| Самки | 121 | 15 | И | 4 | 3 | 9 | 2 | 7 | 1 | 1 | 200 | 174 | 87*0 | |||
| 2 | 10 . | |||||||||||||||
| Всего | 223 | 31 | 16 | 5 | 12 | 11 | 2 | 8 | 4 | 1 | 400 | 313 | 78.3 | |||
| Самцы | 37 | 11 | 2 | 3 | 2 | 0 | 1 | 0 | 0 | 0 | 100 | 56 | 56.0 | |||
| Самки | 59 | 15 | 8 | 6 | 4 | 3 | 1 | 0 | 1 | 2 | 20Э | 99 | 49.5 | |||
| 3 | 15 , | |||||||||||||||
| Всего | 96 | 26 | 10 | 9 | 6 | 3 | 2 | 0 | 1 | 2 | 300 | 155 | 51.7 | |||
лее 2 м от точки выпуска.
Лишь одиночные особи расползались на 5— 10 м. На расстоянии свыше 10 м не было поймано ни одного клеща. Большая часть клещей оставалась вблизи от точки выпуска. Заметные отличия в подвижности самцов и самок отсутствовали.Приведенный фактический материал заставляет с очень большой осторожностью говорить о способности голодных клещей к значительным активным горизонтальным перемещениям. Клещи действительно способны к направленному переползанию, но лишь под воздействием привлекающих химических стимулов. Вероятно, последние связаны со специфическими запахами хозяев. В случае присутствия подобного химического раздражения клещи начинают довольно медленно двигаться в его сторону в зону максимальной интенсивности раздражителя. Радиус восприятия этих раздражений относительно невелик и, вероятно, немною превышает 10 м. Примерно такой же величины достигали в наших опытах и активные горизонтальные перемещения одиночных особей. На небольших расстояниях, порядка 1—2 м от хозяина клещ уже принимает позу активного ожидания, а в ряде случаев может даже активно и довольно быстро двигаться в его направлении. Направленные перемещения клещей в сторону максимальной интенсивности запаха хозяев несомненно следует рассматривать как хорошо выраженную реакцию хемотропизма. При отсутствии соответствующих стимулов расползание клещей в горизонтальной плоскости очень невелико, а его направления беспорядочны.
У некоторых видов иксодовых клещей подстерегающий тип нападения совмещается с активным преследованием обнаруженных хозяев. Подобный тип нападения свойствен имаго Hyalomma asiaticum и, вероятно, другим видам Hyalomma, широко распространенным в пустынных и полупустынных районах Средней Азии и Закавказья. Большую роль в обнару-жении позвоночных и ориентации при их преследовании играют достаточно сложно устроенные глаза. По нашим наблюдениям в закрепленных барханных песках пустыни Кара-Кум, голодные имаго Н. asiaticum обычно неподвижно сидят на поверхности почвы и, реже, на невысокой злаковой растительности, выбирая затененные участки, или совершают беспорядочные перемещения во всех направлениях. При обнаружении крупного позвоночного животного или человека клещи сразу же бегут к нему и могут преследовать на расстоянии до нескольких десятков метров.
Для выяснения подвижности клещей Н. asiaticum мы выпустили в природных биотопах этого вида в сезон активности (апрель 1958 г.) 2500 голодных самцов и самок, меченных порошком сухого красителя Судан III, и затем повторно отлавливали их через 5, 10 и 30 дней. Полученные данные свидетельствуют о весьма значительной способности этого вида к горизонтальным перемещениям (табл. 5). Несколько клещей были пойманы в 500 м от точки выпуска. Расстояние в 100—400 м прошли 10—25% пойманных меченых клещей. Наибольшее количество выпущенных особей оставалось в радиусе 25—100 м от места выпуска, но в непосредственной близости от него (в радиусе до 10 м) осталось лишь несколько клещей. Фактическая подвижность голодных Н. asiaticum может быть значительно выше, так как по мере удаления от точки выпуска шансы поимки всех меченых особей все время уменьшаются, а на расстоянии свыше 200 м полный отлов выпущенных клещей становится нереальным. Косвенно о высокой подвижности свидетельствовали также невысокий процент возврата (12—16%) при низкой смертности. Зависимость пройденного расстояния от времени была особенно заметна в первые дни после выпуска. Это, вероятно, было связано с активными поисками подходящих микростаций,
Таблица 5
Подвижность голодных Нуаіотта asiaticum (по данным повторно пойманных меченых клещей)
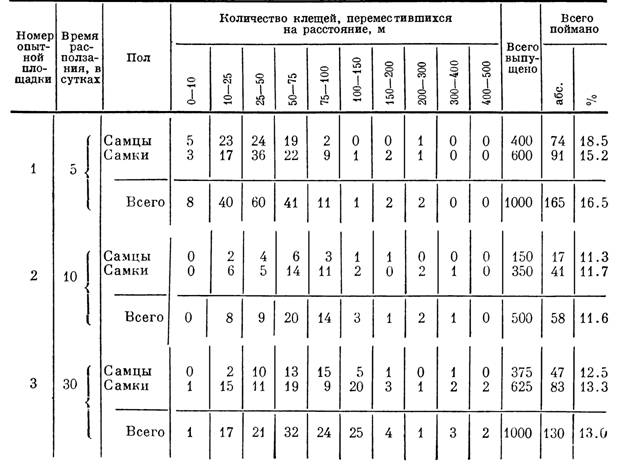
так как пойманные особи всегда концентрировались вблизи кустов саксаула, кандыма и других растений, обеспечивающих некоторое затенение. Заметные различия в подвижности самцов и самок отсутствовали.
В связи с весьма ограниченной способностью к активным перемещениям расселение клещей надсем. Ixodoidea во многих случаях обеспечивается пассивным переносом питающихся особей на теле хозяев. Естественно, что виды с продолжительными сроками питания, паразитирующие на птицах, имеют большие возможности расселения по сравнению с паразитами малоподвижных млекопитающих или видами, не имеющими в цикле развития стадии многодневного кровососания. Этим можно объяснить широкие области распространения некоторых видов рода А г gas или Ornltho- doros capensis (паразиты птиц с многодневным питанием личинок) и более ограниченное распространение некоторых видов рода Ornithodoros с кратковременным питанием личинок (Филиппова, 1966). Виды, паразитирующие на перелетных птицах, в период осеннего и весеннего пролета могут переноситься на многие сотни и тысячи километров от своего ареала. Так, на территории Финляндии были найдены клещи Hyalomma plum- Ъеит plumbeum, а в Болгарии — АтЫуотта hebraeum, занесенные на мигрировавших с юга птицах (Nuorteva a. Hoogstraal, 1963; Pavlov a. Popov, 1951).
В дельте Нила во время весеннего пролета на теле перелетных птиц постоянно обнаруживались личинки и нимфы африканского подвида Hyalomma plumbeum rufipes и, в меньшем количестве, других видов родов Hyalomma, АтЫуотта и Haemaphysails, которые, по мнению Г. Хог- страала с сотрудниками (Hoogstraal a. oth., 1961, 1963, 1964), постоянно транспортируются в Европу и Северную Азию. Напротив, во время осеннего пролета на тех же видах хозяев найдены в большом количестве личинки и нимфы Hyalomma plumbeum plumbeum, Ixodes frontalis, I. riel- nus, Haemaphy sails punctata, H. otophila, H. sulcata, занесенные в Египет, из Европы и Азии.
Поведение голодных клещей в период их активности определяется многими факторами внешней среды и зависит от физиологическою состояния членистоногих. Детальные исследования поведения имаго клещей Ixodes riclnus и I. persulcatus, выполненные в различных частях их ареала и различными методами, установили существование периодических вертикальных миграций из подстилки на растительность и в обратном направлении (Lees a. Milne, 1951; Хейсин, 1953; Лутта и Шульман, 1958; Бабенко и Хижинский, 1964). Подъем клещей на растительность обеспечивается существованием у них отрицательного геотаксиса, но при неблагоприятных условиях температуры и влажности, уменьшающих нормальное содержание воды в организме, у клещей развивается положительный геотаксис и они уходят в подстилку. Важную роль при этом играет и непосредственная реакция на градиент влажности в различных слоях припочвенного воздуха. В связи с этой особенностью поведения период фактического пребывания клещей на растительности значительно короче, чем весь период их активности — от начала приобретения способности к нападению на позвоночных и до окончательного исчезновения. Так, по наблюдениям А. Лиса и А. Мильна (Lees a. Milne, 1951), общая продолжительность индивидуальной активности имаго I. riclnus в Англии составляет в среднем 30 суток, но из них на растительности клещи проводят всего 9 дней, а остальное время находятся в подстилке, совершая четыре вертикальные миграции. В зависимости от климатических особенностей местности продолжительность индивидуальной активности может значительно варьировать и, в частности, в Карелии (Лутта и Шульман, 1958) интервалы между последовательными вертикальными перемещениями, как и общая продолжительность индивидуальной активности, значительно короче.
Еще короче время индивидуальной активности для I. persulcatus, обитающих в районах с континентальным климатом. В Пермской обл. индивидуальная активность имаго продолжалась в среднем — в молодом березовом лесу 7 дней и в старолесье 15 дней (Морозов, 1963), а в Красноярском крае от 1 до 61 дня (Хижинский, 1963), включая и время ухода в подстилку. Соотношение между числом активных клещей в подстилке и на растительности, а также степень «агрессивности» последних определяется погодными условиями и в первую очередь температурой и относительной влажностью воздуха. При этом, учитывая огромные размеры ареалов обоих видов, нельзя ожидать однотипности поведения в различных географических областях. Активность клещей может быть круглосуточной, но максимум ее всегда приходится на светлое время. В Карелии наибольшее число активных самцов и самок /. persulcatus наблюдалось при температуре 5 — 15° и относительной влажности воздуха в 55—80%. Температура выше 20° и относительная влажность ниже 50% вызывают снижение активности и уход клещей в подстилку (Хейсин, 1953). В Красноярском крае (Бабенко и Хижинский, 1964) за сезон наблюдений наибольшее количество активных клещей обнаружено при сочетании температуры в 0—15° и относительной влажности в 60—100% и наименьшее при 16— 30° и той же относительной влажности. Наименьшее количество активных клещей наблюдалось в предутренние и раннеутренние часы, а в остальное время суток активность зависела от погоды: в солнечные дни она была в прямой зависимости от относительной влажности и в обратной от температуры, в пасмурные дни картина была обратной.
Аналогичное чередование периодов собственно активного подстерегания хозяев на поверхности почвы и пребывания в норах грызунов и других сходных микробиотопах было обнаружено и у имаго клещей Нуаіотта asiaticum в пустыне Каракумы (Балашов, 1960а). Наши наблюдения охватывали переходный период окончания весны и начала лета 1959 г. в окрестностях ст. Репетек. В апреле поверхность почвы была покрыта богатой травяной растительностью, выгоревшей в начале мая. Температура воздуха в дневные часы достигала в апреле 25—35°, а в мае повысилась до 30—40°. Незащищенная поверхность почвы нагревалась до 60—70° В утренние и вечерние часы температура воздуха колебалась в пределах 18—30°, а поверхности почвы — 20—35° Относительная влажность воздуха составляла днем 10—20%, а перед рассветом поднималась до 40— 60%.
До выгорания травяной растительности клещи довольно равномерно распределялись по всей описываемой зоне, отсутствуя лишь в разбитых, не закрепленных песках. Особенно часто они встречались вблизи колоний большой песчанки — основного прокормителя их личинок и нимф. По мере повышения средних дневных температур и выгорания растительности наблюдалась закономерная концентрация клещей вокруг деревьев саксаула, крупных кустов ревеня, нор грызунов и других укрытий, создающих затенение от прямых солнечных лучей.
В начале апреля, в относительно нежаркие дни, клещи бегают по песку и активно преследуют человека или животных в течение всего дня. При повышении дневной температуры воздуха до 30° и почвы до 45—50° их активность переместилась на утро с рассвета до 10 час. ина вечер с 6 час. и до наступления темноты. Ночью нападение клещей обычно не наблюдается. Днем они прячутся в различные убежища с менее высокими температурами. Особенно часто Н. asiaticum скрывались в норах большой песчанки.
Наблюдения над поведением клещей в садках (табл. 6) позволили нам точно выяснить влияние окружающей температуры и влажности на их суточную активность.Влияние температуры и влажности на поведение голодных самцов н самок Hyalomma asiaticum в сезон их активности в песчаной пустыне в Туркменской ССР (средние температуры и относительные влажности за апрель—первую половину мая 1959 г.; всего в опытах использовано
600 клещей)
| Время суток, час. | Температура | Относительная влажность воздуха, °/0 | Процент активных клещей на поверхности песка | ||||||
| открытые биотопы | убежища | ||||||||
| воздух на высоте 2 м | припоч- венный слой воздуха | поверх ность почвы | нора песчанки на глубине 50 см от входа | почва под листьями ревеня | растительная подстилка в тени саксаула | на высоте 2 м | припоч- вениый слой | ||
| 5 | 17.0 | 16 | 23 | 16 | 17 | 45 | 3 | ||
| 8 | 18.5 | 19.8 | 23 | 22 | 18 | 18 | 49 | 49 | 59 |
| 9 | — | 25.0 | 36 | 23 | 24 | 22 | — | 31 | 84 |
| 10 | — | 29.0 | 43 | 24 | 26 | 24 | — | 23 | 78 |
| 11 | — | 31.0 | 50 | 24 | 30 | 27 | — | 20 | 4 |
| 14 | 30.9 | 35.2 | 62 | 24 | 37 | 32 | 20 | 16 | 0 |
| 18 | — | 31.0 | 37 | 24 | 25 | 31 | — | 20 | 36 |
| 19 | — | 28.0 | 30 | 24 | 23 | 28 | — | 26 | 81 |
| 20 | 26.8 | 25.5 | 28 | 24 | 21 | 25 | 29 | 29 | 62 |
| 23 | — | 23.0 | 22 | 24 | 19 | 19 | — | 39 | 2 |
Как видно из табл. 6, до восхода солнца клещи неактивны. Температура воздуха в 19—20° является нижним порогом активности, и клещи в основном неподвижно сидели на поверхности почвы или находились в убежищах. С восходом солнца температура воздуха и особенно поверхности почвы быстро поднимается и клещи начинали двигаться по садку. В 8 час. утра при температуре воздуха 20—21° и почвы 23° в активном состоянии находилось 59% клещей. Между 8 и 10 час. они проявляли максимальную активность (до 84% в 9 час.). Температура почвы при этом колеблется от 23 до 43°, а относительная влажность припочвенного слоя воздуха — от 20 до 50 %. С 10 час. при дальнейшем повышении температуры почвы и воздуха клещи начинали прятаться в различные убежища. К 11 час. при температуре почвы около 50° на ее поверхности оставалось только 4% клещей, да и те сидели в полутени. В дневные часы все клещи находились в убежищах. Активность их возобновлялась с 18 час., когда температура почвы опускалась до 35—40° при 20% относительной влажности. С наступлением полной темноты они вновь становились неподвижными, хотя температура почвы и воздуха была еще достаточно высокой.
Дневные убежища клещей отличаются значительным постоянством микроклимата. Особенно стабильны температура и влажность в норе большой песчанки. На глубине 50 см от входа в апреле—мае средняя суточная температура составляла 22—24°. Относительная влажность держится на этой глубине в пределах 84—100% (Ширанович и др., 1959). В подстилке под деревьями саксаула и на почве под листьями ревеня суточные колебания температуры значительно больше, но ее максимальные значения никогда не превышали 40° при нагреве рядом расположенных незащищенных участков почвы до 70°.
Из приведенного материала видно, что основным фактором, ограничивающим суточную активность Н. asiaticum, служит температура. Нижний порог активности этого вида составляет 19—20° и верхний — 40—45°. При нагреве почвы в дневные часы до 70° клещи, спасаясь от перегрева, уходят в различного рода убежища. Температура в последних относительно стабильна и не превышает днем 40°. Изменения относительной влажности воздуха не оказывают заметного влияния на их поведение.Наконец, прекращение нападения клещей в ночные часы, вероятно, можно объяснить ведущей ролью зрительного анализатора в нахождении хозяев у описываемого вида.
Наряду с суточным ритмом активности у Н. asiaticum существует также закономерное чередование более продолжительных периодов преимущественного нахождения на поверхности или в убежищах. В опытах с ме- чением клещей сухим Суданом III удалось установить, что 10—20-дневные периоды активности чередуются у них с 2—4-суточным нахождением в убежищах с высокой относительной влажностью. В наших садках постоянно от 10 до 20% от общего числа клещей по нескольку суток не вылезало на поверхность.
У клещей Dermacentor variabilis, многочисленных в открытых стациях Виргинии (США), активность голодных личинок и имаго находится в прямой зависимости от интенсивности солнечного освещения, но на нее мало влияет температура воздуха. В пасмурную погоду активность клещей сильно снижается даже при высоких температурах (Atwood a. Sonen- shine, 1967).
Еще по теме СТАДИЯ АКТИВНОСТИ ОСОБЕННОСТИ ПОВЕДЕНИЯ:
- Индивидуально-психологические особенности «Реактивного», «Активного», «Проактивного» типов совладающего поведения
- Стадия активности
- 17. Понятие «девиантное поведение». Критерии определения понятия. Особенности отклоняющегося поведения личности.
- Патофизиологическая стадия или стадия функциональных и структурных нарушений.
- 3.4.2 Взаимосвязь «Реактивного», «Активного», «Проактивного» типов совладающего поведения и показателей эффективности/неэффективности процесса совладания
- Типы совладающего поведения: «Реактивное», «Активное», «Проактивное»
- 21.Отличительные особенности уверенного, неуверенного и агрессивного поведения личности.
- 21.Отличительные особенности уверенного, неуверенного и агрессивного поведения личности.
- 6.Возрастные особенности суицидального поведения.
- P.O.R. Особенности инъекционного поведения участников группы
- P.1.R. Особенности сексуального поведения участников группы
- P.4.R. Особенности сексуального поведения участников группы
- P.P.6. Особенности сексуального поведения участников группы
- P.O.6. Особенности сексуального поведения участников группы
- Индивидуально-психологические особенности совладающего поведения
- Особенности мышления и поведения сектанта.
- P.1.4. Особенности инъекционного поведения участников группы
- P.P.R. Особенности инъекционного поведения участников группы