Иксодовые клещи.
Среди иксодовых клещей наблюдаются определенные различия в строении прокутикулы между подсемействами Ixodinae и АтЫуоттіпае, рассмотренные на примере клещей Ixodes ricinus и Hyalomma asiaticum.
У недавно перелинявших особей обоих видов покровы образованы одной недифференцированной прокутикулой, которая интенсивно окрашивается анилиновым синим (после окраски по Маллори или азокармином по Гейденгайну). В дальнейшем на стадии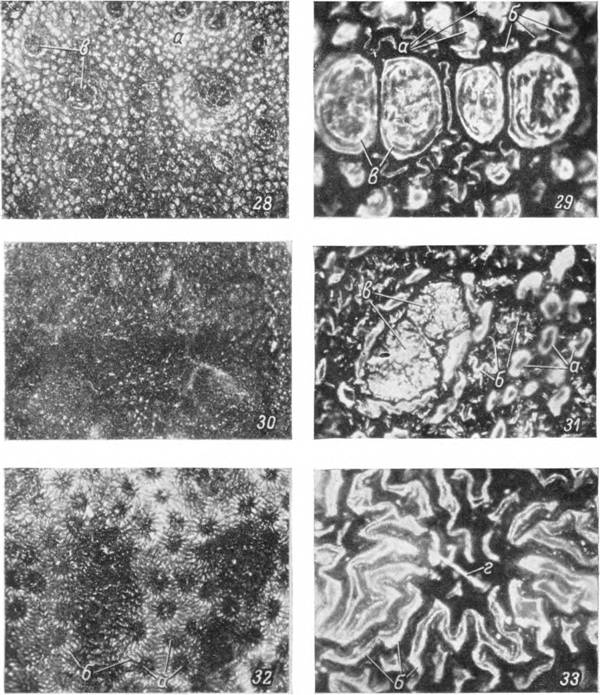
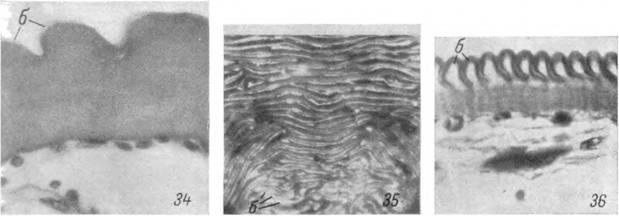
Рис. 28—36. Строение дорсальной поверхности идиосомы.
28, 29 — самка А rgas persicus; ЗО, 31 — самка Ornithodoros papillipes; 32—36 — Alveo- nasus lahorensis: 32, 33 — самка, 35 — личинка, 34 — поперечный срез через покровы самки, 36 — то же личинки. Микрофотографии в падающем свете: 28, 30 и 32 — Х20; 29, 31, 33 — Х110; 34 — Х600; 35 — Х600; 36 — ХІ350.
а — бугорки; б — складки; в — мышечные диски; г — щетинки.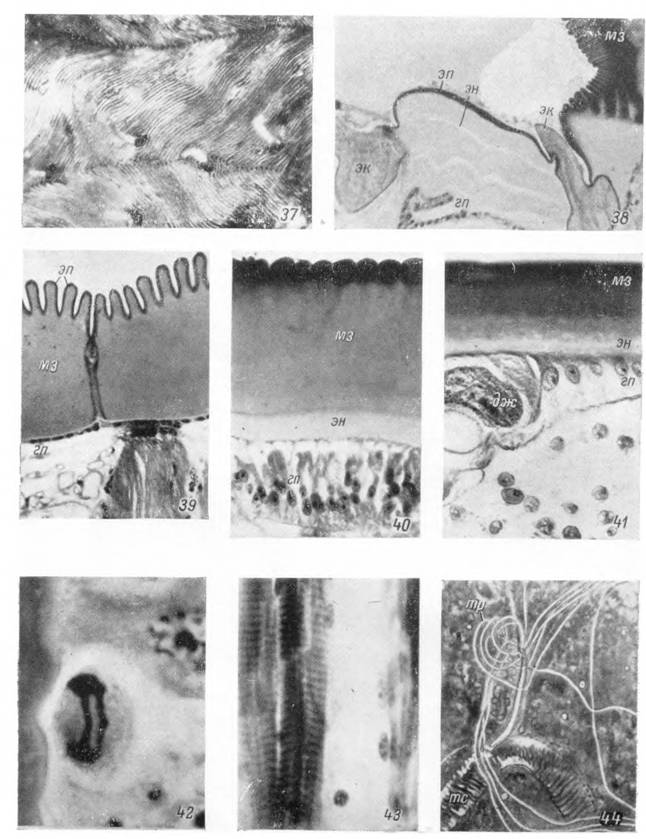
Рис. 37—44. Покровы тела, мышцы и трахеи самки Hyalomma asiaticum.
37 — участок поверхности аллоскутума голодного клеща; 38 — место сочленения коксы с идиосомой; срезы через растяжимую кутикулу клеща: 39 — голодного, 40 — питающегося и 41 — закончившего питание; 42 — митотическое деление гиподермальной клетки на стадии питания; 43 — участок субкоксальной мышцы; 44 — отхождение пучка трахеол от трахейного ствола. Микрофотографии: 37 — Х20; 38 и 44 — X 200; 39—41, 43 — Х400; 42— Х2000.
эп — эпикутикула; эн — эндокутикула; эк — экзокутикула; мз — мезокутикула; гп — гиподерма; дж — дермальная железа I типа; тс — трахейный ствол; тр — трахеолы.
Рис. 45—53. Строение покровов Argas persicus (срезы).
45— мышечный диск; 46 — отдельный бугорок; 47 — растяжимый участок идиосомы самки; 48 — голодная нимфа; 49 — напитавшаяся нимфа I; личинка: 50 — голодная, 51 — недавно начавшая питаться, 52 — заканчивающая стадию роста и 53 — полностью напитавшаяся.
цс — цементный, еле — восково-липоидный, кс — кутикулиновый слои эпикутикулы; дп — эпикутикула; эк — экзокутикула мышечных дисков; мз — мезокутикула; эк — эндокутикула; гп — гиподерма; дж — дермальная железа; пр — проток дермальной железы; м — дорсо-вентральные
мышцы; х — хета.
послелиночного доразвития первичная прокутикула HyaLomma asiaticum подвергается частичной склеротизации и превращается в первичную мезокутикулу, которая интенсивно окрашивается кислым фуксином или азокармином (рис.
39). У Ixodes ricinus на стадии послелиночного доразвития первичная прокутикула также превращается в мезокутикулу, но одновременно под ней откладывается слой эндокутикулы, красящийся анилиновым синим и другими кислыми красителями (рис. 177). Слой субкутикулы у голодных клещей очень тонкий (меньше 1 мк) и в большинстве случаев обычными гистологическими методами не выявляется.Мезокутикула голодных особей имеет сходное строение у обоих видов. Она весьма однородна и состоит из нескольких слоев общей толщиной в 14—18 мк у самок Hyalomma asiaticum и несколько тоньше у Ixodes ricinus. Эти слои идут либо параллельно поверхности, и тогда видно, что прокутикула образована 3—4 подобными макроструктурными единицами, или же под некоторым углом к поверхности. У только что перелинявших особей в самом нижнем слое при использовании фазового контраста удается рассмотреть следы более тонкой слоистости. Позднее пластинчатое строение становится неразличимым. Противоположная картина наблюдается с поровыми канальцами. В первом случае они неразличимы, а во втором, напротив, выступают очень отчетливо. Поровые канальцы в мезокутикуле неразветвленные и слабо волнистые, а в эи- докутикуле сильно изогнутые и ветвящиеся.
Эндокутикула у голодных особей /. ricinus имеет хорошо развитую горизонтальную слоистость. Отдельные слои слабо волнистые, толщиной около 1 мк. По аналогии с таковыми у насекомых, их, вероятно, следует считать правильно чередующимися участками различной оптической плотности, создающими видимость псевдопластинчатой структуры.
С началом питания иксодовых клещей наряду с растяжением покровов идиосомы происходит отложение новых слоев кутикулы и изменение строения слоев, образовавшихся во время линьки. Эти процессы, как и секреторная активность гиподермальных клеток, описываются в разделе «Приспособления к увеличению объема тела во время питания» (см. стр. 117—126).
Склеротизованная кутикула. У иксодовых клещей из твердой кутикулы построены гнатосома, спинной щиток, конечности, анальный клапан, вентральные щитки самцов и ряд других мелких склеритов (рис.
38). Описанные участки отличаются значительной твердостью и очень малой эластичностью. К растяжению они не способны, и в период кровососания их линейные размеры практически не меняются. Эпикутикула гладкая, обычного строения. Прокутикула целиком склеротизована, так что ее следует считать построенной из одной экзокутикулы. Последняя, как и у насекомых, без предварительной обработки диафанолом вообще не окрашивается и имеет на срезах естественный янтарно-желтый оттенок. После размягчения диафанолом она начинает краситься кислым фуксином или азокармином. На срезах видно, что вся ее толща пронизана тончайшими, слабо извитыми поровыми канальцами. Какие-либо следы продольной слоистости в ней неразличимы. При наблюдении в поляризованном свете экзокутикула дает очень сильное двойное лучепреломление, вероятно, за счет высокой степени ориентации белково-хитиновых мицелл.В спинном щитке нижний слой кутикулы бывает часто окрашен значительно сильнее верхнего. Так, у самок Hyalomma asiaticum нижний, интенсивно пигментированный слой составляет 10—12 мк, а верхний около 30 мк. По гистохимическим особенностям оба описанные слоя, однако, идентичны. Поэтому от гомологизации нижнего слоя с эндокутикулой, распространенной во многих работах, следует отказаться.
У аргасовых клещей из твердой кутикулы построены покровы конечностей, гнато'сомы, перитремы, дорсальная пластинка у личинок некоторых видов (Argas persicus, Alveonasus lahorensis, Ornithodoros coniceps и др.) и диски для прикрепления мышц, особенно развитые у видов рода Argas (рис. 45).
В конечностях и гнатосоме аргазид эндокутикула и мезокутикула отсутствуют и они состоят из одной экзокутикулы янтарно-желтого цвета, пронизанной поровыми канальцами и почти не окрашивающейся железным гематоксилином и азокармином. Красится лишь очень узкая (2 мк) зона на границе с гиподермой. Общая толщина экзокутикулы составляет 15—30 мк. Гиподермальный слой немного толще обычного. В дорсальной пластинке личинок, достигающей у Alveonasus lahorensis 5—6 мк толщины, экзокутикула неоднородна и состоит из наружной и внутренней зон, различающихся по интенсивности окрашивания азо- кармином.
Под ними располагается эндокутикула в 1.5—2 мк. В дисках Argas persicus также сохраняется тонкий слой эндокутикулы.Субкутикула в твердых участках не выявляется. Гиподермальные клетки, подстилающие экзокутикулу, мелкие, границы между ними почти неразличимы. Во время питания гиподермальный эпителий не проявляет каких-либо признаков усиления функциональной активности. Толщина твердой кутикулы значительно варьирует в зависимости от локализации и видовой принадлежности. Однако во время кровососания она всегда остается постоянной.
Кутикула сочленений. Особый тип кутикулы встречается в суставах конечностей, камеростомальной складке и других подвижных сочленениях у иксодовых клещей. По своему строению она очень сходна с сочленовными мембранами у других членистоногих (рис. 38). Снаружи она ограничена хорошо развитым кутикулиновым слоем эпикутикулы, интенсивно красящимся азокармином или кислым фуксином. Непосредственно под ним располагается эндокутикула, достигающая местами толщины в 60—100 мк. Последняя состоит из отдельных горизонтальных слоев толщиной в 1—2 мк. Поровые канальцы в ней неразличимы. При обработке азокармином по Гейденгайну или по Маллори она окрашивается анилиновым синим в ярко-голубой цвет, а после обработки по Мил- лоту прокрашивается световым зеленым. Субкутикула развита слабо и с трудом выявляется. Под ней располагается тонкий слой гиподермальных клеток. Местами они исчезают, а сохраняющиеся лежат отдельными небольшими островками.
Сходным строением обладают и сочленовные мембраны аргасовых клещей. Они также не подвержены склеротизации и построены из относительно толстой (3—4 мк) эпикутикулы со слегка зубчатой поверхностью и однородной эндокутикулы (20—30 мк). В последней очень четко выступает пластинчатая структура. Отдельные слои связаны между собой слабо, так что иногда при фиксации происходит их расслоение.
Гистохимические особенности кутикулы. Одним из основных различий в химическом составе эпикутикулы и прокутикулы клещей оказалось отсутствие в первой хитина, как и у остальных членистоногих.
В прокутикуле хитин легко выявляется хитозановой реакцией, тогда как обычные методы окраски срезов на хитин раствором иода в хлористом цинке (после предварительной обработки диафанолом) или по методу ШИК не дают положительного результата (табл. 1). Лишь в отдельных случаях слабое окрашивание наблюдалось в гнатосоме, конечностях и склеритах, построенных из экзокутикулы. По-видимому, молекулы хитина в кутикуле иксодовых клещей очень прочно связаны в гликопротеиновых комплексах и выявляются только после разрушения последних, как в случае с хитозановой реакцией.Присутствие белков в прокутикуле легко выявляется окраской бромфеноловым синим и биуретовой реакцией. Особенно четкие результаты дает первый метод. Интенсивность окраски максимальна в мезоку- тикуле. При этом в бугорках и гребнях аргазид часто развивается не обычное синее, а яркое фиолетовое окрашивание, связанное с метахромазией. Последняя свойственна и мезоку- тикуле иксодид, особенно у начавших питаться особей. Эндокутикула красится только в синий цвет и значительно слабее, а экзокутикула практически не окрашивается, приобретая лишь очень слабый синеватый оттенок. Окраска эпикутикулы всегда дает отрицательный результат. Отрицательная реакция в склеротизованных участках и эпикутикуле связана не с отсутствием в них белков, а с блокированием их реактивных групп в сложных комплексах вследствие процессов химического дубления.
Значительный интерес представляет распределение в кутикуле веществ, содержащих ароматические группы. Очень интенсивную окраску приобретают по миллоновой и ксанто- протеиновой реакциям кутику- линовый слой, эпикутикулы, экзокутикула и наружная зона мезокутикулы. От эпикутикулы к гиподерме окраска мезокутикулы убывает, вероятно, отражая градиент концентрации этих веществ. Эндокутикула дает отрицательный резуль тат.
Аргентаффиновая реакция дает результаты, сходные с двумя предыдущими. Аммиачный раствор серебра особенно интенсивно восстанавливается до металлического в кутикули- новом слое эпикутикулы и значительно слабее в экзокутикуле и наружной зоне мезокутикулы.
Во внутренней зоне мезокути-пулы и эндокутикуле, за исключением протоков дермальных желез и хет, почернения не наблюдается.
На срезах, изготовленных на замораживающем микротоме после формалиновой фиксации, липоиды окрашиваются Суданом черным лишь в восково-липоидном слое эпикутикулы. Судя по их высокой критической точке, они несомненно обладают высокой температурой плавления. В прокутикуле липоиды выявляются лишь после обработки диафанолом. Последний наряду с разрушением ароматических колец освобождает также связанные липоиды. После подобной обработки они начинают окрашиваться Суданом в кутикулиновом слое, экзокутикуле, и в меньшей степени в мезокутикуле.
Характерную особенность кутикулы представляет высокое содержание серы, что свойственно и акариформным клещам (Hughes, 1959). Нитропруссидная реакция на дисульфидные группы дает особенно интенсивное окрашивание в кутикулиновом слое и экзокутикуле. Мезокутикула дает неопределенный, а эндокутикула отрицательный результат.
Ни в одном из слоев кутикулы Hyalomma asiaticum и Ixodes ricinus, вопреки утверждениям Г. Фрика (Frick, 1936) и П. Шульце (Schulze, 1955), нам не удалось обнаружить гуанин. Мурексидная реакция всегда давала отрицательный результат. П. Шульце идентифицировал присутствие гуанина в покровах по наличию двойного лучепреломления. Однако в рассматриваемых случаях указанный критерий неприменим, так как белково-хитиновые мицеллы и кутикулин эпикутикулы отличаются сильной собственной анизотропией.
Субкутикула дает положительную реакцию на полисахариды и белки и отрицательную на хитин и, вероятно, как и у насекомых, построена из мукопротеидов.
Из приведенных данных видно, что химический состав кутикулы аргасовых и иксодовых клещей в основном сходен с насекомыми и изученными паукообразными. Прокутикула построена из хитино-протеиновых макромолекул-мицелл, в которых хитин прочно связан с белком, так что без их разрушения он обычно не выявляется. Прокутикула импрегнируется липоидами и белками, соединяющимися в липо-протеиновый комплекс. Существенный компонент кутикулы составляют ароматические вещества. Максимальная их концентрация в кутикулиновом слое и экзокутикуле, представляющих наиболее склеротизованные зоны, указывает на сходство процессов затвердевания покровов у клещей и насекомых. Вероятно, при соединении свободных аминокислотных групп липо-протеинового комплекса с хинонами происходит химическое дубление кутикулы — завершающий этап склеротизации. Возможно, что у клещей, как и у скорпионов, в затвердевании покровов, и особенно кутикулинового слоя, определенную роль играет образование дисуль- фидных мостиков.
МУСКУЛАТУРА
Клещи надсем. Ixodoidea обладают хорошо развитой мускулатурой, строение которой с различной степенью подробности описывается в нескольких общеанатомических работах (Robinson a. Davidson, 1913— 1914; Ruser, 1933; Douglas, 1943). По местоположению и функциям различают четыре основные группы мышц. Мускулатура тела представлена несколькими группами дорсо-вентральных мышц (краевые, генитальные и анальные), места прикрепления которых к покровам идиосомы обозначены соответствующими бороздами у иксодовых клещей и мышечными дисками у аргазид (рис. 54, 55). Сокращения дорсо-вентральной мускулатуры обеспечивают изменения объема и формы идиосомы и играют важную роль в движениях гнатосомы (за счет изменения внутриполостного давления), при актах дефекации и откладки яиц. Мускулатура ног представлена мощными косыми пучками коксальных и субкоксальных групп мышц, а в каждом членике конечности — антогонистически действующими сгибателями и разгибателями (рис. 56, 57). Мускулатура гнатосомы рассматривается при описании ротового аппарата, а мускулатура внутрен-
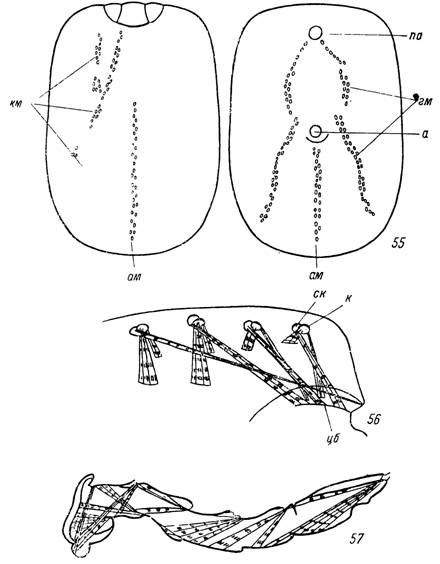
Рис. 54—57. Мускулатура идиосомы самки Hyalomma plum- Ьешп и ноги личинки Ixodes ricinus (но Ruser, 1933). Места нрикренления дорсо-вентральных мышц: 54 — на дорсальной стороне идиосомы, 55 — на вентральной стороне идиосомы; 56 — коксальные и субкоксальные мышцы Н. ріит- beum\ 57 — сгибатели и разгибатели члеников ноги /. ricinus.
км — краевые, ам — анальные, гм — генитальные пучки дорсо- вентральных мышц; а — анус; по — половое отверстие; к — кокса; ск — субкокса; цб — цервикальная борозда.
них органов — в разделах, касающихся строения органов пищеварения, размножения и выделения.
Цитологические и физиологические особенности мышц клещей совершенно не изучены. На основании наших гистологических препаратов и наблюдений над отпрепарированными органами в фазовом контрасте можно лишь указать, что, как и у насекомых, мышечные волокна клещей поперечнополосатого типа. Пучки миофибрилл занимают центральное положение внутри волокон, а саркоплазма с ядрами окружает их с периферии (рис. 43, 359).Органы пищеварения клещей представлены ротовым аппаратом с обширной предротовой полостью (рассматриваются в отдельном разделе), парой слюнных желез, глоткой, пищеводом, средней кишкой (желудок и боковые дивертикулы), тонкой кишкой, ректальным пузырем и прямой кишкой, открывающейся анальным отверстием (рис. 58, 59). Слюнные железы, глотка, пищевод и прямая кишка являются эктодермальными производными; средняя и тонкая кишка, ректальный пузырь и открывающиеся в него мальпигиевы сосуды — эндодермальные производные.
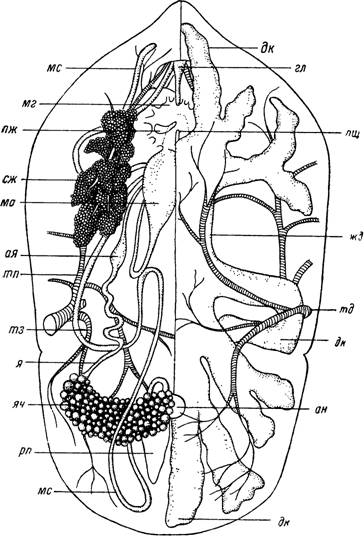
Рис. 58. Вскрытая со спинной стороны самка Огпі- ihodoros papillipes, левая половина кишечника удалена.
В литературе накоплен обширный фактический материал об анатомическом строении пищеварительного тракта клещей Ixodes ricinus (Pagenstecher, 1861;
Bonnet, 1907; Суворов,
1908; Nordenskiold, 1908,
1911; Samson, 1909a), Boop- hilus annulatus (Williams,
1905; Dennis, 1932), Der- macentor andersoni (Dou- glas, 1943), Haemaphysalts flava (Saito, I960), Rhi- picephalus appendiculatus (Till, 1961), Ornithodoros savignyi (Christophers,
1906), O. coriaceus (True,
1932), Argas persicus (Robinson a. Davidson, 1913—
1914), A. vespertilionis (Roshdy, 1961a), A. boueti (Roshdy, 1962), A. transga- riepinus (Roshdy, 1963).
ап — анус; ая — ампуловидное расширение яйцевода; гл — глотка; жд — желудок; ма — матка; мс — мальпигиев сосуд; мг — мозг; пщ — пищевод; пж — придаточная железа; рп — ректальный пузырь; сж — слюнная железа; дк — дивертикулы средней кишки; трахейные стволы: тд — дорсальный, тз — задний и тп — передний; я — яйцевод; яч — яичник.
Сведения о строении кишечника аргасовых клещей приводятся также у Е. Н. Павловского (1935), а сравнительно-анатомический анализ расчленения средней кишки вместе с описанием ее строения
для многих видов иксодовых и аргасовых клещей дается П. Шульце (Schulze, 1942а) и в наших работах (Балашов, 1957в, 19616).
Слюнные железы. У клещей надсем. Ixodoidea слюнные железы представлены парой гроздевидных, вытянутых в передне-заднем направлении по бокам идиосомы органов, построенных из альвеол нескольких типов (см. стр. 135—152), сидящих на сильно ветвящихся выводных протоках, связанных между собой рыхлой соединительной тканью и разветвлениями трахей. У аргасовых клещей (рис. 58) слюнные железы весьма компактныи лежат над коксами первых трех пар ног. Размеры их у голодных или недавно напитавшихся особей примерно одинаковы. У иксодовых клещей, и особенно в подсем. АтЫуоттіпае, слюнные железы значительно крупнее, их задние концы заходят за IV коксы и в них более четко проступает тенденция к расщеплению на отдельные грозди. Размеры их определяются физиологическим состоянием клещей. У питающихся особей слюнные железы значительно крупнее, чем у голодных (рис. 109—110).
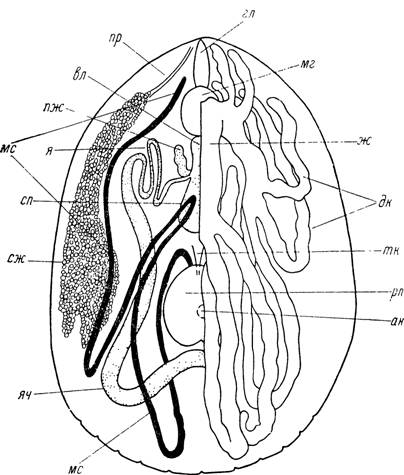
Рис. 59. Вскрытая со спинной стороны самка Нуаіотта asiaticum, левая половина кишечника удалена.
гл — глотка; мг — мозг; ж — желудок; дк — дивертикулы средней кишки; тк — тонкая кишка; рп — ректальный пузырь; ан — анус; ліс — мальпигиев сосуд; яч — яичник; сж — слюнная железа; сп — семеприемник; я — яйцевод; пж — придаточная железа; вл — влагалище; пр — проток слюнной железы.
Выводные протоки слюнных желез, являющиеся производными гипо* дермы, представляют глубокие внутренние впячивания стенок тела. Начинающиеся от слюнного резервуара парные главные выводные протоки почти округлые на поперечном разрезе. Войдя в слюнные железы, они дают многочисленные разветвления. Альвеолы сидят и на главных протоках, но основная их масса локализуется на вторичных разветвлениях. Изнутри главные протоки и их разветвления выстланы сравнительно толстой кутикулой из тонкой наружной кутикулиновой эпикутикулы и внутренней эндокутикулы. В последней на всем протяжении протоков проходит более плотная спиральная нить, напоминающия тенидиальную нить трахейных стволов. Она выполняет чисто механические функции, предотвращая спадение внутреннего просвета выводных протоков (рис. 64), и этим, по-видимому, можно объяснить ее конвергентное сходство с аналогичными структурами трахейных стволов. Кути- кулиновый слой выстилки протоков заходит и внутрь альвеол, где покрывает короткий внутриальвеолярный выводной проток, сообщающийся с внутренней полостью альвеолы. Границы между отдельными гиподермальными клетками выстилки выводных протоков плохо различимы в световом микроскопек но очень четко видны на электронных фотографиях (Джафаров, 19656). Ядра их средних размеров, овальные, а плазма не содержит каких-либо включений, напоминающих продукты секреторной активности. Мышечные волокна в их стенках обнаружить не удалось.
Глотка. У иксодовых клещей глотка сильно вытянута в передне- заднем направлении и веретеновидной формы (рис. 25). На уровне ротового отверстия просвет глотки еще сохраняет характерную для пищевой борозды треугольную форму (рис. 193), но по мере удаления от лабрума на ее потолке появляется сильно склеротизованный гребень, приобретающий клиновидную форму и глубоко вдающийся в просвет (рис. 194). Снизу глотка поддерживается хитиновым полукольцом так называемых глоточных опор. Последние представляют вентро-латеральные выросты межкоксального моста, срастающиеся между собой по средней линии. На уровне клиновидного гребня они сливаются с вентро-латеральными стенками глотки и теряют связь с межкоксальным мостом. Еще дальше назад клиновидный гребень начинает понижаться и исчезает в медиальной части, тогда как его вилкообразно расщепленные латеральные части расходятся в стороны и вниз и образуют крыловидные отростки (рис. 25, 194, 195). Дорсально от них отходят мощные мышечные пучки, прикрепляющиеся противоположными концами к краям субхелицеральной пластинки, а вентрально — мышцы к вентро-латеральным углам гнатосомы. Обе группы мышц действуют антагонистически, и их попеременное сокращение обеспечивает поднятие или опускание клиновидного гребня в просвете глотки. На этом же уровне от вентрально-латеральных стенок глотки отходят пучки мышц-расширителей, а их внутреняя поверхность несет косые ряды мелких зубчиков. Судя по расположению мышц и скле- ротизованных элементов, передний отдел глотки, по-видимому, выполняет функции переднего глоточного клапана.
Вентральные несклеротизованные утолщения глоточных стенок, выполняющие, по мнению Д. Грегсона (Gregson, 1960), функции второго глоточного клапана, у изученных нами видов развиты значительно слабее. Они находятся на уровне слияния стенок глотки с глоточными опорами и окрашиваются при азановой методике в синий цвет. Ввиду их незначительных размеров они вряд ли могут замыкать глоточный просвет и скорее обеспечивают некоторую гибкость сочленения глотки с аподемаль- ными структурами гнатосомы.
Собственно глоткой следует считать отдел, расположенный позади от клиновидного гребня (рис. 195, 196). От вентрального угла первично треугольного просвета отходят два луча в вентро-латеральном направлении; дорсо-латеральные углы глотки также вытягиваются, и на поперечном разрезе орган приобретает характерную форму двух букв Y, соединенных основаниями. Стенки глотки образованы парой длинных, изогнутых внутрь хитиновых латеральных лопастей и двумя парами более коротких вентральных и дорсальных лопастей. Постоянное положение глотки в полости тела обеспечивается главным образом латеральными мышцами-расширителями. Пучки последних идут от средней линии латеральных лопастей к боковым стенкам основания гнатосомы. В передней трети органа он поддерживается также несколькими парами дорсальных расширителей. Они отходят от концов дорсальных лучей и прикрепляются снизу к субхелицеральной пластинке (рис. 194). От наружной поверхности вентральных лучей на всем их протяжении отходят пучки вентральных мышц-расширителей. Антагонисты расширителей глотки представлены группой кольцевых мышц-сжимателей, отходящих от концов лучей и окружающих глотку мощным мышечным слоем (рис. 195, 196). Места прикрепления мышц-расширителей и сжимателей чередуются в правильном порядке. В местах, где отсутствует прикрепление мышц, хитинизированные стенки глотки окружены слоем паренхиматозной ткани гиподермального происхождения.
У аргасовых клещей глотка такого же строения, что и у иксодид, и лишь несколько короче и толще. Ротовое отверстие и самая дистальная часть глотки на поперечных срезах имеют форму перевернутого треугольника (рис. 203). Несколько далее назад появляется срединный вентральный вырост, а затем два дорсо-латеральных, а глотка принимает характерную трехлучевую форму (рис. 204—205). У вершины каждый из лучей в свою очередь вилкообразно расщепляется. При расширении глотки форма ее просвета приближается к мальтийскому кресту. Каких- либо морфологически оформленных клапанов на ее переднем и заднем концах нам обнаружить не удалось. Возможно лишь, что клиновидный выступ нижней лабральной стенки, расположенный на дорсальной поверхности рта, может замыкать вход в глотку при ее сокращении. Снаружи, как и у иксодид, она окружена кольцами мышц ретракторов, правильно чередующихся с пучками дилататоров. Дорсальные мышцы- расширители начинаются на утолщенных латеральных гребнях эпистома, а вентральные и вентро-латеральные — на нижних стенках гнатосомы. Кроме описанных мощных дорсальных дилататоров, от переднего конца глотки отходят несколько коротких мышечных пучков. Они идут к самому переднему концу эпистома, формирующему пол слюнной полости, и, вероятно, контролируют объем последней (рис. 198).
Пищевод. В задней части глотки ее диаметр и толщина склеротизован- ных стенок постепенно уменьшаются, расширители исчезают, и она переходит в трубчатый пищевод. Последний представляет короткую, тонкую трубку, соединяющую глотку со средней кишкой. От конца глотки он сперва делает небольшой изгиб в нижне-переднем направлении, а затем круто поворачивает вперед и вверх, впадая в желудок с вентральной стороны у его переднего конца. На своем пути он проходит сквозь мозг, входя в него в передневентральной части и выходя в заднедорсальной (рис. 60). Задний конец пищевода впячивается в просвет желудка наподобие телескопической трубки. Образующуюся при этом кольцевую складку называют провентрикулярным клапаном и считают, что она препятствует вытеканию содержимого средней кишки в глотку. При движении крови вперед стенки его отгибаются и закрывают вход в пищевод.
На поперечных срезах пищевод почти округлый. Он построен из слабо вытянутых эпителиальных клеток, тесно прижатых друг к другу
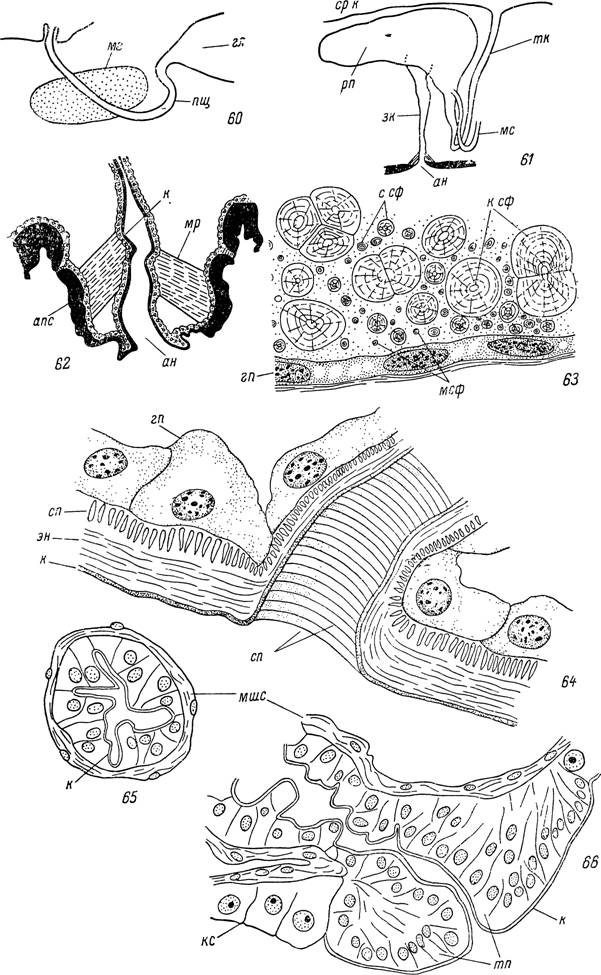
Рис. 60—66. Строение пищевода, ректального пузыря и задней кишки. Ornithodoros papillipes: 60 — соединение пищевода с глоткой и средней кишкой, 61 — соединение ректального пузыря с мальпигиевыми сосудами, тонкой и задней кишками, 62 — задняя кишка самки, 63 — участок стенки ректального пузыря самки Ixodes ricinus\ Hyalomma asiaticum: 64 — место впадения вторичного выводного протока в главный выводной проток слюнной железы, 65 — передняя часть пищевода, 66 — место впадения пищевода в среднюю кишку самки.
гл — глотка; мг — мозг; пщ — пищевод; срп — средняя кишка; рп — ректальный пузырь; тпп — тонкая кишка; зк — задняя кишка; мс — мальпигиевы сосуды; ан — анальное отверстие; мр — мышцы-расширители анального отверстия; апс — створки анального клапана; к — кутикулярная выстилка; эн — эндокутикулярный слой; сп — спиральная кутикулярная нить стенки выводного протока; мшс — мышечный слой и соединительнотканная оболочка; тпп — телескопическое впячи- вание пищевода в полость средней кишки; г — клетки стенки средней кишки; мсф — мелкие, ссдб — средние и ксф — крупные сфериты гуанина; пр — клетки стенки ректального пузыря; гп — гиподермальные клетки.
и образующих многочисленные мелкие складки. Клетки не проявляют каких-либо признаков секреторной активности или образования слизи. Плазма их бедна включениями. Ядра довольно мелкие, овальные. За исключением периодов личиночной и нимфальной линек, они всегда находятся в покоящемся состоянии. Апикальные концы клеток выстланы кутикулярным слоем, окружающим весь просвет пищевода, базальные покоятся на очень тонкой базальной мембране. Снаружи пищевод окружен сплошным слоем кольцевой и отдельными пучками продольной мускулатуры (рис. 65).
Стенки складок провентрикулярного клапана относятся к производным первичной передней кишки. Клетки их сильно вытянуты и сжаты, так что местами создают впечатление многослойного эпителия. Они не выполняют ни секреторной, ни пищеварительной функций и плазма их не содержит включений. Снаружи они покрыты тонкой кутикулой, резко обрывающейся на границе с клетками средней кишки. Мышечный слой пищевода в провентрикулярной складке отсутствует (рис. 66).
Средний отдел кишечника. Средняя кишка клещей эндодермального происхождения и занимает значительную часть полости идиосомы, прикрывая с дорсальной стороны внутренние органы. Она подразделяется на центральный отдел, или желудок, и отходящие от него парные трубковидные или лопастевидные дивертикулы (рис. 58, 59). Форма и число последних кажутся, на первый взгляд, весьма изменчивыми. Однако на основании сравнительного изучения строения кишечника различных видов и на разных фазах развития выявляются особенности, свойственные как всему надсем. Ixodoidea в целом, так и отдельным группировкам внутри него. При этом в строении средней кишки следует различать весьма постоянное первичное расчленение и очень изменчивое вторичное расщепление первичных отростков. Первичные отростки (дивертикулы) образуются на ранних стадиях эмбриогенеза в виде симметричных не- расчлененных выростов первичной средней кишки. Ю. Вагнер (1894) показал, что у эмбрионов Boophilus calcaratus первоначально образуются дивертикулы 2—7-й пар, а отростки 1-й пары появляются значительно позднее. У Ixodes ricinus они развиваются у перелинявших личинок (Falke, 1931). Однако, по более новым наблюдениям (Aeschlimann, 1958). у эмбрионов Ornithodoros moubata сразу же закладываются все 7 пар дивертикулов. Возможно, что запаздывание закладки 1-й пары дивертикулов связано с ее значительной редукцией у многих видов иксодид и имеет вторичный характер. В этом случае следует принять за исходное число дивертикулов для всего надсемейства 7, а отклонения от него считать вторичными изменениями.
Аргасовым клещам в целом свойственно сохранение относительно развитой, часто мешковидной центральной части средней кишки и очень сильное вторичное расщепление первичных отростков, наблюдающееся уже на личиночной фазе. У Ornithodoros papillipes сохраняются все семь пар первичных отростков (рис. 67 и 68). Из них первые четыре пары, называемые передне-боковыми отростками, отходят латерально одним общим основанием от переднего конца желудка, а уже далее они разделяются на самостоятельные дивертикулы. В ряде случаев, возможно, в зависимости от степени растяжения кровью, создается впечатление, что от общего основания сперва отходит лишь пара отростков, вскоре в свою очередь дихотомически расщепляющихся. Передне-боковые отростки тесно связаны с пальпами и первыми тремя парами ног. У личинок и нимф часто наблюдается их вхождение в конечности. Пятую пару принято называть срединными отростками. Они отходят почти перпендикулярно к продольной оси тела, а дойдя до боковых стенок, подгибаются вентрально и возвращаются к средней линии,-где круто поворачивают вперед и заканчиваются под передним концом желудка. Две последние пары, называемые задне-боковыми, отходят общим основанием от заднего конца желудка и заканчиваются у задней стенки тела. Они,
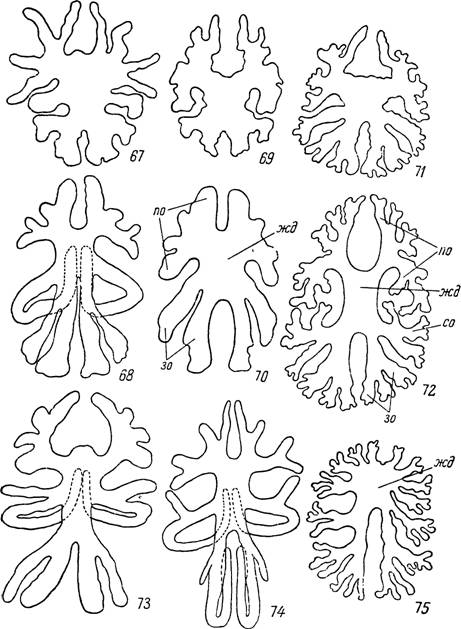
Рис. 67—75. Расчленение средней кишки аргасовых клещей. 67 — личинка и 68 — самка Ornithodoros papillipes; 69 — личинка и 70 — самка Alveonasus lahorensis; 71 — личинка и 72 — самка Argas persicus; 73 — самка Ornithodoros alactagalis; 74 — самка О. coniceps; 75 — самка A rgas vespertilionis,
жд — желудок; отростки средней кишки: по — переднебоковые, со — срединные и зо — заднебоковые.
как правило, бывают наиболее сильно развиты. Дистальные их концы обычно неглубоко расщеплены многочисленными складками.
Сходный тип расчленения средней кишки на небольшой центральный отдел и длинные трубчатые дивертикулы со сравнительно слабым вторичным ветвлением имеется у О. verrucosus, О. tartakovskyi, О. alactagalis, О, coniceps и О. moubata. Для этих видов характерно также подгибание пятой пары отростков на брюшную сторону. Видовые отличия связаны со степенью слияния оснований отростков и их вторичным ветвлением. У Ornithodoros tartakovskyi, О. alactagalis и О. coniceps срединный отросток дает недалеко от основания короткий вторичный дивертикул (рис. 73 и 74). Для О. coniceps характерна также значительная редукция 6-й пары отростков.
Иной тип расчленения мы наблюдаем у Alveonasus lahorensis. От массивного мешковидного центрального желудка у него отходят 4 пары коротких и толстых переднебоковых отростков и 2 пары заднебоковых. От 5-й пары остались лишь зачаточные выросты (рис. 69, 70).
Для видов рода Argas, отличающихся сильно уплощенным телом с краевым рантом, характерно сплющивание кишечника и очень сильное вторичное расщепление лопастевидных боковых отростков. Центральный отдел обычно развит слабо. У A. persicus еще можно различить 7 пар исходных стволов (рис. 71, 72). У A. vespertilionis дивертикулы расщепляются вторично до самого основания, а желудок очень сильно укорочен (рис. 75).
Личинки и нимфы всех изученных видов по характеру расчленения кишечника в основном сходны со взрослыми. Для них типичны лишь более сильное развитие центральной части, меньшая длина дивертикулов и часто захождение их в конечности (рис. 67, 69, 71).
У представителей сем Ixodidae в большинстве случаев также имеются 7 пар первичных отростков, из которых 1-я пара может быть недоразвита. Желудок у иксодид обычно значительно короче и тоньше, а боковые дивертикулы длиннее и реже подвергаются вторичному расщеплению. Кроме того, у дивертикулов наблюдается тенденция к заворачиванию на брюшную сторону с образованием по ходу значительных изгибов (рис. 59). Типичным строением средней кишки обладает клещ Нуаіотша asiaticum. У самок этого вида (рис. 83) 1—5-я пары передне-боковых отростков отходят общими основаниями от передних углов желудка. Из них 1-я и 3-я пары отростков следуют в переднем направлении, а 4-я и 5-я пары отходят каудально. Особенно сильно развитая 5-я пара отростков доходит почти до заднего конца тела, а затем подгибается на вентральную сторону и возвращается вперед до основания гнатосомы. 6-я и 7-я пары заднебоковых отростков также отходят общими основаниями. Из них сильнее развита 7-я пара, вентральная ветвь которой также почти доходит до гнатосомы.
У самцов Н. asiaticum, как и у большинства других АтЫуоттіпае, расчленение кишечника то же, что и у самок, но отростки значительно тоньше и короче (рис. 84). У личинок и нимф желудок массивнее, чем у половозрелой фазы, и от него отходят более короткие и толстые отростки. У видов рода Boophilus наблюдается тенденция ко вторичному расщеплению переднебоковых отростков и захождение их концов внутрь конечностей (рис. 81). Последняя особенность встречается довольно часто у неполовозрелых фаз и самцов рода Нуаіотша.
У самцов Ixodinae, в частности у Ixodes ricinus (рис. 77), наблюдается редукция части отростков средней кишки, так что от вытянутого трубчатого желудка отходят лишь две пары поперечных отростков, передняя из которых заканчивается двумя вертикальными ответвлениями, вероятно, гомологичными отросткам 2—3-й пар.
Желудок и дивертикулы имеют сходное гистологическое строение, и стенки их построены из пищеварительных, секреторных и недифференцированных клеток. Их тонкое строение и функциональная активность рассматриваются в разделе «Пищеварение» (см. стр. 161).
Тонкая кишка. Многие исследователи считали, что у иксодовых клещей тонкая кишка представлена слепым клеточным тяжем, идущим от средней кишки к ректальному пузырю и лишь в редких случаях имеющим
внутренний капиллярный просвет, так что сообщение между средней кишкой и анальным отверстием отсутствует, равно как и выделение фекалий (Вагнер, 1894; Bonnet, 1907; Mudrow, 1932). Это мнение возникло,
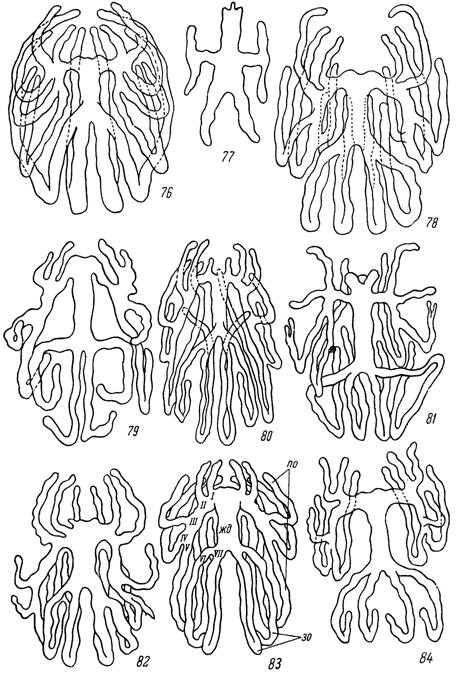
Рис. 76—84. Расчленение средней кишки у иксодовых клещей. 76 — самка и 77 — самец Ixodes ricinus; 78 — самка Derma- centor pictus\ 79 — самец и 80 — самка Нaemaphysalis sulcata; 81 — самка Boophilus calcaratus; 82 — самка Rhipicephalus
turanicus; 83 — самка и 84 — самец Hyalomma asiaticum.
жд — желудок; отростки средней кишки: по — переднебоковые, зо — заднебоковые; I—VII — порядковые номера боковых отростков.
по-видимому, при работе исключительно с голодными клещами, у которых стенки тонкой кишки действительно довольно часто спадаются, а акты дефекации сравнительно редки.
Большинство исследователей, основываясь на присутствии в испражнениях клещей продуктов переваривания крови, признавало существование сообщения между средней кишкой и ректальным пузырем, а в ряде работ приводились доказательства существования анатомически полно-анж

Рис. 85—93. Коксальные железы, сердце и ткани внутренней среды иксодоидных
клещей.
85 — сагиттальный разрез тела Alveonasus lahorensis на уровне коксального отверстия (по Сидорову, 1960в); 86 — отпрепарированная коксальная железа Ornithodoros moubata (вид сверху; по Lees, 1946b); 87 — кровеносная система Hyalomma asiati- сит; 88 — трофоцит, 89 — фагоцит и 90 — прогемоцит в капле гемолимфы самки Boophilus calcaratus (по Цвиленевой, 1961а); 91 — пленка аморфного вещества соединительной ткани с десмобластами и мышечными волокнами; 92 — участок тяжа жирового тела; 93 — эноцит, прикрепленный к тяжу жирового тела самки В. calcaratus
(по Цвиленевой, 19616).акж — акцессорная железа; вкп — внутренняя коксальная пора; мик — интеркоксальная мышца; мкж — мышцы коксальной железы; фкм — фильтрационная камера с системой каналов; сд — сердце; ар — аорта; пне — периневральный синус; сн — стволы к ногам; га — головная аорта.
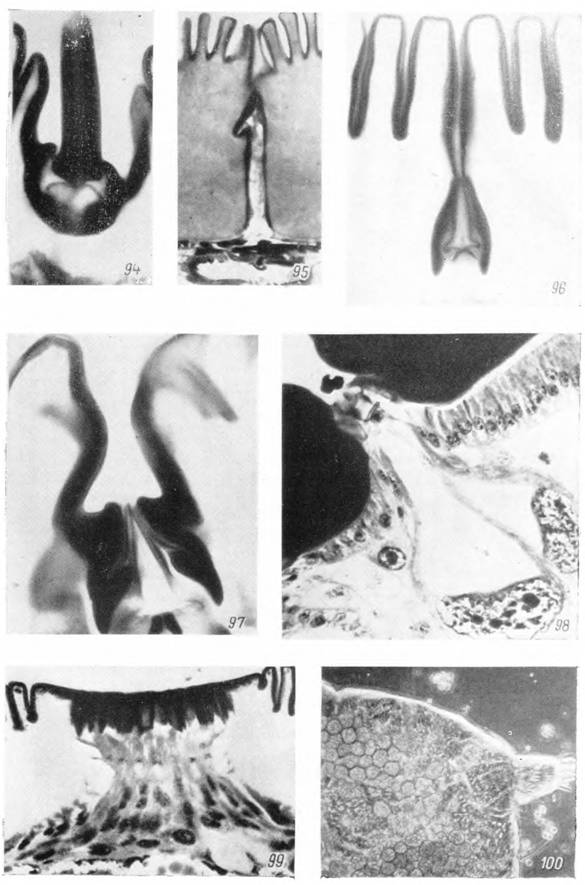
Рис. 94—100. Органы чувств самки — Hyalomma asiaticum.
94 — хета; 95 — аурикулярия; 96 — копьевидная сенсилла; 97 — стреловидная сенсилла голодной и 98 — питающейся особи; 99 — фовеа; 100 — пальпа самки перед освобождением от линочной шкурки. Микрофотографии: 94—97 — X 1200; 98, 99 — Х600; 100 — X 120.
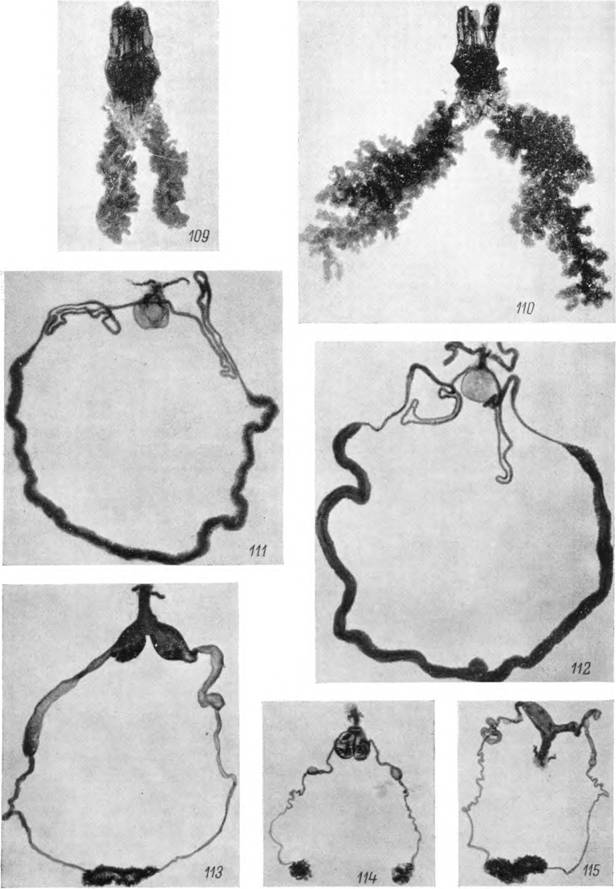
Рис. 109—115. Слюшіыс железы и женская половая система нксодоидных клещей. Отпрепарированные слюнные железы самки Hyalomma asiaticum: 109 — голодной, 110 — питающейся; 111 — половая система напитавшейся самки Н. anatolicum\ 112 — то же Dermacenlor pictus; 113 — половая система голодной самки Alveonasus lahorensis; 114 — то же Ar^as veapertiiionis; 115 — то же Argas persicus. Фотографии:
І09, ПО — /Л2\ 111—115 — XT.ценной тонкой кишки (Pagenstecher, 1861; Nordenskiold, 1908; Суворов, 1908; Roesler, *1934; Enigk u. Grittner, 1952). У всех исследованных нами видов иксодид мы также обнаружили нормально функционирующую тонкую кишку.
Существование тонкой кишки у аргасовых клещей вообще не вызывало сомнений и исключение составляет лишь Ornithodoros moubata, у которого, по наблюдениям В. Бургдорфера (Burgdorfer, 1951), постоянно отсутствует сообщение между средней кишкой и ректальным пузырем.
На основании анатомического изучения О. papillipes, О. verrucosus, О. tartakovskyi, Alveonasus lahorensis и Argas persicus мы установили, что у этих видов всегда имеется хорошо развитая тонкая кишка. Изучение строения Ornithodoros moubata подтвердило данные В. Бургдорфера об отсутствии сообщения между средней кишкой и ректальным пузырем у этого вида. Тонкая кишка у него заканчивается слепо и содержимое кишечника не может выводиться наружу в виде экскрементов.
Тонкая кишка представляет сравнительно короткую трубку, сужающуюся в направлении ректального пузыря. Она отходит медио-вентрально от заднего конца желудка перед его расщеплением на задне-боковые дивертикулы, опускается в задневентральном направлении и, немного не доходя до вентральной стенки тела, изгибается вверх и входит вместе с мальпигиевыми сосудами в передневентральную часть ректального пузыря (рис. 61). Диаметр тонкой кишки в значительной степени варьирует в зависимости от фазы развитая и стадии пищеварения. У личинок иксодиД она наиболее тонкая и по существу представляет клеточный тяж с очень узким внутренним просветом. У половозрелых особей она особенно хорошо видна во время питания вследствие заполнения ее полости кровью хозяина. В этом состоянии тонкая кишка мало отличается от концов дивертикулов средней кишки.
Являясь продолжением средней кишки, тонкая кишка и по гистологическому строению, особенно в передней части, очень сходна с первой. Ее стенки построены из эпителиальных клеток, лежащих на базальной мембране и окруженных сплетением продольных и кольцевых мышечных волокон. В зависимости от степени растяжения тонкой кишки эпителиальные клетки могут глубоко вдаваться апикальными концами в ее полость или же принимают кубическую и уплощенную форму. Цитоплазма клеток нормально свободна от пищевых включений, но иногда в ней обнаруживаются мелкие конкреции гематина. Последнее подтверждает потенциальную возможность клеток этой части кишечника принимать участие в процессах внутриклеточного пищеварения. В месте впадения тонкой кишки в ректальный пузырь кольцевая мускулатура образует уплотнение, напоминающее сфинктер. Он, возможно, контролирует выход содержимого средней кишки в ректальный пузырь.
Ректальный пузырь. Ректальный пузырь представляет непарный мешок, форма и размеры которого в значительной степени зависят от заполнения гуанином и фекалиями (рис. 61). У аргасовых клещей он расщепляется сзади на две лопасти. Слабо заполненный гуанином и экскрементами, он сохраняет почти шаровидную форму, и задние лопасти плохо различимы, тогда как при сильном растяжении последние выдаются далеко назад. Стенки ректального пузыря построены из плоского эпителия с крупными ядрами и бедной включениями плазмой. Снаружи они окружены базальной мембраной с сетью тончайших мышечных волоконец. В зависимости от степени заполнения пузыря форма его клеток меняется от почти кубической до листовидной, причем в последнем случае клеточные границы становятся неразличимыми (рис. 63).
IПрямая кишка. Медио-вентрально пузырь открывается в прямую кишку. Она опускается почти под прямым углом вниз и открывается
анальным отверстием (рис. 62). На всем протяжении она выстлана довольно толстой, сильно складчатой кутикулой. Границы отдельных эпителиальных клеток неразличимы. У иксодид она может сильно растягиваться фекалиями или экскретами, которые затем с силой выталкиваются из анального отверстия при сокращении мускулатуры стенок кишки.
Анальное отверстие представляет продольную щель, прорезающую анальную пластинку. Последняя состоит из двух створок и окружена так называемым анальным кольцом — участком склеротизованной кутикулы. Между створками находится эластичная сочленяющая мембрана. От стенок прямой кишки к анальной пластинке идет пара мышечных пучков. При их сокращении анальная щель раскрывается, а нижняя часть прямой кишки увеличивается в объеме и немного выворачивается наружу, облегчая дефекацию. При расслаблении мускулатуры анальная щель закрыта, а просвет прямой кишки почти исчезает из-за спадения стенок.
Еще по теме Иксодовые клещи.:
- Иксодовые клещи (сем. Ixodidae).
- Балашов Ю.С.. Иксодовые клещи — паразиты и переносчики инфекций. — СПб.: Наука,1998. — 287 с., 1998
- ИКСОДОВЫЕ КЛЕЩИ (Ixodidae)
- Отряд Клещи
- Раздел IV КЛЕЩИ, ИМЕЮЩИЕ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
- Аргасовые клещи (сем. Argasidae).
- Отряд Паразитоформные клещи - Parasitiformes.
- КЛЕЩИ-КРАСНОТЕЛКИ (TrombicuIidae)
- Отряд Акариформные клещи - Acariformes
- Глава IX ИКСОДОВЫЕ КЛЕЩИ И ВОЗБУДИТЕЛИ ИНФЕКЦИЙ