ГЛАВА 22. ПАРВОВИРУСЫ
Парвовирусы (Parvoviridae)—это большая неоднородная группа вирусов, в которую входят вирусы, вызывающие разнообразные заболевания млекопитающих и птиц, сателлиты '(дефектные вирусы) аденовирусов, вирусы насекомых [Ананьев В.
А., 1982; Bachman P. et al., 1979; Matthews R., 1982]. Всех их объединяет то, что геномом является небольшая, длиной около 5000 нуклеотидов (1,5X IO6—2,0 X IO6), однони- тевая ДНК. Нередко в разных вирионах содержатся положительная и комплементарная ей нити ДНК. Поэтому при экстракции ее из вирионов может образовываться двунитевая структура.
Семейство парвовирусов включает 3 довольно разные группы вирусов: возбудители своеобразных заболеваний млекопитающих и птиц (род Parvovirus),дефектные сателлиты аденовирусов (род Depetidovirus)и вирусы денсонуклеоза насекомых (род Densovirus).Несмотря на экологическую разобщенность этих 3 групп вирусов, они имеют много общего как в строении вирионов, так и стратегии их генома.
Вирионы являются икосаэдрами, имеют диаметр 18—26 нм, 32 капсомера; диаметр капсомеров 3—4 нм. Вирионы имеют высокую плотность (1,34—1,42 г/мл) в связи с большим (до 20% массы) содержанием в них ДНК и небольшой коэффициент седиментации (110—120 3). Генетический материал представлен однонитевой линейной ДНК с молекулярной массой l,5?106—1,8?106, состоящей из 4675—5176 нуклеотидов. Первые два рода различаются между собой тем, что у первого ДНК в вирионах всегда имеют положительную полярность, тогда как у второго в вирионах могут содержаться плюс- или минус-нити. Кроме того, геномы этих двух групп, вирусов имеют и другие отличия, хотя Т-образные структуры, образуемые З'-концевыми последовательностями обеих групп, чрезвычайно сходны. Сходна и структура двух рамок считывания (неструктурные и структурные белки) у вирусов- сравниваемых групп.
Вирионы содержат три капсидных белка (VPl, VP2и VP3),молекулярная масса которых варьирует у разных вирусов.
По-видимому, синтез всех 3 структурных белков кодируется общей последовательностью генома и различия объясняются разными точками считывания и сплайсингом.После адсорбции, проникновения в ядра клеток и депротеинизации начинает функционировать вирусный геном, транскрипция которого обеспечивается клеточной РНК-полимера- зой, причем синтезируются белки (NSl, NS2или только первый из них), участвующие в репликации вирусной ДНК. У сателлитов аденовирусов в этом процессе участвуют и белки аденовируса. Предполагают, что З'-конец складывается, образуя шпильку, и функционирует как праймер синтеза ДНК, после чего происходят разрезание в области шпильки и разделение родительской или дочерней молекул ДНК. Механизмы транспортировки' структурных белков с мест синтеза в цитоплазме в ядро изучены мало, как и сборка вирионов. К роду Parvovirusотнесены 14 вирусов, вызывающих разнообразные заболевания грызунов, домашних животных, норок, кроликов, хомяков, гусей. Вирионные белки имеют молекулярную массу 81000—86000 (VPl), 64000—77 000 (VP2), 60 000—67000 (VP3).Вирусы реплицируются в ядрах интен
сивно делящихся клеток. Впрочем, при изучении парвовируса панлейкопении кошек-было показано, что он может реплицироваться в клетках с остановленной (блокированной) фазой S' [Lenghaus С. et al., 1985].
В репликации парвовирусов (Н1 вирус мышей) участвует некапсидный вирусспецифический белок NCVPl,который существует в двух формах — собственно NCVPl (84 000) и NCVPl' (92 000). Оба белка появляются на ранней стадии инфекции [Ron D., Tal J., 1984]. Парвовирусы нередко вызывают латентную инфекцию, длительную персистенцию. По антигенной структуре выделяют 4 группы вирусов, имеющих внутри групп иммунологическую общность. При заражении клеток Lлимфотропным парвовирусом мышей устанавливается персистенция, в ходе которой, с одной стороны, происходят мутации вируса, а с другой — мутации клеток, что типично для этого взаимоотношения двух партнеров и может быть определено как модель коэволюции [Ron D., Tal J., 1985].
Один из неструктурных белков парвовирусов весьма.консервативен, в том числе у автономно реплицирующихся парвовирусов и сателлитов аденовирусов, а также у парвовируса человека (В19) [Shade R. et al., 1986].
Парвовирус мыши (MVM) имеет гены для синтеза двух неструктурных белков: NSl (83 000) и NS2 (25000). Геном кодирует 3 транскрипционных единицы, которые совпадают, но обладают разными местами сплайсинга, в результате чего мРНК имеют молекулярную массу 48000, 3300 и 3000. PHK с массой 3000 кодирует синтез двух капсидных белков VPl (83 000) и VP2 (64 000). Третий структурный белок VP3 (62 000) образуется после протеолитического выщепления NH2-κoH∏eBθro участка VP2 [Cotmore S., Tattersall P., 1986].
При сравнении парвовируса собак с вирусом энтерита норок была выявлена 86% гомология их ДНК, что свидетельствует о близком родстве обоих вирусов [McMaster G. et al., 1981]. Парвовирус, вызывающий алеутскую болезнь норок (хроническое системное заболевание), имеет однонитевую ДНК с молекулярной массой l,4?106, седиментирующую при 16S. Два вирусных пептида, закодированные на ней, имеют молекулярную массу 89 100 и 77 600 [Bloom М. et al., 1980].
Парвовирусы млекопитающих (кошек, норок и собак) имеют выраженное антигенное родство, хотя вызывают разные болезни: у кошек — панлейкопению. У норок — энцефалопатию, у собак — энтерит й миокардит [Surleraux M., Burtonbay G., 1984]. Последний вирус индуцирует синтез 3 полипептидов (67 000, 70 000 и 85 000), а также, возможно, еще одного полипептида (50 000). Автономный парвовирус человека был обнаружен у здоровых доноров, а также у лиц с кореобразной сыпью, суставными болями, лейкопенией и другими
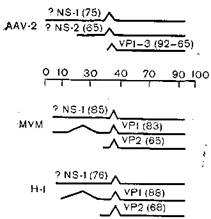
Рис.40. Транскрипция ДНК-вирусов AAV-2, MVMи H-I. Горизонтальные линии — вирусные транскрипты, наклонные линии — интроны; указаны области, кодирующие капсидные и неструктурные вирусные белки, в скобках — молекулярная масса белков (?103).
симптомами, включая вспышки инфекционной эритемы [Anderson M., Pattison J., 1984].
К роду Dependovirusотнесены сателлиты аденовирусов, выделенные вместе C аденовирусами человека, ■сельскохозяйственных животных и птиц. Молекулярная масса вирионных белков составляет 87 000 (VPl), 73 000 (VP2) и 62 000 ( VP3).
Три капсидных белка содержат совпадающие аминокислотные последовательности. Их синтез кодируется на правой половине генома и для каждой из них синтезируется своя мРНК [Janik J. et al., 1984] (рис. 40).
Вирусы размножаются лишь в присутствии вируса-помощника, которым в естественных условиях является соответствующий аденовирус. Репликация сателлитов аденовирусов обеспечивается ранними генами аденовирусов (la, Ib, 2а и 4), а также транскриптом малой РНК, синтезируемым клеточной РНК-полимеразой III [Richardson W., Westphal H., 1984]. В эксперименте показана возможность размножения их в присутствии герпесвирусов [Buller R. et а]., 1981].
При отсутствии аденовируса-хелпера репликации сателлита не происходит. Однако вирионы могут проникать в клетку, их ДНК может достичь ядра и там сохраниться, будучи интегрированной в клеточный геном. Освобождение ее может быть обеспечено вирусом-хелпером и стать началом продуктивной инфекции. Необходимо также отметить, что интеграция генома сателлита аденовируса не сопровождается трансформацией клетки и изменением ее фенотипа. В то время как аденовирус обеспечивает репликацию сателлита, последний при коинфекции угнетает как литический процесс, так и онкогенез, вызываемый аденовирусом. Вероятно, некоторые сателлиты аденовирусов, например ААВ4, могут поражать и человека, и домашних животных. Вся эта группа вирусов очень компактная и имеет общие «корни» происхождения.
Род Densovirusвключает 10 вирусов чешуекрылых насекомых, а также, возможно, двукрылых и прямокрылых. В от
личие от предыдущих родов представители этого рода имеют 4 вирионных белка: VPl (18000), VP2 (72 000), VP3 (57 000) и VP4 (46 000).
Вирусы размножаются преимущественно в личинках, накапливаясь в гипертрофированных ядрах.Одними из интересных особенностей 4 структурных белков вируса денсонуклеоза тутового шелкопряда являются их выраженная гомология и наличие антигенного родства. По-видимому, синтез всех этих белков кодируется общей последовательностью ДНК. Гомология имеется также между разными видами денсовирусов насекомых [Bando H. et al., 1984].
Репликация осуществляется при активном участии клеточной системы синтеза ДНК, поэтому вирусы развиваются преимущественно в делящихся клетках. При этом к ДНК ковалентно прикрепляется белок между тирозиновым остатком и 5'-концом молекулы ДНК- Этот белок не кодируется вирусом. [Chow М. et al., 1986].
Несмотря на сходство ряда свойств, парвовирусы все же не являются компактной группой обособленных вирусов, и объединение их в одно семейство может представляться несколько искусственным. Но, с другой стороны, эти вирусы и не являются случайной группировкой. Близкие размеры геномов, молекулярные массы структурных белков, тесная связь репликации с ядерной активностью клеток — все это объединяет вирусы теплокровных животных с вирусами насекомых.
Особенный интерес представляют дефектные вирусы.. В данном случае это компактная группа сателлитов аденовирусов. Вообще же наряду с ДНК-содержащими имеются и РНК-содержащие вирусы, паразитирующие в растениях и грибах. Типичным вирусом-сателлитом, размножающимся в растениях, является сателлит вируса некроза табака. В данном случае типы нуклеиновой кислоты сателлита и вируса- помощника совпадают. В случае с дельта-вирусом дефектный вирус является РНК-содержащим, а реплицирующий его вирус — ДНК-содержащим. Однако необходима поправка: последний вирус имеет репликативным интермедиатом РНК. Происхождение этих многочисленных и разнообразных вирусов и их эволюция требуют специального изучения.
Еще по теме ГЛАВА 22. ПАРВОВИРУСЫ:
- Семейство Парвовирусов (Family Parvoviridae)
- Глава 17 ВЫЧИСЛИТЕЛЬНЫЙ ЭКСПЕРИМЕНТ
- Система семейной поддержки (глава 19)
- Глава 1. Введение в иммунологию
- Глава 7. Этапы факоэмульсификации
- Встреча с Внутренним Наставником (глава 15)
- Глава 8. Болезни легких
- ГЛАВА 12 АНОМАЛИИ И ЗАБОЛЕВАНИЯ РОГОВИЦЫ
- ГЛАВА 6. ЭПИДЕМИОЛОГИЯ И ПРИНЦИПЫ ОРГАНИЗАЦИИ ПСИХИАТРИЧЕСКОЙ ПОМОЩИ
- Глава 6 Сопроводительная терапия в детской онкологии и гематологии
- ГЛАВА 6 ПАЛЬЦЕВАЯ ДЕРМАТОГЛИФИКА ПРИ ОГРАНИЧЕННЫХ ФИЗИЧЕСКИХ ВОЗМОЖНОСТЯХ
- Глава 3 КЛИНИКА, ДИАГНОСТИКА И ПРОГНОЗ АНДРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ У ДЕТЕЙ И ПОДРОСТКОВ