Штаммы-продуценты аргинина и методы их получения
Представления о механизмах, контролирующих биосинтез данной аминокислоты, о взаимосвязи путей ее синтеза с синтезом других метаболитов составляют ту теоретическую основу, на которой обычно строится схема селекционной работы, предусматривающая получение определенных типов мутантов, свойства которых должны отвечать ряду поставленных требований.
Сверхсинтез аминокислот обычно не наблюдается у диких штаммов микроорганизмов. Исключение составляют продуценты глутаминовой кислоты, аланина и валина. Целью селекционной работы является преобразование метаболизма таким образом, чтобы вызвать сверхпродукцию желаемой аминокислоты. Это достигается методами селекции, направленными на устранение негативного контроля биосинтеза целевого продукта (т.е., устранение факторов, определяющих ингибирование и репрессию его синтеза).
Накопление интермедиатов может осуществляться в результате генетического блокирования метаболического пути и лимитации необходимого для роста продуцента конечного продукта. Т.е., в клетке блокируется биосинтез и, как следствие, накопление веществ, подавляющих образование целевого продукта или вызывающих его отток (либо его предшественников) на свой биосинтез. Блокированное вещество вносится затем искусственно в питательную среду в минимальных концентрациях, необходимых для развития культуры. При правильном выборе вещества, по которому создается ауксотрофность, реализация этого метода позволяет резко увеличить биосинтетическую способность штамма. В этом состоит принцип использования ауксотрофных мутантов в производстве аминокислот. Часто целесообразно создание дефекта по гену первого фермента боковой ветви пути биосинтеза. Если целевой продукт не конечный в данном метаболическом пути, то инактивация фермента дальнейшего превращения этого вещества также может оказаться необходимым. У полученных ауксотрофных мутантов сверхсинтез целевого продукта может быть обусловлен тем, что ингибирование или репрессия первого фермента общей части пути носит поливалентный характер, так что избыток одного конечного продукта при дефиците второго не приводит к подавлению его активности.
Аминокислоту, биосинтез которой блокирован, следует ограничить в среде роста, создавая дефицит ее в клетке.Другим методом селекции является получение мутантов с нарушенной регуляцией биосинтеза целевого продукта, что чаще всего достигается отбором мутантов, устойчивых к структурному ана-
логу целевого продукта. Аналог, добавленный к минимальной среде, на которую высеяно газоном большое количество клеток исходного штамма, воздействует на регуляторную систему клеток, имитируя избыток природной аминокислоты, антагонистом которой он является. Синтез аминокислоты прекращается, клетки испытывают голодание по этой аминокислоте. Расти и образовывать колонии могут только мутанты, у которых нарушена регуляция биосинтеза целевого продукта (регуляторные мутанты). Такие мутанты называются аналогорезистентными.
Механизм аналогорезистентности определяется типом регуляции: если контроль осуществляется на уровне транскрипции (т.е. по механизму репрессии), то отбираются частично или полностью дерепрессированные мутанты; если же биосинтез продукта контролируется по типу ретроингибирования, то отбираются клоны с ферментом, нечувствительным к ретроингибированию, т.е. десенсибилизированным. Так как фенотип резистентности к аналогам аминокислот может быть следствием мутаций как регуляторных, так и различных структурных генов, то использование этого метода при всей его эффективности часто дает неожиданные, в том числе негативные результаты из-за недостатка сведений о причинах резистентности.
Для повышения продуктивности, как правило, применяют комбинацию ауксотрофности и аналогорезистентности, т.е. в результате селекции получают штамм, ауксотрофный по одному (редко двум) ключевым веществам (в случае продуцентов аргинина это преимущественно аминокислоты и пиримидины, образование которых связано с его биосинтезом) и устойчивый к различным аналогам целевого продукта (т.е. веществам, которые ингибируют биосинтез целевого продукта, не выполняя при этом в клетке его функций).
Использование ауксотрофности и аналогорезистентности одновременно в одном штамме позволяет минимизировать контроль образования целевой аминокислоты как конечным, так и побочными продуктами биосинтеза. Эти два способа отбора мутантов широко используются в селекции продуцентов аминокислот.Продуценты, пригодные для промышленного применения, обычно проходят многоэтапную ступенчатую селекцию на устойчивость к целому ряду аналогов целевой аминокислоты и несут несколько мутаций, повреждающих те или иные механизмы регуляции как самой аминокислоты, так и ее предшественников. Большое количество мутаций обеспечивает высокий уровень накопления продукта и конверсии источников углерода в целевую аминокислоту, но может сильно ослабить жизнеспособность штамма. Основной причиной этого является накопление в ходе последовательных циклов мутагенеза мутаций, которые инактивируют биосинтетические функции штамма (например, могут быть затронуты гены репликации, транскрипции и др.).
Если для данного вида микроорганизмов выяснены закономерности и разработаны методы генетического обмена, то отпадает необходимость в длительной ступенчатой селекции. Мутации, полученные в разных линиях селекции, можно объединить в одном геноме. В последнее время в селекции продуцентов аминокислот активно используются методы генной инженерии, позволяющие повышать дозу генов биосинтеза аминокислот путем их клонирования на плазмидах.
В настоящее время известен ряд продуцентов аргинина, включающий аналогорезистентные мутанты Corynebacterium glutamucum, Bacillus subtilis и Serratia marcescens, а также рекомбинантные штаммы Escherichia coli. Аэробные, спорулирующие, грамположительные микроорганизмы, относимые к роду Bacillus, — эффективные продуценты аминокислот. В. subtilis является классическим генетическим объектом, для анализа которого применяются генетическая трансформация и трансдук- ция. Е. coli и S. marcescens относятся к энтеробактериям — многочисленной группе грамотрицательных нефотосинтезирующих бактерий.
Е. coli является детально изученным объектом, и последнее время находит широкое применение в создании сверхпродуцентов генно-инженерными методами.Сверхпродукция аргинина у коринебактерий коррелирует с десенсибилизацией АГК и увеличением удельной активности АГК, КФС и некоторых других ферментов. Широко используются штаммы, резистентные по 6-азаурацилу, гидроксамату аргинина, D-аргинину, канаванину, 2-тиазолала- нину, аргинингидроксамату, 8-азагуанидину, сульфагуанидину, D-серину, цистеину и его аналогам. Некоторые ауксотрофные мутации, в частности, чувствительность к гистидину, треонину, триптофану, лизину, гуанину, также повышают уровень продукции аргинина у его продуцентов.
Штаммы-продуценты аргинина, используемые при разработке данного процесса биосинтеза аргинина, были получены селекционным путем в ФГУПГосНИИгенетике в 1995-1997 гг. Ю. А. Рыбаковым и Т.В. Леоновой. В качестве объекта исследования был выбран дикий штамм Brevibacterium flavum АТСС 14067, относящийся к группе глутаматсинтезирующих бактерий. В качестве селективных факторов при получении продуцентов аргинина использовали его аналоги — канаванин (Сап), гидроксамат аргинина (ArgHx), а также аналог урацила 6-азаурацил (AU). Кроме того, был использован цистеин (Cys), поскольку среди устойчивых к нему штаммов С. glutamicum высокой частотой отбираются варианты с повышенным уровнем биосинтеза аригинина.
Для каждого соединения определяли угнетающие рост концентрации и показывали, что добавление аргинина в среду устраняет ингибирующее рост действие аналога.
При проверке более 600 вариантов, устойчивых к различным аналогам, в группе мутантов, растущих в присутствии 1,0 мг/мл AU, был выбран клон А1 (рис. 7), продуцирующий 0,3 г/л аргинина. Путем двухступенчатой селекции на устойчивость этого клона к AU (до 2,5 мг/мл) был выделен вариант НЮ, синтезирующий до 1 г/л аргинина, а среди его цистеинрезистентных производных — штамм М12 с уровнем накопления аминокислоты 2,5 г/л.
В дальнейшем этот штамм был использован в качестве исходного для двух независимых линий отбора.
В первом случае, для селекции использовали смесь двух аналогов, AU (2 мг/мл) в сочетании либо с Сап, либо с ArgHx (по 3 мг/мл). Скриннинг мутантов, устойчивых к сочетанию (AU+Can), привел к выделению варианта VB4, от которого по устойчивости к смеси (AU+ArgHx), было получено несколько штаммов с продуктивностью до 5 г/л аргинина.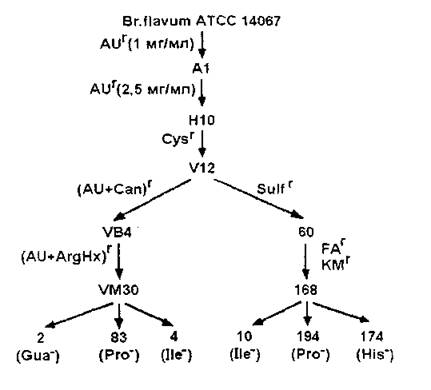
Рис. 7. Схема селекции продуцентов аргинина AUr, ArgHx-r, Cys-r, Can-r, Sulf-r, Fa-r, KM-r — устойчивость к 6-азаурацилу, гидроксамату аргинина, цистеину, канаванину, сульфагуанидину, монофторацетату и кетомалонату, соответственно
Второй подход включал получение мутантов с изменениями в энергетическом метаболизме, активизация которого могла бы благоприятно сказаться на продуктивности. На начальном этапе отбор проводили по устойчивости клеток к сульфагуанидину (1,5 мг/мл). Поскольку в культуральной жидкости мутантов присутствовали наряду с аргинином значительные количества глутаминовой кислоты (7-8 г/л), а также примеси аланина (3-4 г/л) и валина (1-2 г/л), в обеих линиях среди продуктивных по аргинину клонов отбирали варианты с минимальным содержанием глутамата.
Результатом селекции в указанных направлениях стало получение двух клонов, VM30 и 60, продуцирующих 4 г/л аргинина и глутамата не более 2 г/л. Содержание остальных побочных кислот не изменилось. Линия штамма 60 была продолжена получением мутантов, резистентных к смеси аналогов интермедиатов ЦТК монофторацетату (FA, 0,5 мг/мл) и кетомалонату (КМ, 1,5 мг/мл), устойчивость к которым также сопровождается изменениями энергетического обмена клетки и может проявляться в уменьшении оттока пирувата на образование аланина и валина. У одного из отобранных мутантов, 168, уровень накопления аргинина достиг 5 г/л.
На следующем этапе селекции методом обогащения были получены производные штаммов VM30 и 168, ауксотрофные по ряду аминокислот и оснований, прямо или косвенно связанных с путем биосинтеза аргинина.
Критериями выбора типа ауксотрофности служили описанные в литературе мутации недостаточности по гуанину, изолейцину и гистидину у штаммов-продуцентов Br. flavum и С. glutamicum, коррелирующие со сверхсинтезом аргинина, хотя регуляторные механизмы остаются неясными, а также возможность увеличения накопления аргинина у ауксотрофов по урацилу, потенциальному ингибитору КФС, и пролину, имеющему с аргинином обший предшественник.Среди более чем 60 ауксотрофов по пролину, изолейцину, гистидину, урацилу и гуанину были выбраны штаммы, превышающие родительские по уровню продукции на 20-40 % .
При ферментации на исходной питательной среде штаммы накапливали: Br. flavum 60 — 4 г л аргинина, VM30 — 5, 168 — 5, 174(His~) — 7, 2(Gua ) — 5,5, 4(І1е“) — 5, 194(Pro ) — 8, 83(Pro ) — 8 г/л. Штаммы накапливали в культуральной жидкости большое количество побочных аминокислот (аланина, валина, лейцина, гистидина), а также аминокислоты, чей биосинтез связан с биосинтезом аргинина — глутаминовую кислоту, глутамин, цитруллин, орнитин.
Сравнение полученных штаммов-продуцентов при оптимальных для них концентрациях ауксо- трофного фактора показало, что наибольшим уровнем накопления аргинина характеризуются штаммы-продуценты, ауксотрофные по пролину — 83(Рго ) и 194(Рго ), что обусловлено прекращением оттока общего предшественника — глутаминовой кислоты на образование пролина (табл. 1). Дальнейшая работа по созданию технологии биосинтеза L-аргинина было построена на использовании в качестве продуцентов пролинзависимых штаммов.
Табл. 1. Сравнение биосинтетических возможноетей продуцентов аргинина
| Штамм | Аминокислота, мкг/мл | on | Аргинин, г/л | Аланин, г/л |
| 60 | — | 72,5 | 4,2 | 1,2 |
| VM30 | — | 57,2 | 4,5 | 1,5 |
| 168 | — | 60,5 | 5,0 | 2,2 |
| 100 | 32,0 | 3,2 | — | |
| 200 | 38,2 | 6,0 | 0,6 | |
| 300 | 43,3 | 7,0 | 0,9 | |
| 174(His ) | 400 | 50,2 | 5,8 | 1,2 |
| 600 | 50,4 | 5,2 | 1,8 | |
| 800 | 50,5 | 4,8 | 1,5 | |
| 50 | 24,3 | 2,4 | 0,7 | |
| 100 | 40,1 | 5,0 | 1,2 | |
| 150 | 48,1 | 6,0 | 2,0 | |
| 2(Gua ) | 200 | 50,2 | 5,5 | 1,7 |
| 250 | 51,7 | 4,8 | 0,9 | |
| 300 | 46,8 | 4,3 | 0,4 | |
| 100 | 38,2 | 3,1 | 1,0 | |
| 4(І1е-) | 300 | 43,0 | 5,7 | 1,8 |
| 500 | 45,3 | 6,2 | 2,0 | |
| 700 | 47,7 | 4,3 | 1,2 | |
| 100 | 29,2 | 5,2 | 1,1 | |
| 200 | 32,7 | 7,5 | 1,8 | |
| 300 | 36,9 | 8,2 | 2,3 | |
| 83(Pro ) | 400 | 48,2 | 6,8 | 1,9 |
| 600 | 53,4 | 6,2 | 1,3 | |
| 800 | 53,2 | 5,0 | 1,0 | |
| 100 | 32,0 | 6,2 | 0,7 | |
| 200 | 43,0 | 8,3 | 1,7 | |
| 300 | 49,5 | 7,2 | 2,5 | |
| 400 | 48,3 | 6,0 | 1,7 | |
| 600 | 44,2 | 5,2 | 1,5 | |
| 800 | 40,0 | 4,3 | 1,2 |
3.
Еще по теме Штаммы-продуценты аргинина и методы их получения:
- Биосинтез аргинина мутантными штаммами-продуцентами Brevibacterium flavum
- Хранение продуцентов тилозина и проведение поддерживающей селекции штаммов
- Хранение исходных штаммов продуцентов
- Влияние условий культивирования продуцентов на биосинтез аргинина
- Основные физиолого-биохимические свойства штаммов-продуцентов рибофлавина
- Подбор бактерий-продуцентов ЦГТазы для процессов получения циклодекстринов
- Выбор продуктивного штамма для процессов получения ЦГТазы
- Процесс микробиологического получения рибофлавина с использованием генно-инженерного штамма Bac.subtilis
- 3.3 Сравнениерезультатов видовой идентификации штаммов Candida spp. методами MALDI-TOF масс-спектрометрии и тест-системой
- Методы получения фтора-18
- Методы статистической обработки полученных данных
- 2.1.2. ТЕМА: Методы получения эхографического изображения.
- 2.7 Методы статистического анализа полученных данных
- Методы научного анализа и статистической обработки полученных результатов
- Методы получения каротиноидов
- Биологические методы получения накопительных культур