ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАЗМНОЖЕНИЯУ САМОК
Развитие женской половой системы. В период эмбрионального развития яичник закладывается в виде парного скопления клеток. Эти зачатки вытягиваются назад и сливаются позади ректального пузыря (Вагнер, 1894).
У только что вылупившихся личинок Нуаіотта asiaticum, Ixodes ricinus и Ornithodoros papillipes яичник был всегда представлен непарным зачатком из группы вытянутых в поперечном направлении клеток, тесно прилегающих к задней стенке ректального пузыря. У голодных и напитавшихся личинок в зачатке различимы более крупные, почти сферические или неправильно многоугольной формы оогонии с пузыревидными ядрами (рис. 395). Между ними лежат многочисленные более мелкие ядра эпителиальных клеток. Границы между самими клетками неразличимы. У личинок эпителиальные клетки не только вклиниваются между оого- ниями, но и могут окружать их снаружи. Снаружи яичник покрыт тонкой соединительнотканной оболочкой с одиночными, сильно уплощенными ядрами.Эктодермальные зачатки выводных путей у голодных личинок еще отсутствуют. Стимулом к их образованию, так же как и к росту яичника, служит питание. У иксодид эти процессы начинаются уже на стадии кровососания, а у быстро насасывающихся аргазид — на стадии линьки. В первые дни линьки после отделения гиподермы от старой кутикулы позади нервной массы на вентральной стороне тела гиподермальные клетки выпячиваются внутрь в заднедорсальном направлении и формируют зачаток влагалища (рис. 383). За время линьки влагалище приобретает форму конуса, суженный конец которого вклинивается между гиподермальными клетками (рис. 384).
Митотические деления многочисленны на этой стадии линьки также среди оогониев и эпителиальных клеток яичника. Размеры яичника заметно увеличиваются, а у иксодовых клещей концы его заворачивают вперед и он приобретает подковообразную форму. От концов яичника в сторону зачатка влагалища вытягиваются очень тонкие клеточные тяжи диаметром 3—7 мк.
Слияние их с эктодермальным влагалищем происходит в самом конце линьки личинок, а непарная матка формируется уже на нимфальной фазе. Полость внутри яйцеводов неразличима, а клеточные границы их стенок не видны (рис. 385).У голодных нимф Нуаіотта asiaticum и Ixodes ricinus яичник подковообразный, а его концы достигают уровня IV пары кокс. В поперечном сечении он округлый, причем местами уже различим щелевидный центральный просвет. Последний окружен многочисленными ядрами эпителиальных клеток, а по периферии лежат более крупные оогонии. От концов яичника отходят тонкие и почти прямые парные яйцеводы, соединяющие его с задне-боковыми концами зачатка влагалища. Последний уже явственно изгибается параллельно главной оси тела и сплющен дорсо- вентрально. Внутри зачатка образуется небольшая полость (рис. 386).
Стимулом для дальнейшего развития половой системы у нимф служит кровососание. В клетках возобновляются митотические деления и размеры закладок значительно увеличиваются. У I. ricinus задний конец влагалища продолжает вытягиваться каудально и одновременно сильно утолщается. Соприкасающиеся с ним концы яйцеводов сливаются между собой и образуют короткую непарную трубку — будущую матку. Длина яйцеводов значительно увеличивается и они образуют характерные петлеобразные изгибы. В матку вдается задний конец влагалища, но полости этих органов остаются разделенными клеточной перегородкой. Последняя исчезает лишь в конце линьки, когда устанавливается сообщение между полостями матки и влагалища.
У Hyalomma asiaticum задний конец зачатка влагалища еще во время кровососания расщепляется на вентральную и дорсальную половины (рис. 387). Первая из них в дальнейшем превращается в цервикальный отдел и соединительную трубку. Дорсальный мешок представляет зачаток семе приемника. Он сильно разрастается в задневентральном направлении, сохраняя суженный участок только в месте отхождения от влагалища. Стенки дорсального мешка образуют многочисленные складки. Одновременно с образованием семеприемника и соединительной трубки в месте горизонтального изгиба влагалища с дорсальной стороны образуются два вытянутых выроста — будущие трубчатые придаточные железы.
Во время питания и в первую половину линьки преобладают процессы клеточных делений и роста закладок выводных путей.
Границы эктодермальных клеток на этой стадии неразличимы, а ядра лежат на разных уровнях, создавая впечатление многослойности (рис. 389). Строение клеток во всех частях закладки совершенно идентично.Во вторую половину линьки число митозов быстро уменьшается, ядра располагаются базально в один слой и между ними появляются клеточные границы. Обнаруживаются первые признаки гистологической дифференцировки отделов влагалища и формирования кутикулярной выстилки. Высота клеток при этом значительно увеличивается и они становятся столбчатыми. Ядра перемещаются к центру, основания клеток заполнены очень крупными прозрачными вакуолями, тогда как апикальные концы сильно базофильны. Вакуоли заполнены гликогеном, а базо- филия цитоплазмы обусловлена высоким содержанием РНК (рис. 390).
Первой откладывается очень тонкий кутикулиновый слой эпикутикулы, а затем прокутикула. Высота клеток постепенно уменьшается и ядра вновь занимают базальное положение. В вестибулярном отделе поверхность цитоплазмы образует правильные ряды выростов. Откладывающаяся на них эпикутикула сразу же образует систему вертикальных, тесно прилегающих друг к другу складок. Под ними располагаются горизонтальные слои прокутикулы (рис. 391). Сходным образом формируются кутикулярные складки цервикального отдела и передней стенки семеприемника; поверхность эпикутикулы соответствует первоначальным выростам цитоплазмы, а нижние слои прокутикулы лежат горизонтально (рис. 392). На большей части семеприемника и в соединительной трубке отложению кутикулы предшествует образование глубоких гиподермаль- ных складок, содержащих ядра. На их поверхности также появляются многочисленные цитоплазматические выросты. В результате кутикулярная выстилка сразу же приобретает очень сложную конфигурацию и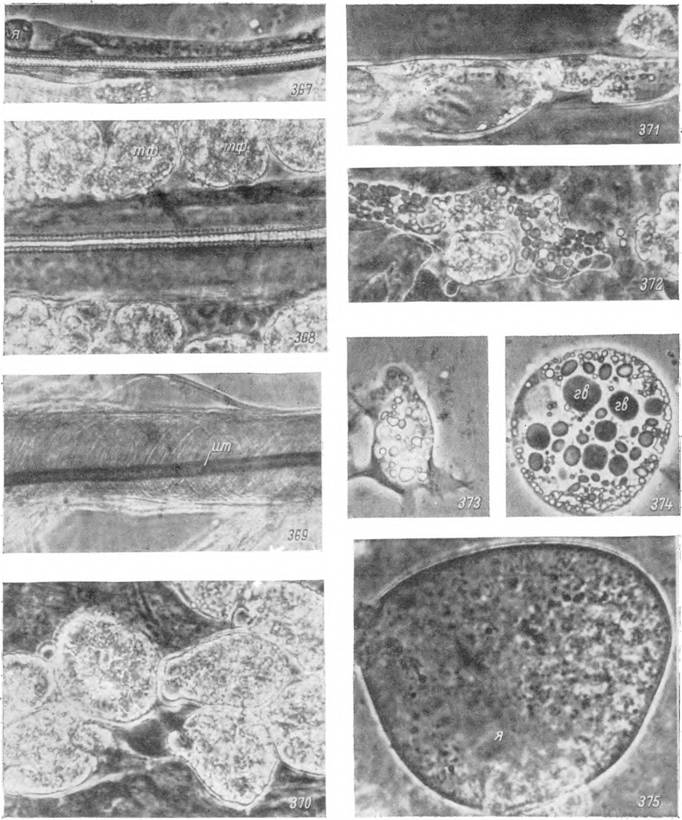
Рис. 367—375. Прижизненные микрофотографии (фазовый контраст) трахейных стволов и тканей внутренней среды нимф Нуаіотта asiaticum на стадии линьки. Х500.
367 — трахейный ствол на 1-е сутки после питания; 368 — то же на 5-е сутки; 369 — то же на 10-е сутки; 370 — группа склеивающихся трофоцитов; 371 — тяж жирового тела в конце линьки; 372 — растворение трофоцита; 373 — фагоцит; 374 — трофоцит с гликопротеиновыми
включениями; 375 — эноцит.
я — ядра; гв — гликопротеиновые включения; ит — отслоившаяся интима трахей нимфальной фазы; тф — слой
трофоцитов, прилегающих к трахейному стволу.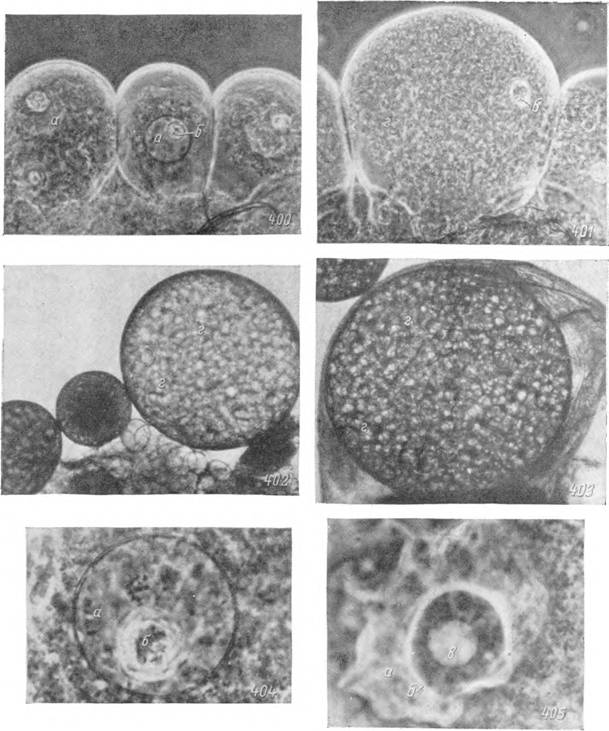
Рис.
400—405. Микрофотографии ооцитов самки Argas persicus.400 — II стадия; 401 — III стадия; 402 — IV стадия; 403 — V стадия; 404 — ядро ооцита II стадии: 405 — ядро ооцита III стадии. Фазовый контраст: 400, 401 — Х250, 402, 40? — Х40 и 404, 405 — Х800.
а — ядро; б — нуклеола; в — вакуоль; г — желточные шары.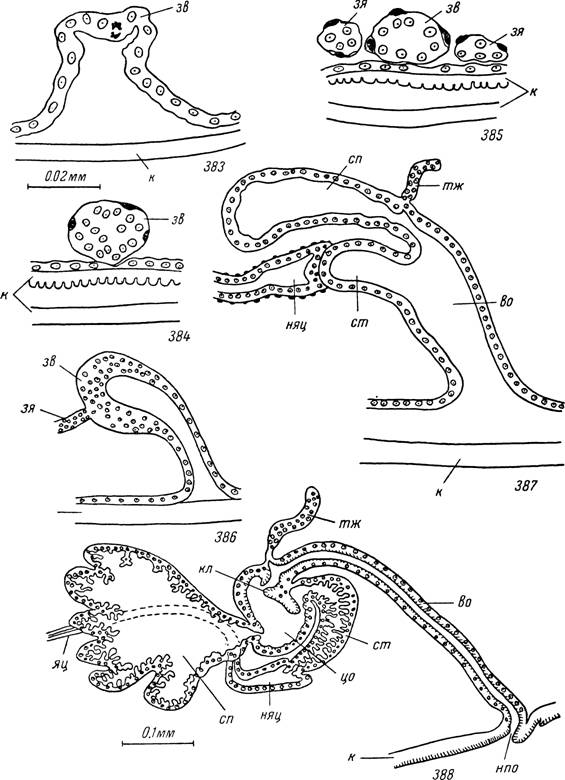
Рис. 383—388. Развитие женской половой системы Hyalomma asiaticum.
383 — личинка в начале линьки; 384 — личинка в середине линьки; 385 — личинка в конце линьки; 386 — голодная нимфа; 387 — линяющая нимфа; 388 —
голодная самка.
зв — зачаток влагалища и придаточных желез; зя — зачаток яйцеводов; во — вестибулярный и цо — цервикальный отделы влагалища; тж — трубчатая придаточная железа; cm — соединительная трубка; кл — клапан; сп — семеприемник; нпо — наружное половое отверстие; яц — яйцевод; к — кутикула; ияц — непарный яйцевод.
в состав ее складок входят прокутикула и подстилающие ее гиподермальные клетки с ядрами (рис. 393).
В зачатках трубчатых придаточных желез клетки очень тесно прилегают друг к другу, границы между ними не заметны и объем цитоплазмы невелик. Деление ядер продолжается до конца линьки. Дифференцировка на крупные ядра железистых клеток и более мелкие синцитиального вещества еще отсутствует. Секреторная зернистость в цитоплазме не выявляется. В суженных протоках стенки состоят из нормального кубического эпителия, секретирующего тонкую кутикулу.
В образовании соединительнотканной оболочки половой системы важная роль принадлежит трофоцитам гемолимфы. Во время линьки они сплошным слоем окружают влагалище. В конце линьки значительная часть их растворяется и освобождающиеся гликопротеиновые гранулы* вероятно, используются для построения основного вещества наружных оболочек. Некоторые из трофоцитов в дальнейшем непосредственно превращаются в десмоциты (рис. 412). Внутри оболочки происходит дифференцировка пучков волокон кольцевой и продольной мускулатуры. Последние особенно сильно развиты в соединительной трубке и шейке семеприемника.
В закладке яичника с началом нимфального питания возобновляются митотические деления оогониев и эпителиальных клеток.
Со второй половины стадии линьки наряду с продолжающимся размножением половых элементов часть оогониев начинает превращаться в ооциты. У нимф их развитие ограничивается первыми стадиями малого роста (лептонема, зигонема и пахинема). У заканчивающих линьку особей в яичнике уже хорошо различим центральный щелевидный просвет, а на дорсальной поверхности благодаря вворачиванию стенки внутрь формируется дорсальная складка.У аргасовых клещей формирование органов размножения в основном сходно с иксодовыми, но органогенез растянут на несколько нимфальных стадий. Развитие половой системы у аргазид происходит только после кровососания во время линьки и прерывается в период голодания. У Огпі- thodoros papillipes основные отделы половой системы различимы уже у нимф II возраста, но для их окончательного развития требуется еще от 2 до 4 линек. У личинок и голодных нимф какие-либо зачатки органа Жене отсутствуют. Он формируется при линьке в половозрелую особь после отделения гиподермы от старой кутикулы. У напитавшихся нимф Нуаіотта asiaticum клетки стенок камеростомальной складки очень интенсивно делятся и впячиваются в полость тела в виде дорсо-вентрально уплощенного кармана. Первоначально толщина его, особенно в боковых частях, очень велика и достигает 100—140 мк. Границы клеток неразличимы, ядра лежат в несколько рядов. К моменту отложения кутикулы орган Жене достигает своих окончательных размеров и клеточные деления прекращаются. Клетки становятся столбчатыми, ядра лежат в один ряд. Базально цитоплазма содержит крупные прозрачные вакуоли, а апикально сильно базофильна. На ее поверхности откладывается тонкая и слоистая эндокутикула. Отделение гиподермы от кутикулы начинается от камеростомальной складки в сторону рогов и завершается перед окончанием линьки.
У голодных самок гиподермальные клетки уплощенной формы с небольшими овальными ядрами в 4—5 мк. В закладках будущих желез строение клеток то же, что и в остальных частях органа Жене (рис. 138). С началом кровососания клетки железистых выпячиваний начинают делиться и увеличиваться в размерах, образуя трубчатые железы (рис.
139). У закончивших питание Нуаіотта asiaticum железы достигают диаметра 60—100 мк (рис. 140), а у Ixodes ricinus — 100—120 мк. К на- чалу яйцекладки железы утолщаются соответственно до 120—150 и 200 мк (рис. 141). Клетки становятся столбчатыми и достигают высоты 50—70 мк. Ядра увеличиваются в несколько раз и диаметр их достигает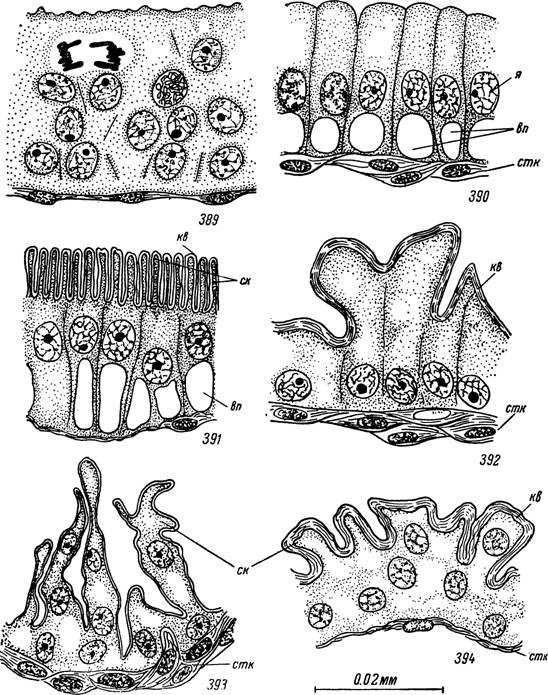
Рис. 389—394. Гистогенез женской половой системы Hyalomma asialicum. 389 — эпителиальные клетки зачатка влагалища в конце пимфального питания; 390 — то же в начале линьки перед отложением кутикулы; образование кутикулярной выстилки: 391 — вестибулярного отдела влагалища, 392 — цервикального отдела влагалища и 393 — соединительной трубки; 394 — стенка семеприемника голодной самки.
я — ядра клеток зачатка влагалища; ск — складки апикальной поверхности клеток; стк — соединительнотканные клетки; кв— кутикулярная выстилка; вп — вакуоли с полисахаридом.
25—30 мк. Базальная часть клеток содержит волокнистые пиронинофиль- ные структуры, а апикальная заполнена мелкими эозинофильными вакуолями, дающими интенсивное окрашивание на белки. Просвет желез увеличивается до 20—30 мк и содержит эозинофильный секрет. Он отделен от клеток тонкой, сильно светопреломляющей оболочкой, слегка окрашивающейся железным гематоксилином. Местами она отделяется от клеточной поверхности (рис. 417). Железистые клетки в процессе секреции не разрушаются, а лишь сморщиваются, теряют эозинофильные вакуоли, а ядра начинают интенсивно прокрашиваться. Строение клеток протоков желез и стенок органа Жене не меняется.
У аргасовых клещей орган Жене развивается только у нимф последнего возраста при линьке в самок. У взрослых особей железистая ткань в задней части рогов проявляет признаки секреторной активности только перед яйцекладкой. Высота клеток увеличивается до 50—60 мк и в их апикальных концах появляются многочисленные эозинофильные вакуоли. После окончания яйцекладки высота клеток быстро уменьшается и секреторные вакуоли исчезают до следующего гонотрофического цикла. Ядра железистых клеток постоянно крупнее, чем в гиподерме стенок мешка, а у голодных или яйцекладущих особей размеры их существенно не отличаются.
Строение яичника, оогенез и оплодотворение. На поперечном разрезе яичник голодной самки овальный или почти круглый. Снаружи он окружен тонкой соединительнотканной оболочкой — tunica propria. Оболочка построена преимущественно из основного вещества с тончайшими пересекающимися волокнами и представляет неклеточное образование. Внутри яичника по всей его длине проходит щелевидный просвет, сообщающийся с полостями парных яйцеводов и выстланный слоем эпителиальных клеток. Границы между отдельными эпителиальными клетками неразличимы, а ядра их мельче, чем таковые ооцитов (рис. 396). Пузыревидные ядра молодых ооцитов, исключая область дорсальной складки, занимают периферийное положение и окружены сравнительно узкими цитоплазматическими ободками с хорошо различимыми клеточными границами.
На основании цитологического изучения особенностей оогенеза клещей Argaspersicus, Hyalomma asiaticum и Ixodes ricinus мы считаем возможным различать пять стадий роста ооцитов.
I стадия. Ооциты, образующиеся при размножении оогониев, минуя период интеркинеза, сразу же переходят в профазу первого деления созревания (рис. 396). Первая стадия, начинающаяся с прелептонемы мейоза и заканчивающаяся растворением хромосом в ядре, точно соответствует периоду малого роста ооцитов I порядка. Размеры последних за это время обычно увеличиваются очень незначительно. Диаметр их достигает у Argas persicus 12—15 мк и у Ixodes ricinus 10—14 мк, а диаметр ядер соответственно 8—10 и 7—11 мк. Цитоплазма слабо базофильна и бедна РНК. Внутри ядра отсутствует рибонуклеиновое ядрышко. В начале этой стадии в ядре Argas persicus появляются Фельген-положительные глыбки хроматина (стадия пролептонемы). Образовавшиеся прохромосомы начинают деспирализироваться и образуют клубок нитей, заполняющих ядро (стадия лептонемы). После конъюгации хромосом происходит утолщение нитей (стадия пахинемы), но в дальнейшем в начале стадии диплонемы хромосомы теряют способность к окраске и их не удается больше выявить внутри ядра. По данным Ю. Л. Горощенко (1960), во времени растворения хромосом в ядре между изученными видами клещей наблюдаются определенные различия, но этот процесс никогда не происходит позднее начала диакинеза. Ооциты I стадии всегда заключены внутри яичника.
II стадия. Период с начала большого роста и до появления первых желточных гранул в цитоплазме ооцита мы называем II стадией (рис. 400 на стр. 217). К концу ее размеры ооцитов увеличиваются у Argas persicus до 60—100 мк и Ixodes ricinus до 60—80 мк, а ядер до 20—25 и 25—30 мк. Плазма почти прозрачная и не содержит включений. Она хорошо прокрашивается различными гистологическими красителями и содержит много
РНК. Ядрышко обычно сферической формы и относительно гомогенно. Размеры его быстро увеличиваются и к концу этой стадии оно достигает диаметра в 6—12 мк. Биваленты быстро разрыхляются и исчезают, ядро вы-
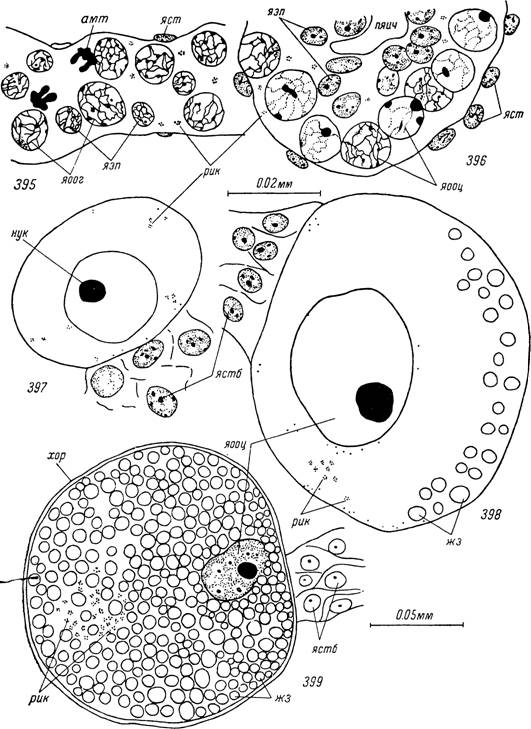
Рис. 395—399. Оогенез и распределение риккетсий Wolbachia sp. в оого- ниях и ооцитах Нуаіопгта asiaticum.
395 — зачаток яичника у недавно напитавшейся личинки; 396 — участок яичника голодной самки; 397 — ооцит питающейся самки на II стадии развития; 398 — то же в начале III стадии и 399 — в конце III
стадии развития.
рик — риккетсии, яст — ядра соединительнотканной оболочки стенки яичника; яэп — ядра эпителиальных клеток яичника; яоог — ядра оогониев; яооц — ядра ооцитов; ясгпб — ядра стебелька; пук — нуклеола; пяич — полость яичника; жз — желточные зерна; хор — «хорион»; амтп — анафаза митоза.
глядит оптически пустым и не окрашивается по Фельгену (рис. 404). В цитоплазме подобных ооцитов постоянно присутствует Фельген-положительная зернистость. На определенных стадиях из нее образуются характерные
сферические или грибовидные скопления (рис. 397). По данным электронной микроскопии (Roshdy, 1961b), Фельген-положительная зернистость не связана с ядром и представляет группы из симбиотических риккет- ский, а не частицы вышедшей из ядра ДНК, как предполагали некоторые исследователи (Wagner-Jevseenko, 1958 и др.). К концу второй стадии ооциты выпячиваются над поверхностью яичника.
III стадия. Третья стадия начинается с появлением первых желточных включений в наружной зоне цитоплазмы (рис. 398) и заканчивается с заполнением крупными желточными шарами всего ооцита (рис. 399, 401). Его размеры увеличиваются у Argas persicus до 150—200 мк и Ixodes ricinus — 200—250 мк, а ядер до 25—30 и 40—45 мк. В цитоплазме вокруг ядра образуются гранулы желтка диаметром в 2—5 мк, постепенно распространяющиеся к периферии. В цитоплазме по-прежнему сохраняется высокая концентрация РНК. Ядра достигают своих максимальных размеров и образуют боковые лопасти и выросты (рис. 405). В ядрышке, достигающем диаметра 15—20 мк, хорошо видны одна или несколько вакуолей. Иногда от него отпочковываются небольшие фрагменты, растворяющиеся в кариолимфе. ДНК в ядре не выявляется, а в цитоплазме сохраняется Фельген-положительная зернистость. Вокруг ооцита начинает формироваться яйцевая оболочка, достигающая полного развития на IV стадии. Это плотная и гомогенная двухконтурная структура, лишенная какого-либо отверстия, сходного с микропиле. Она образуется самим ооцитом и называть ее, по аналогии с насекомыми, хорионом можно лишь с оговорками, помня об их различном происхождении.
IV стадия. Для четвертой стадии характерно завершение отложения желтка в виде крупных шаров диаметром в 20—30 мк. Они полностью маскируют ядро, которое к этому времени приобретает неправильно угловатую форму и перемещается в периферийную зону ооцита. Крупное ядрышко и ядерная оболочка на этой стадии растворяются. Сохраняющиеся диакинетические биваленты образуют одно компактное тело, называемое кариосферой (Горощенко, 1965), в котором неразличимы отдельные хромосомы. Кариосфера окружена узкой цитоплазматической зоной и сохраняется на протяжении всего дальнейшего роста ооцита вплоть до оплодотворения. Цитоплазма в ооцитах IV стадии представлена в виде тонкой сети между глыбами желтка и относительно бедна РНК (рис. 402). На этой стадии заканчивается рост ооцитов, которые достигают своих окончательных размеров в 600—800 мк у Argas persicus или 700 мк у Ixodes ricinus, выдаются в полость тела и связаны со стенкой яичника стебельком из эпителиальных клеток.
V стадия. Зрелые яйца, потерявшие связь со стенками яичника и находящиеся в его просвете или в яйцеводах, мы относим к пятой стадии. Овуляция, по-видимому, связана с усиливающимся по мере роста яйцеклеток натяжением tunica propria, так что под его воздействием зрелый ооцит раздвигает клетки стебелька и выпадает в полость яичника. Размеры и внешний вид ооцитов после овуляции не меняются и их дальнейшее развитие связано с завершением делений созревания и оплодотворением (рис. 403).
Предложенная схема позволяет объективно оценивать состояние яичника по внешним признакам, строго соответствующим цитологическим изменениям в развивающейся яйцеклетке. I стадия совпадает с периодом малого роста, II стадия — с цитоплазматической фазой большого роста и III и IV — отвечают периоду отложения желтка (трофоплазматическая фаза), часто называемому третьим периодом роста ооцита. После окончания профазы мейоза и образования кариосферы наблюдается остановка в подготовке к I делению созревания, которое возможно только после окончания роста ооцита и, по данным Ю. Л. Горощенко (1965), стимулируется процессом оплодотворения. Таким образом, весь период большого роста и отложения желтка протекает в ооцитах первого порядка, а превращение их в ооциты второго порядка происходит уже во вполне сформированных яйцах.
Процесс оплодотворения у клещей до последнего времени оставался неизвестным и был впервые описан Ю. Л. Горощенко (1965) у аргасовых клещей. Оплодотворение происходит в ампуловидных расширениях яйцеводов, где скапливаются зрелые спермин — спермиофоры, проникающие сюда из семеприемников. После поступления яйца в ампуловидное расширение яйцевода его толстая наружная оболочка («хорион») растворяется спермиофором, но внутрь ооцита входит только маленькое ядро спермия. Остающиеся трубчатые части спермиофора не участвуют в оплодотворении и растворяются железистым эпителием стенок яйцевода и матки. После оплодотворения мужской пронуклеус занимает в ооците строго центральное положение и стимулирует возобновление хода I деления созревания ооцита. II деление созревания и слияние мужского и женского пронуклеусов происходило уже в отложенных яйцах.
У иксодовых клещей сцециализированные ампуловидные расширения яйцеводов отсутствуют, но спермиофоры также скапливаются лишь в передних третях яйцеводов. Оплодотворение ооцитов Hyalomma asiaticum происходит только после их овуляции и поступления в передние части яйцеводов. Весь дальнейший ход делений созревания и слияния пронуклеусов сходен с описанным для аргасовых клещей.
Яичники недавно перелинявших самок аргазид содержат оогонии и ооциты I и II стадий. На этом их рост обычно приостанавливается и возобновляется лишь после имагинального кровососания. У видов, обладающих способностью к автогенному развитию яиц, последние без задержки заканчивают свой рост за счет нимфального питания. После окончания яйцекладки незакончившие развития ооциты III—IV стадий дегенерируют и яичник вновь содержит одни оогонии и молодые ооциты. Следующая волна размножения оогониев и роста ооцитов обычно наступает только после нового питания.
У иксодовых клещей на стадии послелиночного доразвития в яичнике еще встречаются митотические деления эпителиальных клеток и малый рост ооцитов I порядка. Однако мейотические изменения не заходят дальше диакинеза и приостанавливаются до начала кровососания. Исчезновение митозов среди эпителиальных клеток стенок по времени совпадает с окончанием послелиночного доразвития.
У голодных самок ооциты, лежащие по периферии яичника, крупнее и находятся на более поздних стадиях мейоза (преимущественно диакинез и в меньшей степени диплонема), тогда как внутри дорсальной складки они отстают в развитии (не старше диплонемы) и среди них встречаются даже оогонии. У видов рода Ixodes значительная часть ооцитов на стадии послелиночного доразвития успевает закончить период малого роста. Они останавливаются в развитии в начале цитоплазматической стадии большого роста и слегка выступают над поверхностью яичника. У изученных представителей АтЫуоттіпае ооциты обычно не приступают к цитоплазматическому росту и поверхность яичника у голодных клещей бывает лишь слегка волнистой или даже гладкой (рис. 396).
Стимулом к возобновлению процессов оогенеза служит начало кровососания. Ооциты быстро заканчивают цитоплазматическую фазу большого роста и в них начинается отложение желтка (трофоплазматическая фаза). Благодаря очень значительному увеличению в размерах ооциты растягивают тонкую tunica propria и выдаются над поверхностью яичника. У иксодид ооциты дорсальной складки яичника развиваются медленнее, чем на вентральной и боковых стенках, в результате чего они оказываются
как бы погруженными внутрь полости органа (рис. 127). У аргазид наиболее быстро развиваются яйцеклетки, лежащие ближе к задней стенке
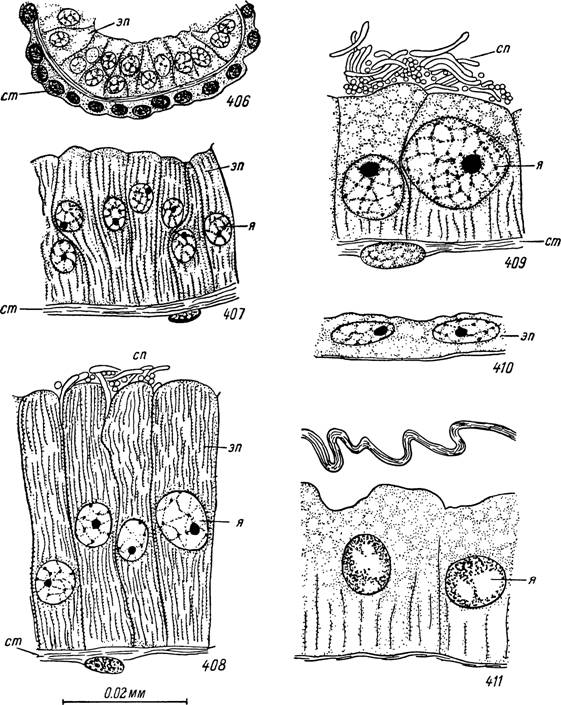
Рис. 406—411. Строение яйцеводов и семеприемника самки Hyalomma asiaticum. 406 — стенка яйцевода голодного клеща; 407 — проксимальная часть яйцевода в конце питания; 408 — дистальный участок яйцевода, почти свободный от спер- миев; 409 — то же, растянутый спермиями; 410 — стенка яйцевода во время яйцекладки; 411 — стенка семеприемника напитавшейся особи.
эп — эпителиальные клетки яйцевода; cm — соединительнотканная оболочка; я — ядра;
сп — сперми офоры.
яичника. С разновременностью развития яйцеклеток связана большая растянутость яйцекладки, особенно у иксодовых клещей.
Среди эпителиальных клеток стенок яичника в этот период наблюдаются многочисленные митотические деления, прекращающиеся после отпадения с хозяина. Выдающиеся в полость тела ооциты соединены со стенкой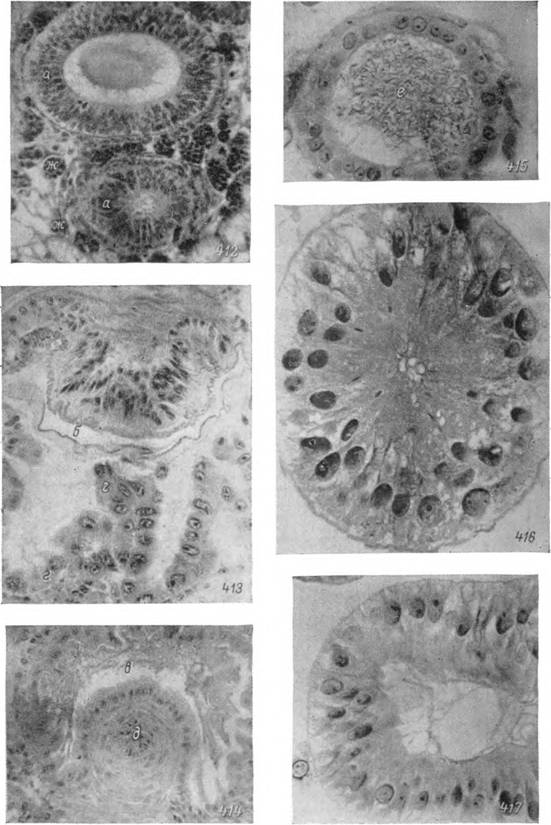
Рис. 412—417. Женская половая система напитавшейся нимфы Hyalomma asiaticum (412) и самки Ixodes ricinus перед началом Яйцекладки (41S—417). 412 — закладка семеприемника и соединительной трубки; 413 — вестибулярный отдел влагалища; 414 — впадение соединительной трубки в цервикальный отдел; 415 — дистальный участок яйцевода; 416 — трубчатая придаточная железа; 417 — участок железы органа Жене. Микрофотографии: 412—414 — X 125, 415—417 — Х250.
а — недифференцированные гиподермальные клетки полового зачатка; б — вестибулярный отдел влагалища; в — цервикальный отдел влагалища; г — клетки лопастевидной придаточной железы; д — соединительная трубка; е — спермин; ж — трофоциты.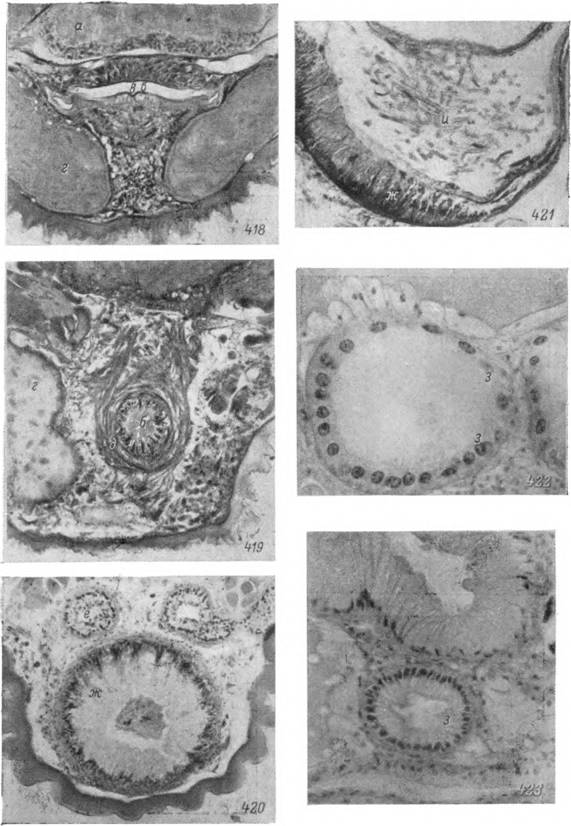
Рис. 418—423. Половая система самки Ornithodo.ros pdpillipes.
418 — вестибулярный отдел влагалища; 419. — цервикальный отдел влагалища; 420 — шейка матки и трубчатые придаточные железы; 421 — участок рога матки; 422 — амупловидное расширение яйцевода; 423 — проксимальная часть яйцевода. Микрофотографии: 418—421 — X 125, 422, 423 — Х250.
а — нервный ганглий; б — полость влагалища; в — кутикулярная выстилка влагалища; г — дивертикулы кишечника; д — мускулатура влагалища; е — трубчатая придаточная железа; ж — железистые клетки стенки матки; э — эпителиальные клетки стенок яйцеводов;
и — спермиофоры.яичника стебельком или ножкой из эпителиальных клеток. Клетки ножки увеличиваются в размерах и претерпевают определенные циклические изменения, возможно, связанные с секрецией или передачей веществ растущей яйцеклетке. Ядра этих клеток также увеличиваются, а в цитоплазме появляются мелкие базофильные включения (рис. 397). Эти клетки, возможно, выполняют часть функций фолликулярных или трофических клеток насекомых, тогда как соединительнотканная неклеточная оболочка, окружающая яичник снаружи, может осуществлять лишь опорную и, в меньшей степени, барьерную функции. Клетки, выстилающие просвет яичника при подготовке к овуляции, вероятно, несут секреторную функцию. В их цитоплазме появляется значительное количество слабо базо- фильных зернистых включений, встречающихся и в просвете яичника.
В яичниках закончивших яйцекладку и погибающих самок иксодовых клещей всегда сохраняются отдельные ооциты I и II стадий, не приступавшие к росту, а также не закончившие его и сморщившиеся ооциты
III— V стадий. Они лежат в массе дегенерирующих эпителиальных клеток, окруженных спавшейся tunica propria. У аргазид не закончившие развития ооциты III—IV стадий также дегенерируют и яичник, как и у молодых особей, вновь содержит одни ооциты I—II стадий и оогонии. Новый период роста ооцитов, так же как и размножения эпителиальных клеток, начинается только после следующего кровососания.
Выводные пути, придаточные железы и яйцекладка. У голодных самок иксодид яйцеводы представляют тонкие трубки, слегка утолщающиеся к дистальным концам. У Hyalomma asiaticum диаметр их 25—30 мк, а в расширенных окончаниях до 40—50 мк. У Ixodes ricinus — соответственно 15—18 и 25—30 мк. На поперечном разрезе они почти круглые и небольшой центральный просвет различим лишь в дистальной части. Небольшие овальные ядра эпителиальных клеток, диаметром в 5—7 мк, образуют сплошной базальный слой. Снаружи яйцеводы окружены соединительнотканной оболочкой. Ее ядра располагаются по самой периферии ж очень тесно прилегают друг к другу (рис. 123 и 406).
С первых дней кровососания наблюдаются многочисленные деления ядер эпителиальных клеток, размеры которых очень сильно увеличиваются. Различия в строении дистальных и проксимальных участков при этом усиливаются. Так, у Hyalomma asiaticum диаметр первых достигает к концу питания 200 мк при высоте клеток в 60—70 мк, в проксимальных частях диаметр обычно не превышает 100—150 мк и высота клеток 30— 60 мк. К началу яйцекладки диаметр растянутых спермиофорами дистальных участков может иногда увеличиваться до 400 мк (рис. 415),
Апикальные концы клеток дистальных отделов яйцеводов выдаются в просвет, а между ними внедряются многочисленные спермиофоры, вышедшие из сперматодоз (рис. 408). Клетки стенок яйцеводов на этой стадии приобретают железистый характер. Ядра их занимают центральное положение и сильно гипертрофированы (диаметр 16—22 мк). В базальной части клеток появляются волокнистые структуры, интенсивно красящиеся на РНК, а цитоплазма апикальных частей заполнена ШИК-положительной зернистостью.
Местами спермиофоры целиком заполняют полость яйцеводов, так что толщина их растянутых стенок уменьшается до 20—40 мк, а клетки заметно уплощаются (рис. 409). После внедрения ядер спермиев в проходящие по яйцеводам ооциты сохраняющиеся трубчатые части спермио- фор могут поглощаться эпителиальными клетками стенок. Подробное описание этого интересного явления впервые было дано И. И. Соколовым (1958). В иностранной литературе до последнего времени было распространено неправильное представление о томг что фагоцитоз спермиофор представлял особую форму оплодотворения и был связан с последующей диффузией вещества мужской гаметы в проходящие по яйцеводу ооциты (Geigy a. Wagner, 1957; Wagner-Jevseenko, 1958).
По направлению к яичнику высота эпителиальных клеток уменьшается, причем одновременно они становятся более узкими. Стенки яйцевода образуют продольные складки, так что клетки оказываются собранными в подушки, вдающиеся в просвет (рис. 407). Цитоплазма их слегка волокнистая, но признаки секреторной активности выражены слабее, чем в дистальных отделах.
В период яйцекладки сильно растягиваются и проксимальные отделы яйцеводов, так что их диаметр достигает 300 мк и более. Толщина их стенок при этом уменьшается до 10—12 мк (рис. 410). Внутри оболочки в продольном и кольцевом направлениях дифференцируются многочисленные гладкие мышечные волокна. Они обеспечивают непрерывные волны перистальтических сокращений, хорошо различимых на отпрепарованных переживающих органах. После окончания яйцекладки стенки яйцеводов спадаются, сморщиваются, ядра становятся пикнотическими и клетки быстро дегенерируют.
Деления или увеличения размеров ядер соединительнотканной оболочки яйцеводов во время кровососания не происходит. В связи с этим по мере утолщения яйцеводов расстояния между описываемыми ядрами значительно увеличиваются.
У аргасовых клещей дистальный и проксимальный отделы яйцеводов более резко обособлены друг от друга и отличаются гистологическим строением. Начинающиеся от рогов матки дистальные концы яйцеводов расширяются в ампуловидные образования. Клетки эпителиальных стенок дистальных отделов в зависимости от степени заполнения яйцеводов спер- миофорами столбчатые (высота 30—40 мк) или уплощенные (20—25 мк). Ядра сильно гипертрофированные, неправильно многоугольные или лопастевидные, диаметром около 20 мк. Апикальные концы клеток часто заполнены крупными вакуолями с секретом, которые напоминают таковые в матке, но значительно меньших размеров и никогда не заполняют всей клетки (рис. 422). Между эпителиальными клетками внедряются спермиофоры, причем часть из них несомненно подвергается фагоцитозу. Ампуловидные расширения у самок Ornithodoros papillipes могут достигать 400—600 мк в диаметре, тогда как суживающиеся к матке концы — 150-200 мк.
Проксимальные отделы телескопически вдвинуты в ампуловидные расширения, так что создаются клапанообразные структуры. Диаметр этих отделов обычно составляет 80—100 мк, клетки столбчатые высотой 30—40 мк. Признаков секреторной активности в них не наблюдается и ядра не превышают в поперечнике 9—10 мк (рис. 423).
У иксодовых клещей строение матки сходно с дистальными концами яйцеводов, от которых она отличается лишь большим диаметром и сильнее развитым мышечным слоем. Матка аргазид представляет исключительно сильно развитое железистое образование. Толщина ее стенок, построенных из одного слоя сильно вытянутых клеток, составляет от 100 до 200 мк. Секрет в виде крупных вакуолей (15—20x30—60 мк) занимает большую часть клеток. Ядра диаметром 6—8 мк лежат базально на границе с соединительнотканной оболочкой. Последняя образует многочисленные и глубокие внутренние впячивания, обеспечивающие возможность увеличения объема органа при нахождении в нем сперматодоз или яиц. Обширная полость матки после копуляции содержит крупные спермато- дозы и большое количество секрета. После выхода спермиофоров стенки матки слегка спадаются, а внутри она содержит аморфную массу из секрета и разрушающихся спермиофор (рис. 421).
Соединительная трубка Hyalomma asiaticum отличается очень глубокими складками кутикулярной выстилки, причем внутри складок находятся многочисленные ядра гиподермальных клеток (рис. 128). У голодных особей концы кутикулярных складок соприкасаются и просвет трубки напоминает сильно извитый лабиринт. Снаружи соединительная трубка окружена очень толстым слоем кольцевой мускулатуры, который во время кровососания значительно увеличивается. Просвет трубки в это время также расширяется благодаря расхождению складок. При яйцекладке стенки трубки сильно растягиваются проходящими яйцами и кутикулярные складки полностью расправляются. У Ixodes ricinus и Оті- thodoros papillipes соединительная трубка обладает сходным гистологическим строением и лишь несколько короче (рис. 414).
Полость вестибулярного отдела влагалища на поперечных срезах имеет вид щелевидного пространства, сплющенного в вертикальной части органа в передне-заднем и в горизонтальной — в дорсо-вентральном направлениях (рис. 117 и 418).
Кутикулярная выстилка очень сходна с таковой вентральной поверхности тела и, по существу, представляет ее внутреннее впячивание. Под ней лежит гиподермальный слой из столбчатых или кубических клеток высотой 8—10 мк с небольшими овальными ядрами в 5—6 мк. Снаружи гиподермальный слой окружает соединительнотканная оболочка, а ближе к цервикальной части кольцевая мускулатура (рис. 117).
С началом питания иксодовых клещей гиподерма вестибулярного отдела отделяется от кутикулы и, разрастаясь, образует многочисленные складки лопастевидной придаточной железы. Рост последней осуществляется за счет многочисленных митотических делений, обычно прекращающихся к моменту отпадения с хозяина. У напитавшегося клеща кутикулярная трубка вестибулярного отдела связана с гиподермой лишь на границе с половой щелью и цервикальным отделом, а в основной своей части окружена обширной полостью вновь образовавшейся железы (рис. 413).
Клетки этой железы достигают высоты 40—60 мк и в их апикальной части появляются многочисленные мелкие эозинофильные вакуоли с секретом. Последний выводится в полость железы и максимальное его количество скапливается к началу яйцекладки. По строению клеток, свойствам и времени накопления секрета лопастевидная железа напоминает железы органа Жене (рис. 122).
Цервикальный отдел влагалища Hyalomma asiaticum невелик и выстлан толстой кутикулой, образующей складки неправильных очертаний (рис. 118). Снаружи он окружен сильно развитой кольцевой мускулатурой. Кроме того, от его наружной поверхности отходят мышечные пучки, идущие в передневентральном направлении и прикрепляющиеся к стенке тела спереди от половой щели. Они сокращаются во время яйцекладки синхронно с дорво-вентральной мускулатурой и обеспечивают выпячивание наружу вестибулярного отдела. В цервикальном отделе АтЫуот- тіпае и Argasidae заметных изменений во время яйцекладки не обнаружено. У Ixodinae обширный цервикальный отдел по своему гистологическому строению и функциям идентичен семеприемнику АтЫуоттіпае (рис. 129).
Шейка и прилегающая к ней передняя стенка семеприемника Hyalomma asiaticum, как и цервикальный отдел, выстланы толстой кутикулой и кубической гиподермой, а снаружи окружены хорошо развитой кольцевой мускулатурой. Стенки большей части семеприемника собраны в глубокие складки. Кутикулярная выстилка тонкая (около 3 мк). Высота клеток составляет 5—7 мк и диаметр ядер — 3—4 мк (рис. 394). После оплодотворения стенки семеприемника растягиваются сперматодозами и кутику- лярные складки расправляются. Высота клеток увеличивается местами до 30—40 мк, а размеры ядер до 10—12 мк. Базальная часть цитоплазмы становится волокнистой, а в апикальной появляются мелкие вакуоли с эозинофильным секретом. После выхода спермиофор из сперматодоз кутикулярная выстилка во многих участках отслаивается от гиподермы (рис. 411). Полость семеприемника наряду с остатками сперматодоз содержит большое количество эозинофильного вещества.
Трубчатые придаточные железы у голодных самок Hyalomma asiaticum и Ixodes ricinus имеют диаметр в 30—40 мк. Высота клеток — 13—15 мк, а клеточные границы неразличимы. Мелкие сферические или овальные ядра в 5—6 мк очень тесно прилегают друг к другу и на срезах кажутся лежащими в несколько рядов. Секреторные гранулы в цитоплазме отсутствуют, а просвет железы очень мал (рис. 119).
За период кровососания диаметр желез увеличивается до 200 мк. Железистые клетки приобретают пирамидальную форму, достигают высоты 70—80 мк и их свободные концы вдаются в увеличивающийся в размерах просвет (рис. 120). Ядра подобных клеток достигают диаметра в 20—25 мк. Цитоплазма базальной зоны волокнистая, с высоким содержанием РНК. Между железистыми клетками ближе к просвету обнаруживаются одиночные мелкие ядра в 5—6 мк, сходные с таковыми голодных особей. В апикальных концах появляются многочисленные секреторные вакуоли белковой природы. К началу яйцекладки вакуоли заполняют всю цитоплазму и местами прикрывают ядра (рис. 416). Во время яйцекладки апикальные концы клеток разрушаются и секрет изливается в полость железы. Остающаяся цитоплазма спадается, а ядра подвергаются пикнотическим изменениям.
У Ornithodoros papillipes диаметр трубчатых придаточных желез в процессе секреции меняется меньше и обычно составляет 80—120 мк. У недавно перелинявших особей секреторные вакуоли в цитоплазме отсутствуют, а клеточные границы неразличимы. Ядра двух типов: мелкие — диаметром 5—7 мк и крупные — 10—14 мк. Перед яйцекладкой четко выявляются границы железистых клеток, а в их цитоплазме накапливаются мелкие и средние вакуоли с секретом. Содержимое вакуолей изливается в просвет железы, но разрушения клеток при этом не происходит. Описанные вакуоли сохраняются у голодных особей, а при повторных гонотрофических циклах число их нарастает и содержимое выводится в полость железы (рис. 420).
Процесс яйцекладки у иксодовых и аргасовых клещей протекает сходным образом. Он был прослежен нами на самках Hyalomma asiaticum. Перистальтическими сокращениями стенок яичника и яйцеводов яйца продвигаются в сторону влагалища. В то время как в яйцеводах одновременно находится множество яиц, придающих яйцеводам четковидную внешность, в вестибулярном отделе влагалища может находиться только одно яйцо. При сокращении дорсо-вентральной мускулатуры и пучков мышц, идущих от влагалища к вентральной стенке тела, вестибулярный отдел выпячивается вместе с яйцом через наружное половое отверстие. Одновременно орган Жене выворачивается над дорсальной поверхностью гнатосомы. Последняя подгибается на брюшную сторону, а пальпы при этом слегка раздвигаются. Рога органа Жене соприкасаются и охватывают яйцо, лежащее на конце вестибулярного отдела. Яйцо приклеивается к поверхности рогов и обволакивается восково-липоидным водонепроницаемым секретом. При изгибании гнатосомы дорсально благодаря выступающему краю спинного щитка яйцо переносится на спинную поверхность клеща. После этого весь цикл повторяется сначала, так что на дорсальной стороне скапливается комок яиц.
Еще по теме ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАЗМНОЖЕНИЯУ САМОК:
- Цитологические и гистологические особенности костного мозга и периферической крови у больных МДС
- ЧАСТЬ ДВАДЦАТАЯ РАЗЛИЧНЫЕ СОСТОЯНИЯ ОРГАНОВ РАЗМНОЖЕНИЯ У МУЖЧИ
- Диагностика и особенности цитологической, кольпоскопической картины папилломавирусной инфекции, гистологического метода исследования у беременных. Состояние влагалищного биотопа у женщин с папилломавирусной инфекцией.
- Органы размножения самок.
- ОПРЕДЕЛИТЕЛЬ САМОК МОСКИТОВ
- 3. Нейрогуморальна регуляція статевої функції самок і самців.
- Цитологическая диагностика.
- Цитологическое исследование.
- Аспирационная биопсия и цитологическое исследование
- 3.2. Методики цитологического исследования
- Цитологическое отделение
- Цитологические и кольпоскопические картины у обследованных беременных.
- 3.1. Морфология половых органов самок свиней, содержавшихся в различных условиях двигательной активности
- 3.3. Вены половых органов самок свиней, содержавшихся в различных условиях двигательной активности
- Цитологическое исследование мокpоты
- Цитологическое исследование отделяемого из соска
- 1.2 Этапы развития цитологического метода в диагностике заболеваний желудка
- Цитологическое исследование мокроты
- Цитологическое исследование.