Высокая генотоксическая устойчивость сетчатки глаза
Полученные нами результаты по облучению и действию МНМ подтверждают тезис о высокой генотоксической устойчивости зрелой сетчатки мышей. Одна из предпосылок к этому - активная репарация ДНК после действия у- и протонного излучений в дозе 14 Гр (см.
рис 7). Репарация сочетается с увеличенной экспрессией Р53 и АТМ, которая нормализуется спустя 12 ч после облучения, времени завершения репарации индуцированных излучениями разрывов в ДНК (см. рис. 6).Заметные морфологические изменения в фоторецепторном слое сетчатки глаза отмечаются после облучения в дозе 25 Гр. Эти изменения выражаются в деградации наружных сегментов фоторецепторов сетчатки, а также в снижении плотности и толщины ядерного слоя фоторецепторов (см. рис. 9). Деградация нарастает во времени и связана с гибелью фоторецепторов, протекающей по механизму апоптоза. На апоптоз указывает возросшая экспрессия проапоптотических белков Р53, FasR и активной каспазы 3 (см. рис. 10). Все это в совокупности наблюдается и при действии МНМ на сетчатку глаза (см. рис. 11, 14). Высокая устойчивость сетчатки и активный механизм репарации ДНК указывают на существование генотоксического порога, который обуславливает нелинейный характер зависимости эффекта от дозы (Johnson et al., 2009) агентов. На пролиферирующих клетках генотоксический порог наблюдался при действии алкилирующих агентов (Jenkins et al., 2005). Он выражался в нелинейной зависимости эффекта от дозы и прямо связан с активной системой репарации ДНК.
Исследование генотоксического действия МНМ обнаружило две ранее не описанные особенности сетчатки глаза. Во-первых, высокую спонтанную поврежден- ность ДНК в сетчатке глаза мышей. По этому показателю органы мыши располагаются в порядке возрастания поврежденности ДНК в клетках - лимфоциты < клетки печени < клетки мозга < клетки сетчатки, который совпадает со степенью оксигенации тканей.
Средний момент хвоста ssДНК-комет интактной сетчатки, mt=32±2, соответствует дозе облучения протонами 15-20 Гр. Такая доза облучения неизбежно индуцирует заметное повреждение и dsДНК в сетчатке. Однако интак-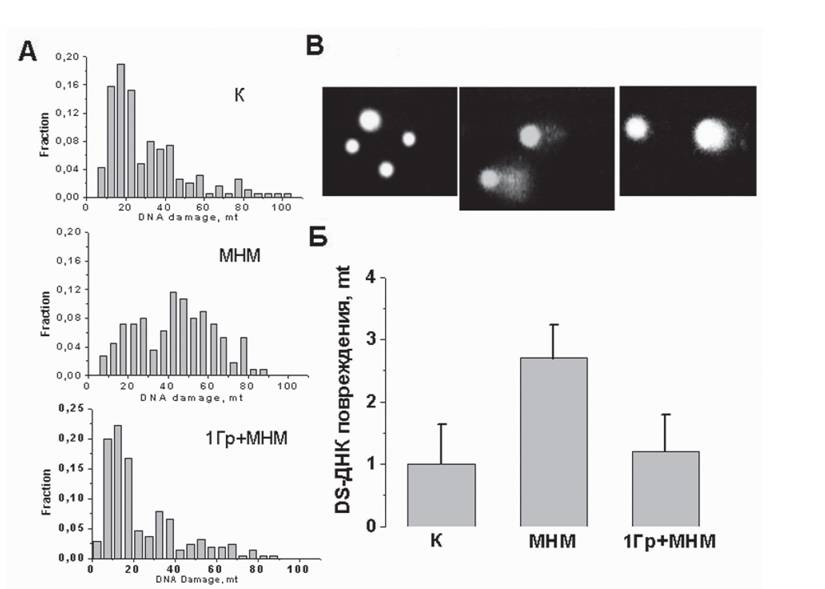
Рис. 16. Поврежденность двунитевой ДНК (сНДНЕС) в клетках сетчатки глаза спустя 48 ч после введения MHM 70 мг/кг и облучения протонами (1 ГрШНМ)
А - гистограммы распределений клеток по степени повреждениести сНДНК; Б - средние значения, найденные их этих распределений ±SD; со В - микрофотографии сНДНК-комет, доминирующие в каждом из распределений
тная сетчатка в нейтральных условиях, dsДНК, не обнаруживает разрывов, хотя форма комет, напоминающая одуванчики, указывает на декомпактизацию хроматина в клетке и на ослабление его связи с ядерным матриксом. Все это говорит о том, что спонтанные повреждения ДНК в сетчатке не являются разрывами, но трансформируются в таковые в щелочных условиях (рН>13), то есть являются ок- си-модифицированными основаниями ДНК.
Вторая обнаруженная нами особенность сетчатки - активная репарация, удаляющая большую часть повреждений ДНК, индуцированных облучением и МНМ, но не затрагивающая предсуществующие спонтанные повреждения ДНК (см. рис. 14). Спонтанные повреждения в виде однонитевых разрывов ДНК в различных органах мышей отмечались в работах (Valarde et al., 2000, 2001). Степень поврежденности ДНК нарастала в клетках в том же порядке, что и у нас: лейкоциты < клетки печени < клетки мозга (Wang et al., 2010). Таким образом, наличие таких нерепарируемых дефектов в ДНК интактной сетчатки глаза и снижение ее функций и жизнеспособности тоже говорит о высокой генотоксической устойчивости.
Апоптоз в ответ на повреждение ДНК в пролиферирующих клетках признается радикальным способом предотвратить формирование генетически нестабильного потомства клеток. В случае дифференцированных клеток не ясно, способны ли они подвергаться апоптозу в ответ на генотоксический стресс (Simonatto et al., 2007).
Известно, что в постмитотических клетках подавлены отдельные механизмы репарации. Так, терминально дифференцированные клетки не способны осуществлять репарацию по механизму гомологичной рекомбинации, ассоциированному с репликацией. Кроме того, в этих клетках регуляторно подавлена глобальная репарация генома, одна из разновидностей механизма NER (Walsh, Perlman, 1997). Вместе с тем, терминально дифференцированные клетки транскрипционно активны и нуждаются в поддержании целостности транскрибируемой части генома на протяжение жизни клетки. Эту функцию выполняет механизм репарации, ассоциированный с транскрипцией (Nouspikel, 2007). Принимая во внимание эти факты, можно считать большую часть глобального генома терминально дифференцированных клеток (в т.ч. и клеток сетчатки) некритичной для выполнения их специфических функций. Поэтому репарацией ДНК в этой части генома клетка может до какой-то степени пренебречь. В результате в ней могут накапливаться спонтанные повреждения без очевидных цитотоксических последствий. Спонтанная поврежденность ядерной ДНК в сетчатке отражает толерантность клеток к генотоксическому стрессу.Таким образом, наши результаты показывают, что, так же как для делящихся клеток, одной из причин толерантности постмитотических клеток сетчатки к повреждениям ДНК является репарация. Вторая причина их толерантности - уменьшение физического размера радиочувствительной мишени до размеров транскрибируемого локуса генома. Мы предполагаем, что решающая роль в трансформации изначально пермиссивных повреждений ДНК (разрывов после облучения и модифицированных оснований и АП-сайтов после действия МНМ) в цитотоксическое принадлежит локализованным в транскрибируемых сайтах молекулам топоизомеразы 2 (topo 2).
Известно, что регуляция генной экспрессии осуществляется путем структурной перестройки (remodeling) хроматина и формированием петель (Kouzarides, 2007; Kadauke, Blobel, 2009). Важная роль в этом принадлежит топоизомеразе topo 2, входящей в состав комплекса, осуществляющего модификацию (Varga-Weisz et al., 1997; Kuniaki Sano et al., 2008).
Тopo 2 локализована в транскрибируемой области, где изменяет топологию ДНК, делая в ДНК временный двойной разрыв и лигируя его в последующем, благодаря своим эндонуклеазной и лигазной функциям (Champoux, 2001). Активацию эндонуклеазной активности topo 2 могут осуществлять как индуцированные внешним воздействием (Sabourin, Osheroff, 2000), так и спонтанно возникающие повреждения ДНК (Liu, Gerson, 2004). В результате в транскрибируемых локусах формируются нерепарируемые двунитевые разрывы, приводящие к образованию высокомолекулярных (300-0,5 Mb) фрагментов ds,nHK (Taverna et al., 2001), являющихся ранним маркером апоптотической деградации генома (Tronov et al., 1999). В работе (Fishel et al., 2007) показано, что для инициации этих процессов необходимо попадание повреждения в область, перекрываемую эн- донуклеазной активностью topo 2. В противном случае повреждение может быть репарировано, не вызвав цитотоксического эффекта. Этот механизм объясняет как наличие генотоксического порога и вклад репарации в его формирование, так и, в конечном счете, процесс трансформации повреждения ДНК в цитотоксический эффект в постмитотических клетках сетчатки.4.
Еще по теме Высокая генотоксическая устойчивость сетчатки глаза:
- Повреждение и восстановление сетчатки у мышей после воздействия генотоксических агентов ионизирующей радиации и метилнитрозомочевины
- Функциональное и структурное восстановление сетчатки глаза после генотоксическоговоздействия
- Ответ сетчатки глаза на введение метилнитрозомочевины
- Ответ сетчатки глаза мышей на облучение
- Внутренняя (чувствительная) оболочка глаза (Tunica interna (sensoria) bulbi) — сетчатка (retina)
- 164. Кистовидная и кольцевидная дегенерация сетчатки. Лечение склеротической дегенерации сетчатки
- 189. Ранения глаза: общее понятие, классификация, причины, прогноз. Непроникающие ранения глаза
- Глава 2. Разработка математической электрофизической модели глаза для исследования гемодинамики переднего отдела глаза методом транспальпебральной реоофтальмографии
- 1. Показатели финансовой устойчивости
- Проблема устойчивости пневмококка к антибактериальной терапии