Механизмы белкового транспорта
Мембранные структуры клетки активно участвуют во внутриклеточном транспорте белков. После синтеза белков на рибосомах в цитозоле эукариотических клеток эти белки должны быть доставлены для их работы в необходимые органеллы (или за пределы клетки).
Существует три основных механизма, с помощью которых клетка решает эту задачу (рисунок 76).
1. Белок после синтеза и фолдинга в неизменном виде доставляется в нужную органеллу через специализированные поры в мембранах. Такой вид доставки называется управляемый транспорт (gated transport).
2. Трансмембранная транслокация (transmembrane translocation} белков, в ходе которой полипептид сначала денатурируется, затем полипептидная цепочка протягивается через одну или несколько мембран, а затем снова происходит фолдинг функционального белка.
3. Везикулярное движение (vesicular trafficking белков, в ходе которого от мембраны отпочковывается везикула, в состав которой входят транспортируемые белки.
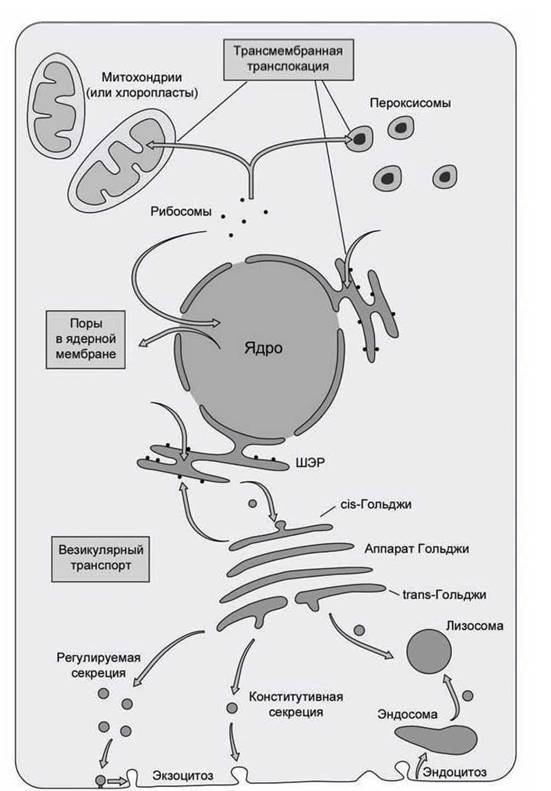
Рисунок 76- Механизмы внутриклеточного транспорта белков
С помощью управляемого транспорта, например, в ядро клетки через ядерные поры доставляются все белки. Трансмембранная транслокация обеспечивает доставку синтезированных в цитозоле белков в пероксисомы, митохондрии и хлоропласты. Везикулярное движение обеспечивает доставку белков в лизосомы и секрецию белков из клетки.
Адресация белков обеспечивается наличием в их структуре специальных сортировочных сигналов (sortingsignal^.
Сразу после окончания синтеза белка рибосомой в цитозоле, когда он является просто полипептидом, сортировочными сигналами являются последовательности аминокислот на концах белковой цепи, которые называют нацеливающими последовательностями (targeting sequences или localization sequence^.
Для белков, которые синтезируются на мембранах шероховатого эндоплазматического ретикулума, дополнительные сортировочные сигналы (sorting signal^ (такие, как сахара или фосфатные группы) могут быть добавлены с помощью специализированных ферментов в цистернах аппарата Гольджи в ходе посттрансляционной модификации белков.
Такие сигналы обычно представляют собой специфические лиганды, которые распознаются рецепторными белками, а эти рецепторные белки с присоединёнными к ним транспортируемыми белками, в свою очередь, присоединяются к мембранным транслокационным комплексам соответствующего компартмента.
Нацеливающие последовательности белка, которые представляют собой цепочку из 3-80 аминокислот, тоже распознаются специализированными рецепторами, которые доставляют данный белок к соответствующим транслокационным комплексам. После доставки в нужный компартмент нацеливающие последовательности обычно отрезаются от белковой цепи специализированными ферментами.
Одними из наиболее изученных нацеливающих последовательностей являются сигнальные пептиды (или сигнальные последовательности, signal sequence^ - цепочки из 5-15 преимущественно гидрофобных аминокислот. Наличие такой сигнальной последовательности в синтезируемом белке вынуждает рибосому присоединиться к эндоплазматическому ретикулуму, и синтезируемая белковая цепь направляется не в цитозоль, а в люмен ретикулума.
Другой пример нацеливающей последовательности - сигнал импортирования {import signal} для белков, которые должны быть перемещены из цитозоля в матрикс митохондрии. Этот сигнал представляет собой цепочку из 20-80 аминокислот, формирующих амфипатическую полярную а-спираль, у которой положительно заряженные аминокислоты выстроены с одной стороны спирали, а гидрофобные аминокислоты - с другой.
Для нацеливания белков в ядро клетки определена последовательность из пяти положительно заряженных аминокислот.
Для переноса белков в пероксисому служит пероксимальная нацеливающая последовательность Ser-Lys-Lys-COOH - С-конечный трипептид.
Существуют также сортировочные сигналы, которые не способствуют перемещению белка, а наоборот, служат сигналом о том, что белок уже доставлен к месту назначения и никуда далее его не следует перемещать. Например, белки с так называемой KDEL-последова- тельностью Lys-Asp-Glu-Leu-COOH на С-конце остаются в эндоплазматическом ретикулуме и не должны удаляться из него везикулярным транспортом.
Иллюстрацией вышесказанного может быть кальций-связывающий белок гладкого эндоплазматического ретикулума калретикулин {calciumbinding protein of the endoplasmatic reticulum - calreticuliii), чья первичная структура изображена на рисунке 77.
17 аминокислот
(NH2 jMLLSVPLLLGLLGLAVAFPAVYFKEQFLDGDGWTSRWIESKHKSDFGKFVLSSGKF
YGDEEKDKGLQTSQDARFYALSASFEPFSNKGQTLVVQFTVKHEQNIDCGGGYVKLFP
NSLDQTDMHGDSEYNIMFGPDICGPGTKKVHVIFNYKGKNVLINKDIRCKDDEFTHLYT
LIVRPDNTYEVKIDNSQVESGSLEDDWDFLPPKKIKDPDASKPEDWDERAKIDDPTDSKP
EDWDKPEHIPDPDAKKPEDWDEEMDGEWEPPVIQNPEYKGEWKPRQ1DNPDYKGTWIH
PEIDNPEYSPDPSIYAYDNFGVLGLDLWQVKSGTTFDNFUTNDEAYAEEFGNETWGVTK
AAEKQMKDKQDEEQRLKEEEEDKKRKEEEEAEDKEDDEDKDEDEEDEEDKEEDEEED
VPGQA.KDEL(COOH)
4 аминокислоты
Рисунок 77- Аминокислотная последовательность калретикулина
Первые семнадцать аминокислот на N-конце калретикулина являются сигнальной последовательностью, которая инициирует транслокацию белка в люмен эндоплазматического ретикулума, а последние четыре аминокислоты - последовательность KDEL - не позволяет белку уйти из ретикулума. Между этими сортировочными сигналами располагается первичная структура функционального белка.
8.2.
Еще по теме Механизмы белкового транспорта:
- Транспорт белков в митохондрию.
- Глава 8 Внутриклеточный транспорт белков
- Транспорт веществ через мембрану с помощью БЕЛКОВ-ТРАНСПОРТЁРОВ
- Свойства селена, механизм транспорта и биохимических превращений в клетке
- Транспорт в пероксисомы.
- Глава 7 Активный транспорт
- 2.5. Исследование времени мукоцилиарного транспорта.
- Недостаточность транспорта лимфы
- Транспорт железа в организме.
- ТРАНСПОРТ ДВУОКИСИ УГЛЕРОДА
- Везикулярный транспорт
- Дезинфекция вооружения, боевой техники, транспорта
- Глава 11 Вторичный транспорт котранспортёрами
- Медико-тактическая характеристика чрезвычайных ситуаций на водном и трубопроводном транспорте.
- Трансэпителиальный транспорт
- 6. Катастрофы на речном и морском транспорте