ОРГАННЫЕ И СИСТЕМНЫЕ АДАПТАЦИИ У МЛЕКОПИТАЮЩИХ И ПТИЦ
Приспособления органов и систем к разным условиям существования млекопитающих и птиц часто хорошо выражены морфологически. Густота и теплопроводимость шерстного и перьевого покровов, распределение жира, длина ушей, хвоста, конечностей, вес сердца, лег-
KIIX, почек п т.
д. могут служить наиболее яркими примерами. Морфологические и физиологические адаптации отдельных органов оказываются настолько мощными, что другие системы организма могут иногда совершенно не изменяться в связи с приспособлением к крайним условиям среды. Таковы, например, изменения структуры почек у пустынных видов млекопитающих. Приспособительные реакции органов могут охватывать все системы, включая систему крови.Морфологические адаптации органов пищеварения хорошо выражены, проявляются в особом строении ротовой полости, желудка, длине кишечника. В качестве физиологических адаптаций можно привести многочисленные примеры изменений активности ферментов в связи с характером питания (Уголев, 1958) и двигательной функции кишечника у близких видов грызунов, особенностей водного обмена в кишечнике у верблюда в связи с существованием в аридной зоне (жаркие пустыни). Эти системные физиологические адаптации обеспечивают существование организма в естественных для вида условиях питания, снабжения водой и т. д.
Система крови также находится в тесной связи с экологической специализацией животного. Это касается прежде всего ее дыхательной функции (содержания эритроцитов, гемоглобина, кривой диссоциации оксигемоглобина, активности угольной ангидразы). Увеличение способности крови связывать кислород и переносить его тканям наблюдается главным образом в процессе приспособления к гипоксии (к условиям пониженного барометрического давления), интенсивной мышечной работе, нырянию (Барбашова, 1941, 1960; Крепе, 1936). При частичном понижении температуры тканей (например, при очень низкой температуре конечностей) перенос кислорода кровью обеспечивается соответствующим изменением содержания гемоглобина в крови.
У северного оленя в зимний период наблюдается повышенное (с 77 до 90% по Сали) содержание гемоглобина в крови. Одновременно температура тела конечностей оленя падает до 4-4— 10°С. Очевидно, высокая концентрация гемоглобина в крови обеспечивает снабжение кислородом тканей при таких исключительно низких температурах среды.Органы выделения изменяются у млекопитающих различной экологической специализации главным образом в связи с характером обмена веществ и особенностями водного обмена в частности. В этом отношении большое значение имеют морфологические особенности — соотношения между толщиной коркового и мозгового слоев почки и соответственно длиной канальцевого аппарата почки. У пустынных форм млекопитающих (грызунов) обнаруживается крайне увеличенный корковой слой с длинными канальцами, покрытыми утолщенным цилиндрическим эпителием. Этим морфологическим отличиям соответствует и большая способность к концентрации мочи, достигающая соотношений первичной мочи ко вторичной, как 1000/1 вместо 100/1 для животных — обитателей орошаемых районов (Schmidt-Nielsen а. Schmidt-Nielsen, 1952). У некоторых хищных (например, львов) наблюдаются аналогичные отношения, хотя по характеру водного обмена они не отличаются от непустынных млекопитающих (Sperber, 1944). К сожалению, с этой стороны изучены лишь немногие важные в эколого-физиологическом отношении виды млекопитающих.
У птиц органы выделения в значительной мере адаптированы к выведению повышенной концентрации солей. Особенно ярко выражены эти явления у птиц — обитателей пустынной зоны (Dawson a. Schmidt-Nielsen, 1964).
Очень большие морфологические изменения в связи с условиями обитания претерпевают у млекопитающих кожа и ее придатки. Следует отметить общую закономерность, проявляющуюся в величине шерстного покрова и его теплоизоляционных свойствах в связи с сезонами года, а главное в связи с ареалом данного вида (Калабухов, 1950; Scholander, Hock, Walters a. Irving, 1950) (рис. 5). Увеличенные теплоизолирующие свойства шерстного покрова характерны для полярных млекопитающих, а также для зимнего периода.
Сезонная смена мехового покрова происходит под непосредственным влиянием режима освещения и, в частности, ритмической смены его на протяжении суток. Поэтому раннее наступление зимы или перевод (в целях акклиматизации) животных из северных районов в более южные может вызывать нарушения терморегуляции. Относительно длинный день в южных условиях не вызывает соответствующей температуре среды смены мехового покрова, її животные оказываются таким образом «неподготовленными» к зиме.Кожа, являющаяся органом испарения, очень сильно отличается у разных видов и морфологически (по строению и функциональной активности потовых желез), и по способности к отдаче (перспирации) воды из тканей.
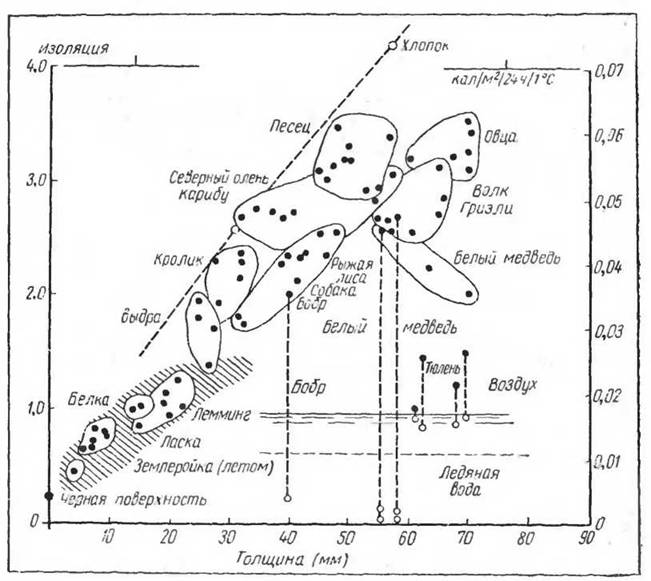
Рис. 5. Теплоизоляция в зависимости от толщины зимнего шерстного покрова у ряда арктических млекопитающих (точки, не окруженные штриховкой) и тропических (окруженные штриховкой) (по Scholander и сотр., 1950). Теплая сторона кожи 37°С во всех случаях; вода и воздух 0°С. Видно, что лемминг не отличается от тропических видов
Ограниченная способность к кожной перспирации наблюдается у пустынных грызунов (Кашкаров и Коровин, 1936). Эта особенность проявляется и в морфологических изменениях кожи. У крупного рогатого скота.
акклиматизируемого в условиях жаркого сухого климата, обнаружено возрастание числа потовых желез и увеличение их функциональной активности (Раушенбах, 1952, 1958). Последняя после акклиматизации приближалась к активности, свойственной аборигенному крупному рогатому скоту Средней Азии.
Кожа и ее сосудистый аппарат как орган терморегуляции имеет у различных видов крайне специализированный характер. Так, например, поверхность кожи уха кролика и зайца чутко реагирует на изменение температуры (а следовательно, и интенсивность теплоотдачи) на мышечную работу, производимую животным. Нарушение симпатической иннервации уха не только устраняет эту тонкую регуляцию при мышечной работе, но и ограничивает продолжительность работы вследствие наступления перегревания.
Если, например, интактный кролик способен пробежать в третбапе около трех часов, то после односторонней перерезки симпатического ствола в шейной области кролик прекращает работу при той же температуре тела (около +42°С) после 1,5-часового бега той же интенсивности (Склярчик, 1963). У крыс таким органом является хвост. К сожалению, дикие виды млекопитающих с этой стороны совершенно не изучены.У птиц перьевой покров и его теплоизоляционные свойства также сильно варьируют в зависимости от условий обитания и сезонов года (King a. Farner, 1961; Шилов, 1968).
Разнообразие в характере и интенсивности мышечной деятельности обеспечивается столь же разнообразными изменениями мышечной системы, носящими адаптивный характер. Они ярко выражены в виде большей или меньшей степени развития соответствующей мускулатуры у роющих, бегающих, прыгающих форм (Огнев, 1951; Гамбарян, 1953). Однако физиологические особенности отдельных высокоспециализированных мышечных групп почти совершенно не изучены.
Поскольку в мышечной ткани осуществляются главным образом окислительные процессы, между развитием скелетной мускулатуры, интенсивностью потребления кислорода и интенсивностью мышечной деятельности имеют место самые тесные зависимости. Особенно ярко они могут быть прослежены при рассмотрении строения и 57
функции сердечно-сосудистой системы и главным образом сердца.
Лучше других отделов кровеносной системы изучено сердце, особенно его морфология и отношение веса к общему весу тела, обычно выражаемое в °/оо- Строение и размеры сердца отражают не только видовые особенности мышечной деятельности, но и общий уровень обмена веществ, связанный с химической терморегуляцией (Шв арц, 1959), а также увеличенный кровоток в связи с недостатком кислорода в горах (Аіашковцев, 1935). Значение видовых особенностей строения сердца для характеристики различных видов млекопитающих в связи с их мышечной деятельностью установлено многими исследователями (Strohl, 1910а, 19106; Hesse, 1921; Машковцев, 1935; Шварц, 1959 и мн.
др.).Как правило, относительные размеры и вес сердца уменьшаются с увеличением размеров тела (внутри одного вида). Многочисленные примеры этого явления приведены в цитированной выше книге С. С. Шварца. Что касается изменений веса сердца в связи с природными климатическими условиями, то на основании имеющегося сейчас уже большого материала, приводимого в книге Шварца, и сопоставления его с данными по химической терморегуляции у палеарктических, субарктических и арктических видов (Слоним, 1952; Быков и Слоним, 1960) имеется полная зависимость между интенсивностью химической терморегуляции и относительным весом сердца. У палеарктических и субарктических видов лучше всего развита химическая терморегуляция, а размеры сердца увеличены. У арктических видов размеры сердца могут не отличаться от южных форм.
Легочное дыхание, как и кровообращение, находится в тесной зависимости от интенсивности окислительных процессов, от особенностей содержания кислорода в окружающей среде. Уже давно установлено увеличение размеров грудной клетки у человека в горах (Forbes, 1870; Barcroft, 1925). Увеличенные размеры легких, больший диаметр бронхов и сильно развитая бронхиальная мускулатура характеризуют породы овец и коз, разводимых в условиях гор Заилийского Алатау (Лебедева, 1954). У земноводных, обитающих в горах, наблюдается большее развитие легочного дыхания, нежели у видов, обитающих на равнине (Шварц, 1959).
Еще по теме ОРГАННЫЕ И СИСТЕМНЫЕ АДАПТАЦИИ У МЛЕКОПИТАЮЩИХ И ПТИЦ:
- Паразиты птиц и млекопитающих.
- ОРГАННЫЕ И СИСТЕМНЫЕ АДАПТАЦИИ У ПОЙКИЛОТЕРМНЫХ ОРГАНИЗМОВ
- Системная методология. Сущность и практическое назначение системного подхода
- Моделирование у млекопитающих
- Особенности пищеварения и водного обмена пустынных млекопитающих
- 2.4 Имитационное моделирование вычисления показателей синхронности системных ритмов для двухканального источника системных ритмов
- Миграции птиц.
- Вирус гриппа птиц
- Морфофункциональная характеристика клеточных элементов пульпы развивающихся и сформированных зубов млекопитающих
- АДАПТАЦИИ ПТИЦ К ПУСТЫНЕ
- МЫШЕЧНАЯ ДЕЯТЕЛЬНОСТЬ У ПТИЦ
- ХОЛОДОВОЕ ОЦЕПЕНЕНИЕ У ПТИЦ (ТОРПИДНОЕ СОСТОЯНИЕ)