ПРИСПОСОБЛЕНИЯ К УМЕНЬШЕНИЮ ПОТЕРЬ ВОДЫ ОРГАНИЗМОМ
Описанные ранее закономерные чередования периодов активного подстерегания хозяев в биотопах с повышенным дефицитом насыщения и нахождения в растительной подстилке, норах, почве и других микробиотопах с достаточно высокой относительной влажностью воздуха определяются необходимостью поддержания в организме определенного уровня содержания воды.
Наземные членистоногие приспособились к жизни в условиях низкой относительной влажности как за счет различных анатомо-физиологических приспособлений, уменьшающих испарение (высокая водонепроницаемость покровов, трахейная дыхательная система, образование малорастворимых конечных продуктов азотистого обмена), так и путем выработки комплекса поведенческих реакций, позволяющих избегать неблагоприятных условий (Гиляров, 1959). Все эти приспособления свойственны и клещам надсем. Ixodoidea, сталкивающимся с опасностью высыхания в период подстерегания хозяев. Устойчивость к высыханию связана с экологическими особенностями вида. Среди аргасовых и иксо- довых клещей удалось обнаружить несколько групп видов, значительно различающихся по способности переживания при высоком дефиците насыщения в лабораторных условиях (Балашов, 1960а; Балашов и Филиппова, 1964; Lees, 1946а, 1947), причем эта способность находится в прямой зависимости от степени увлажненности их природных биотопов.В наших опытах при содержании имаго семи видов аргасовых клещей при 20, 40, 60 и 80% относительной влажности и температуре 18 и 28° установлены значительные различия в среднесуточных потерях веса за счет испарения (табл. 7). Наибольшей засухоустойчивостью отличались Alveonasus lahorensis, Argas persicus и Ornithodoros papillipes. Клещи A. vulgaris, Ornithodoros tartakovskyi и О. capensis оказались более гигрофильными и теряли в весе в среднем в 1.5—2 раза больше, чем виды первой группы, а О. verrucosus занимал в этом отношении промежуточное положение.
Устойчивость к высыханию в значительной степени определялась активными биологическими процессами в организме, а не только механическими защитными приспособлениями. Живые клещи всегда теряли воды в 2—3 раза меньше, чем убитые особи тех же видов, несмотря на сохранение у последних целостности покровов и исключение потерь сквозь стиг- мальные отверстия трахейной системы. В целом всем изученным видам аргасовых клещей была свойственна значительная устойчивость к высыха-Среднесуточные потери веса живыми клещами (в мг/г/сутки) при различной температуре (в °С) и относительной влажности (в %)| Вид клеща | 18° | to oc о | ||||||
| 20% | 40% | 60% | 80% | 20% | 40% | 60% | 80% | |
| Argas persicus | 5.1 | 4.4 | 2.9 | 0.8 | 5.3 | 5.2 | 3.2 | 1.4 |
| А. vulgaris | 6.4 | 5.9 | 5.6 | 3.2 | 10.6 | 9.8 | 5.8 | 4.5 |
| Alveonasus lahorensis | 2.8 | 2.1 | 2.4 | 1.2 | 4.8 | 3.2 | 3.3 | 1.7 |
| Ornithodoros papillipes | 4.7 | 4.7 | 4.1 | 2.5 | 6.2 | 3.0 | 2.8 | 2.5 |
| O. verrucosus . | 7.4 | 5.7 | 3.5 | 2.8 | 6.8 | 5.8 | 5.2 | 0.8 |
| 0. tartakovskyi | 11.0 | 9.7 | 4.8 | 1.5 | 11.6 | 10.9 | 5.9 | 1.6 |
| O. capensis | 8.6 | 7.5 | 6.6 | 0 | 10.6 | 9.1 | 8.6 | 3.5 |
нию, несомненно связанная с существованием в аридных ландшафтах. В условиях низкой относительной влажности клещи могут терять без видимых повреждений в организме до 20% содержания воды. При дальнейшем высыхании начиналась массовая гибель, хотя одиночные особи выдерживали 30% потери в весе.
Все изученные виды длительное время переживали при относительной влажности 60%, которая, вероятно, служит экологическим порогом, выше которого величина испарения уже относительно невелика и не представляет угрозы для существования организма. Относительная влажность 80% и выше представляет уже физиологический порог, за которым водный баланс относительно постоянен и изменения веса связаны преимущественно с метаболическими потерями.
При содержании нескольких видов иксодовых клещей в сухой атмосфере при 25° (Lees, 1946а) их устойчивость к высыханию оказалась в целом ниже, чем у аргасовых клещей. Наименее устойчивым был Ixodes ricinus, терявший в этих условиях до 50% веса за суткп и выживавший не более 1—2 дней. I. canisuga и I. hexagonus выжпвалп 4—5 дней и АтЫуотта cajennense —10 дней при 5% потерях веса за сутки. Наибольшая устойчивость была свойственна клещам Rhipicephalus sanguineus и особенно Dermacentor andersoni и D.
marginatus, которые теряли за сутки всего 2—3% исходного веса и выживали 17—27 дней.Среди иксодовых клещей наиболее приспособленным к существованию при низкой относительной влажности оказался Hyalomma asiaticum (Балашов, 1960а). Голодные имаго Н. asiaticum могут довольно безболезненно терять до 30% воды, содержащейся в их организме (примерно 20% от веса клеща). При дальнейшей потере воды в организме наступают необратимые изменения, скорость испарения резко возрастает и клещ очень быстро погибает. К моменту гибели обычно теряется до 50% воды (30—35?^ веса клеща).
В первой серии опытов (рис. 159, Л), проведенных при температуре, близкой к верхнему порогу активности (36°) при 10% относительной влажности, клещи погибали через 10—15 дней после начала опыта, при 25% — через 15—20 и при 50 % — через 20—25 суток. При 75 и 95 % относительной влажности они выживают по нескольку месяцев, а за 20 дней опыта их вес уменьшился соответственно на 11 и 4%.
При 26° скорость испарения воды в тех же условиях относительной влажности значительно уменьшается (рис. 159, Б). За 20 дней опыта клещи не погибали даже при 0, 10 и 25% относительной влажности, теряя 13, 10 и 7% своего веса. При 50, 75 и 95% относительной влажности потери веса за указанный период достигали всего 2—5%.Из механических приспособлений, ограничивающих испарение, решающая роль принадлежит самому поверхностному слою покровов тела — эпикутикуле и особенно ее восково-липоидным компонентам. При подъеме температуры кутикулы выше определенного значения («критическая температура») межмолекулярная структура восково-липоидного слоя нарушается и проницаемость покровов для воды резко возрастает. С последним связаны общие для наземных членистоногих особенности изменения скорости испарения с повышением температуры. Вначале потери воды за счет
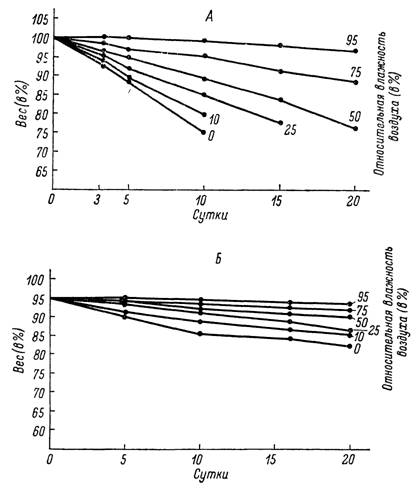
Рис. 159. Изменения веса тела голодных самцов и самок Hyalomma asiaticum в условиях различной относительной влажности при температуре 36 (А) и 26° (Б).
Средние от взвешиваний 20 клещей для каждой точки. За 100% принят вес клещей до начала опытов.испарения увеличиваются очень медленно, но после достижения определенного «критического» значения скорость потери воды возрастает скачкообразно в несколько раз. Считается, что критическая температура эпикутикулы может служить видовым признаком; чем выше ее значение, тем большей устойчивостью к высыханию отличается членистоногое (Lees, 1947; Beament, 1959).
У исследованных нами 8 видов аргасовых клещей с повышением температуры на 10° испарение возрастало всего в 1.5—2 раза. Напротив, при повышении критической температуры на 1° потери воды сразу же увеличивались в 8—10 раз (табл. 8). У Hyalomma asiaticum при повышении температуры с 35 до 50° потери воды увеличивались с 0.16 до 0.24 мг/час на одного клеща. При 52° («критическая температура») происходило внезапное увеличение испарения и при 55° скорость испарения увеличивалась в 4 раза (1.05 мг/час). С дальнейшим повышением температуры скорость испарения вновь увеличивалась более или менее равномерно (рис. 160).
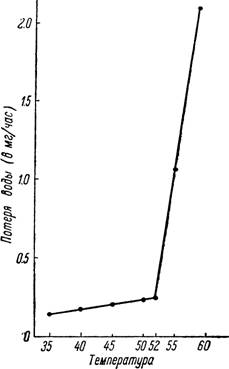
Рис. 160. Потери воды голодными самцами и самками Hyalomma asiaticum, убитыми парами аммиака, при разной температуре воздуха. Средние от взвешиваний 20 клещей для каждой точки.
Критическая температура в целом хорошо коррелирована с устойчивостью исследованных видов к высыханию. Среди аргасовых клещей самая высокая критическая температура (61°) свойственна Alveonasus lahorensis. Остальные виды по этому показателю можно разделить на две группы. У Argas vesper- tilionis, A. persicus, Ornithodorospapillipes, О. verrucosus она лежит между 49—54° ay О. capensis, О. iartakovskyi и Argas vulgaris — 42—48° (табл. 8).
Для иксодовых клещей в целом характерны более низкие значения критических температур, которые также отражают экологические особенности видов. Наиболее низкие показатели получены для высоко влаголюбивых видов, как Ixodes гісі- nus (32°) и I.
canisuga (42°). У умеренно мезофильных Dermacentor andersoni, Rhipi- cephalus sanguineus и Haylomma anatoli- cum критическая температура была около 44—45°, а у наиболее устойчивого к высыханию клеща Н. asiaticum она была выше (52°), чем у многих видов аргасовых клещей. Можно также отметить, что у ксерофильного вида Н. asiaticum критическая температура выше верхнего температурного порога жизни (48—50°), что предотвращает возможность высыхания даже в условиях перегрева. Голодающие клещи, несмотря на совершенство их морфологических и физиологических адаптаций к консервацип воды в организме, непрерывно, хотя и медленно, теряют воду за счет испарения через покровы и трахейную систему. Так, во время активности клещей Hyalomma asiaticum температура припочвенного слоя воздуха колеблется в пределах 20—30° и поверхности почвы — 20—40°. ОтносительнаяТаблица 8
Влияние температуры на скорость испарения воды через покровы клещей, убитых парами аммиака (потери веса в мг/г/час)
| Вид клеща | Критичека температура эпикутикулы, °С | Скорое пр на 11° ниже критической | гь испарен] и температ на 1° ниже критической | ИЯ воды уре ! на 1° выше критической |
| Argas persicus | 52 | 3.3 | 5.0 | 58.3 |
| A. vulgaris | 47 | 3.7 | 6.9 | 47.8 |
| A. vespertilionis | 54 | 2.9 | 4.3 | 50.9 |
| Alveonasus lahorensis | 61 | 2.8 | 3.8 | 69.3 |
| Ornithodoros papillipes | 50 | 2.6 | 4.9 | 52.7 |
| 0. verrucosus . | 49 | 3.1 | 4.4 | 51.5 |
| 0. tartakovskyi | 48 | 3.5 | 5.2 | 41.7 |
| O. capensis | 42 | 3.2 | 8.3 | 35.3 |
влажность воздуха составляет 15—50%. При 30—40° скорость испарения очень велика и существует угроза быстрого высыхания. Снижение температуры до 26° в несколько раз увеличивает продолжительность жизни клещей и значительно уменьшает скорость испарения. Однако даже при наиболее благоприятной комбинации температуры и влажности, возможной в природных условиях (20—25° и 40—50% относительной влажности), они неизбежно должны погибнуть от высыхания через 3— 4 месяца. Между тем голодные Я. asiaticum могут переживать в природе свыше года.
Сходная ситуация складывается и для лесных видов (Ixodes ricinus и /. persulcatus), фактическая продолжительность периода активности которых значительно дольше, чем можно предполагать, исходя из скорости потери воды во время нахождения на растительности. Насколько известно в настоящее время, аргасовые клещи не пьют воду, а иксодовые клещи, если и могут в отдельных случаях восполнять этим способом потери воды (Wilkinson, 1953), то не всегда им пользуются из-за отсутствия капельной влаги в их микробиотопах. Главным источником поступления воды в организм голодающих клещей, как установлено экспериментальными исследованиями со многими видами (Lees, 1946а, 1947; Browning, 1954; Балашов, 1960а; Белозеров и Серавин, 1960; Балашов и Филиппова, 1964), служит активное поглощение водяных паров из атмосферы с высокой относительной влажностью.
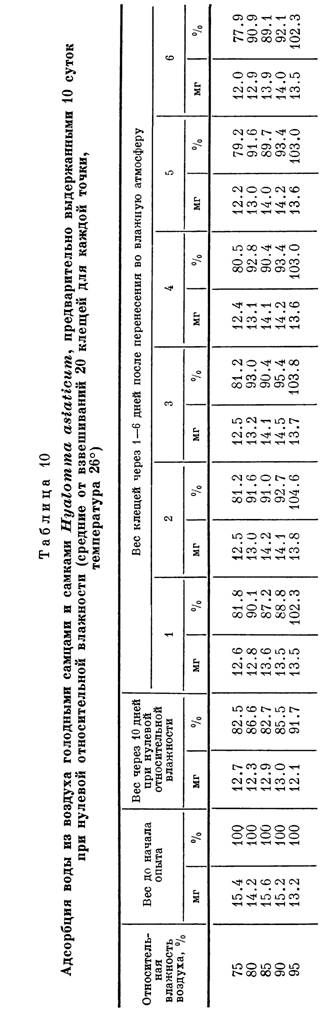
В наших опытах предварительно «подсушенные» особи большинства видов аргазид адсорбировали водяные пары при относительной влажности воздуха в 80% и выше. Как видно из табл. 9, если потери за счет испарения не превышали допустимой для организма клеща нормы в 20% от ис-
Таблица 9
Адсорбция водяных паров аргасовыми клещами, предварительно выдержанными в течение 10 суток при 20% относительной влажности (средние от взвешиваний 10 клещей для каждой точки, вес в мг)
| Вид клеща | Температура, °С | Потери веса (в %) после 10 суток при 20% относительной влажности | Вес «подсушенных* клещей | Вес через 10 суток при 80% относительной влажности | Вес «подсушенных* клещей | Вес через 10 суток при 100% относительной влажности | ||
| абс. | прирост, % | абс. | прирост, % | |||||
| Argas persicus. | 18 | 8.1 | 14.5 | 14.9 | 2.8 | 11.7 | 12.6 | 7.8 |
| 28 | 5.2 | 13.8 | 14.1 | 1.2 | 13.5 | 14.8 | 9.6 | |
| A. vulgaris. | 18 | 6.0 | 12.6 | 12.9 | 2.4 | 8.1 | 8.7 | 7.4 |
| 28 | 12.4 | 11.8 | 12.2 | 3.4 | 7.8 | 9.0 | 15.4 | |
| Alveonasus lahoren- | 18 | 3.5 | 33.0 | 32.4 | -1.8 | 34.9 | 34.7 | -0.6 |
| sis. | 28 | 6.6 | 38.4 | 37.4 | -2.6 | 36.8 | 36.3 | -1.3 |
| Ornithodoros papilli- | 18 | 8.6 | 6.7 | 7.2 | 7.5 | 6.5 | 7.0 | 7.7 |
| pes. | 28 | 7.5 | 7.1 | 7.4 | 4.2 | 7.4 | 8.1 | 9.5 |
| O. verrucosus. | 18 | 11.3 | 6.2 | 6.6 | 6.5 | 6.2 | 7.4 | 19.4 |
| 28 | 7.6 | 6.7 | 7.4 | 10.5 | 6.6 | 7.3 | 10.6 | |
| O. tartakovskyi. | 18 | 17.8 | 6.1 | 6.9 | 13.1 | 6.0 | 7.1 | 18.3 |
| 28 | 11.8 | 4.7 | 5.2 | 10.6 | 5.9 | 7.2 | 22.0 | |
| O. capensis. | 18 | 7.8 | 7.2 | 8.6 | 19.4 | 7.4 | 8.5 | 14.9 |
| 28 | 11.1 | 7.4 | 8.5 | 14.9 | 6.6 | 8.5 | 28.8 | |
Примечани е. При 80 и 100% относительной влажности у Alveonasus lahoremis наблюдалось дальнейшее уменьшение веса, отмеченное в таблице знаком минус.
ходного веса, то клещи в 10-дневный период полностью или частично восстанавливали вес, которым обладали до начала подсушивания. При этом поглощение водяных паров в общем было пропорционально потерям воды за счет испарения. Виды, терявшие в сухой атмосфере большое количество воды (Ornithodoros capensis и О. tartakovskyi), интенсивнее адсорбировали влагу из воздуха и сильнее увеличивались в весе (на 10— 29 % по сравнению с весом подсохших клещей). Напротив, более устойчивые к высыханию Argas persicus и Ornithodoros papillipes в тех же условиях увеличивались в весе всего от 1.2 до 9.6%. Только ксерофильный АІ- veonasus lahorensis вообще не адсорбировал водяные пары после предварительного выдерживания в сухой атмосфере. Можно, однако, предполо* жить, что в условиях опыта уменьшение веса особей этого вида на 3.5 — 6.6% существенно не меняло содержания воды в тканях и не вызывало потребности в восстановлении водного баланса.
Относительная влажность 80% оказалась достаточной для восстановления водного баланса и поддержания его на относительно постоянном уровне. Дальнейшее повышение влажности воздуха до точки насыщения не привело к резкому возрастанию скорости поглощения паров и соответственно увеличению в весе. По-видимому, выше точки равновесия, лежащей где-то междубО—80% относительной влажности, клещи способны поддерживать нормальную концентрацию воды в организме в значительной степени безотносительно к дефициту насыщения. Сходным образом вели себя и клещи Нуаіот- та asiaticum (табл. 10). Минимальная относительная влажность, при которой потери воды на испарение компенсируются ее адсорбцией из воздуха, составляет для них 80%. С повышением относительной влажности увеличиваются как скорость поглощения водяных паров, так и общее количество адсорбированной воды. При 80% относительной влажности клещи, потерявшие перед этим до
14% своего первоначального веса, увеличивались в весе в первые 4 дня, а затем вес их стабилизировался или даже несколько уменьшался, не достигнув исходной величины. При 95% относительной влажности клещи уже за 1 сутки не только восстанавливали исходное содержание воды в организме, но в ряде случаев оно было даже несколько выше, чем перед началом опытов. Общий вес воды, поглощенной клещами за это время, достигал 12% от веса тела «подсушенных» особей.
Точка равновесия, при которой потери воды на испарение полностью восполняются ее активным поглощением из воздуха, служит важным экологическим показателем, характеризующим степень гигрофильное™ разных видов. Для сравнения можно указать, что у Ixodes ricinus (Lees, 1946а) она близка к 92%, у Hyalomma asiaticum к 80% и у аргасовых клещей к 60—80% относительной влажности.
Предполагается, что водяные пары поглощаются гиподермальными клетками (Lees, 1946а). Этот процесс подвержен значительным индивидуальным отклонениям и, вероятно, зависит от общего физиологического состояния организма, так что в одинаковых физических условиях и после одинакового обезвоживания организма скорость адсорбции воды неодинакова даже среди особей одного вида. У недавно перелинявших или напитавшихся особей способность к поглощению водяных паров временно исчезает. Анестезия клещей высокими концентрациями С02 (Browning,
1954) или замазывание дыхалец (Белозеров и Серавин, 1960), вызывавшие резкие нарушения физиологических процессов, также приводили к прекращению адсорбции воды.
Особенностями регулирования водного баланса можно объяснить периодические вертикальные миграции многих видов иксодовых клещей, а также описанные в литературе сезонные перемещения аргасовых клещей внутри нор (Ушакова, 1956; Бусалаева, 1960). Подстерегание хозяев на растительности, поверхности почвы или в верхних отделах нор связано с неизбежными потерями воды на испарение. После понижения содержания воды в организме до определенного критического уровня характер поведенческих реакций изменяется и клещи перемещаются в растительную подстилку, трещины почвы или в норы. Здесь, в условиях высокой относительной влажности, происходит активное поглощение водяных паров и содержание воды в организме восстанавливается до нормального уровня. После этого клещи вновь переходят в более сухие стации, где имеются наибольшие шансы на встречу с хозяином.
Еще по теме ПРИСПОСОБЛЕНИЯ К УМЕНЬШЕНИЮ ПОТЕРЬ ВОДЫ ОРГАНИЗМОМ:
- Потери воды организмом
- Сохранение воды в организме
- Поступление воды в организм
- Избыточное накопление воды в организме
- Клинический пример 5. (синдром потери плода, ОК, потеря зрения)
- Перенос на носилочных устройствах и приспособлениях.
- Алгоритмы обработки информации канала ультразвукового зондирования толщи воды (УЗК) и оптического локатора поверхности воды.
- Механизмы заражения воды в водопроводах; микрофлора водопроводной воды.
- Уменьшение объема внеклеточной жидкости (гиповолемия)
- 4.4.1. Нарушения, характеризующиеся уменьшением количества костной ткани
- Уменьшение количества атипичных клеток
- Уменьшение токсического действия металла в результате хелатообразования, антидоты
- Уменьшение физиологической потребности (V)
- 28.Уменьшенный желчный пузырь.
- №3 Организм человека как единая биологическая система. Уровни организации человеческого организма: молекулярный, клеточный, тканевой, органный, системный, организменный.
- Параграф восьмой. Увеличение и уменьшение слезного мясца3
- Работа с мысленными образами для уменьшения боли