Трансмембранные а-спирали.
У водорастворимых белков обнаружены сотни определённых белковых мотивов (или повторов). В отличие от них количество "стандартных" мотивов в интегральных мембранных белках достаточно невелико.
Чаще других встречается трансмембранная а-спираль.Интегральные белки, содержащие трансмембранные а-спиральные домены, удерживаются в мембране благодаря гидрофобному взаимодействию со специфическими липидами, а также, возможно, ионному взаимодействию с полярными головками фосфолипидов.
Наглядным примером такого рода белков является гликопротеин А, основной белок плазматической мембраны эритроцитов. Его трансмембранным доменом является одна а-спираль (рисунок 36).
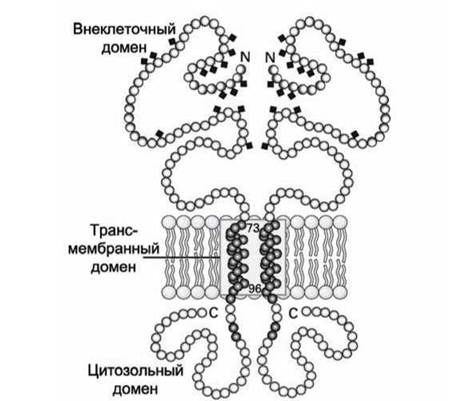
Рисунок 36 - Схема димера гликопротеин А
Обычно трансмембранная а-спираль состоит из 20-25 гидрофобных аминокислот. У гликопротеина А трансмембранная а-спираль состоит из 23 аминокислот. Длина такой спирали составляет порядка 3,75 нм, что хорошо согласуется с данными о толщине липидного бислоя.
В а-спирали гидрофобные аминокислотные радикалы ориентированы иаруж-у и взаимодействуют с гидрофобным липидным окружением, а карбонильные (С=О) и имино- (NH) группы, которые формируют внутри- спиральные водородные связи, расположены внутри а-спирали и экранированы от липидной среды аминокислотными остатками. Энергетически выгодно двум гликопротеинам А образовать димер за счёт формирования мотива "скрученная спираль" (coiled coifj трансмембранными спиралями.
Такая димеризация белков за счёт формирования скрученных спиралей является распространённым явлением. Например, многие мембранные рецепторы активируются только после димеризации.
Существует обширное "семейство" (более 150) интегральных мембранных белков, характеризующихся наличием семи трансмембранных а-спиралей.
К таким "семиспиральным" многопроходным белкам относятся G-протеин-связанные рецепторы, играющие ключевую роль в клеточной сигнализации.Общие принципы построения белков этого семейства иллюстрирует структура бактериородопсииа (рисунок 37), который выполняет функцию фоторецептора в определённых бактериях.
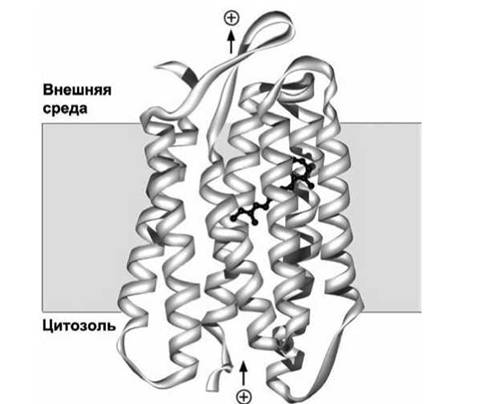
Рисунок 37 - Схема бактериородопсииа. Молекула ретиналя изображена чёрным цветом
К одной из трансмембранных а-спиралей ковалентно присоединена молекула ретиналя. В результате поглощения кванта света молекула ретиналя изменяет конформацию (рисунок 38), что стимулирует информационный переход в белковой молекуле, в результате чего происходит
транслокация одного протона из цитозоля через мембрану во внеклеточное пространство.
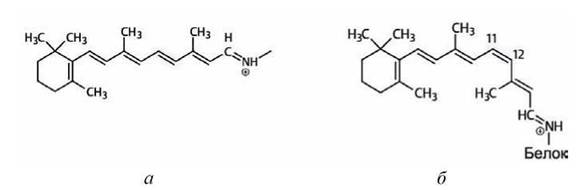
Рисунок 38 - Конформации молекулы ретиналя: а - полностыо-транс реї шіа.іь, 5 - 11-цис pc i нііалі,
Бактериородопсин работает как протонный насос, создавая за счёт энергии света градиент концентрации протонов на мембране. Этот протонный градиент используется затем для синтеза АТФ мембранными FoFi-комплексами (АТФ-синтазами). Экспериментальное определение атомной структуры трансмембранного домена показало, что, действительно, практически все аминокислоты на периферии трансмембранного домена бактериородопсина являются гидрофобными и взаимодействуют с углеводородной внутренней частью липидного бислоя.
Другим важным классом интегральных мембранных белков являются ионные каналы, структура и функции которых будут подробно рассмотрены в гл. 13.
Ионные каналы являются преимущественно тетрамерными белками. Каждая из четырёх субъединиц имеет как минимум две трансмембранные а-спирали, а все вместе а-спирали всех субъединиц формируют селективный водный канал, который пропускает только ионы данного типа.
Аминокислоты, которые "выстилают" этот канал, являются полярными и гидрофильными, а периферические аминокислоты трансмембранного домена ионного канала, так же как и в случае бактериородопсина, являются гидрофобными.
Во многих ионных каналах воздействие внешних факторов, таких как связывание с лигандом, изменение мембранного потенциала, механическое воздействие, приводит к изменению взаимного расположения а-спиралей, что используется для регулировки потока ионов через мембрану.
Еще по теме Трансмембранные а-спирали.:
- СЛОВАРЬ ТЕРМИНОВ
- Изучение ориентации белков в мембранах
- Трансмембранные а-спирали.
- Трансмембранные P-структуры.
- Трансмембранный транспорт глюкозы.
- Механизмы белкового транспорта
- Фильтры селективности ионных каналов.
- 11.1. МА+-СИМПОРТЁРЫ
- 11.5. Аквапорины
- Строение потенциалочувствительных ионных каналов
- Ацетилхолиновый рецептор