Образование конформационно модифицированных наноамилоидов с разной архитектурой
Не менее важным для нейронов является процесс постреакционного сворачивания фрагмента белковой молекулы Ар с участием системы шаперонов в конформа- ционную архитектуру нативной молекулы белка, которая только в таком состоянии может выполнять свои уникальные функции (Narayan et al., 2012).
В нормальных физиологических условиях все молекулы Ар при участии шаперонов приобретают физиологическую (нормальную) конформацию, выполняют свои функции и по принципу обратной связи поддерживают (на одном уровне) или активируют синтез АРР, а затем в ликворе и крови последовательно расщепляются протеолитическими ферментами (Ghiso et al., 2004).На основе экспериментального анализа пусковых факторов амилоидоза Ар мы считаем, что при сложившейся ситуации процесс биосинтеза белка выходит из-под контроля нейрона. В результате происходит повышение концентрации постреакционных фрагментов, а затем Ар-пептидов, которые стимулируют (по закону обратной связи) в нейроне синтез белка предшественника АРР, и последующему P-, у-процессингу подвергается большее количество АРР. Если процесс наращивания в нейроне синтеза АРР и количества Ар-пептидов перестал регулироваться, то это приводит к гиперконцентрации постреакционных Ар-фрагментов. Основная часть образованных Ар-фрагментов взаимодействует с системой шаперонов и приобретает физиологически нормальную конформацию Ар-пептидов. Однако физиологический уровень шаперонов в нейронах, необходимый для эффективного контроля и коррекции сворачивания образованных de novo Ар-фрагментов, не справляется с высокой концентрацией образующихся постреакционных фрагментов. Поэтому другая часть Ар-фрагментов, которая осталась «не охваченной шаперонами», приобретает конформацию, устойчивую к окружающей среде: Ар-фрагменты расплетаются частично или полностью (обратимая денатурация). Затем под влиянием взаимодействия аминокислотных остатков в расплетенных Ар-фрагментах, рН и ионного состава окружающей среды, концентрации фрагментов Ар в реакционном пространстве формируется вторичная структура, а затем образуются мономерные конформационно модифицированные пептиды - нано-Ар.
Известно, что кроме нейронов в некоторых других дифференцированных клетках при определенных условиях конформационно неустойчивые фрагменты или молекулы белков с небольшой молекулярной массой, растворимые в водной среде и выполняющие транспортные или регуляторные функции, могут приобретать необычную, ненативную конформацию. При взаимодействии молекул таких белков с диполями воды вокруг белковой молекулы формируется «крепкая» гидратная оболочка. В результате формируются наномасштабированные биочастицы, устойчивые к биохимическим и иммунологическим атакам клеток и организма. Образуется особый класс: наночастицы биологического происхождения - бионаночастицы. Для некоторых нейронов это событие может послужить пусковым фактором гибели нейронов от амилоидоза. Мономерные нано-Ар плохо растворимы в водных растворах, имеют в своем составе преимущественно Р-структурную конформацию и гидрофобные участки. При этом молекулы меняют свои свойства, они выходят из-под контроля нейронов и организма и становятся самостоятельными объектами нано-Ар (размером 1,2-1,5 и более нанометров), которые обладают патогенными функциями по отношению к нейронам. В определенной концентрации нано-Ар включают каскад событий, приводящий к амилоидозу нейронов и их гибели. Наиболее известными являются конформационно модифицированные Р-амилоиды и прионы - скрепи, которые обладают кроме способности к фибрилообразованию инфекционными свойствами (Prusiner, 1991) и могут передавать информацию о своей форме другим белкам (самовозобновляться) в организме у инфицированных животных - белковая наследственность (Prusiner, 1994, 1997). Эти объекты можно объединить как биогенные наночастицы, так как они имеют общие специфические особенности и закономерности: 1) без внешнего участия образуют олигомерные наночастицы больших размеров различной архитектуры: округлые, замкнутые, растворимые в биологических жидкостях (рис. 2а), и линейные (рис. 2б), плохо растворимые - предфибриллярной и фибриллярной (нерастворимые) структуры; 2) обладают пространственной памятью или «конформационной наследственностью»; 3) не гидролизуются эндогенными ферментами.
Анализ собственных и опубликованных результатов других исследователей показал, что в каскаде молекулярно-клеточных событий, ведущих к повреждению нервных клеток, развитию амилоидоза и патогенеза БА, участвуют наночастицы, образованные из конформационно модифицированных Ар и других белков, а также клеточные структуры с нарушенными функциями (Wall et al., 2011; Maltsev et al.,
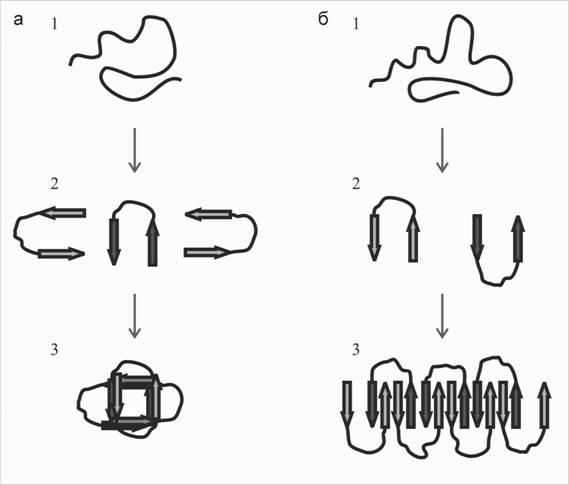
Рис. 2. Схема образования округлых олигомерных нано-р-амилоидов (а). Схема образования линейных олигомерных нано-р-амилоидов и протофибрилл (б)
1 - частично или полностью развернутый фрагмент р-амилоида после процессинга АРР у-секретазой; 2 - образование конформационно модифицированных р-амилоидов - мономерных нано-Ар; 3 - олигомеры, состоящие из мономерных нано-Ар
2011). Их участие в развитии амилоидоза нейронов и патогенеза нейродегенеративных заболеваний (НДЗ) осуществляется посредством активации амилоидогенных ферментных систем, конформационных перестроек и межмолекулярных взаимодействий между различными наночастицами некоторых конформационно модифицированных маркерных белков, характерных для НДЗ.
Склонность к конформационной модификации таких специфических белков проявляется при наличии хотя бы одного из условий: 1. Повышение локальной концентрации Ар вследствие спорадически или генетически обусловленного избытка синтеза или усиления процессинга предшественника (АРР). 2. Мутации, вызывающие аминокислотные замены в пептидной цепи низкомолекулярных белков, которые увеличивают их склонность к агрегации. 3. Функциональная недостаточность в нейронах системы шаперонов и механизма убиквитин-протеасомной деградации белков с аномальной конформацией.
Таким образом, начинается процесс амилоидоза. Локальное образование высокой концентрации гидрофобных мономерных нано-Ap рядом с мембраной нейрона вносит коррективы в основные функции мембраны: полупроницаемость, избирательность в поглощении веществ из среды, осмотическую стабильность и способность генерировать электрические потенциалы (влияя на транспорт ионов Са2+).
Благодаря изменению электронной плотности на функциональных группах нано- Ap изменяется характер их взаимодействия с водой, с ионами и другими веществами. В одном состоянии карбоксильные группы нано-Ap селективно связывают ионы K+, в другом - Na+ (Линг, 2008) и ионы других металлов. Нано-Ар активно связываются с двухвалентными металлами, которые участвуют в окислительном стрессе нейронов. При этом следует отметить, что многие работы указывают на то, что ионы Zn2+ ускоряют образование амилоидных фибрилл, так как уменьшается суммарный заряд нано-Ар, увеличивается его гидрофобность и агрегаци- онные свойства (Talmard et al., 2009). Также было показано, что ионы Zn2+ способствуют агрегации Ар, когда пептид из а-спиральной конформации переходит в Р-амилоидную структуру (Chen et al., 2006; Talmard et al., 2009). Также была изучена реакция образования координационной связи с ионами меди Cu2+ для фрагмента Ap и было показано методом 1H ЯМР и методом спектроскопии, что His6, His13, His14, Tyr10 образуют координационные связи с ионом меди (Ma et al., 2006; Minicozzi et al., 2008). Эти события угнетают функциональное состояние нейронов, которое характеризуется изменением ряда параметров гомеостаза нейронов, их индукцией к дедифференцировке нейронов и вхождение в аберрантный, некоординированный, летальный цикл в стадии G-2 (Гуляева, 2005). В первую очередь происходит резкое уменьшение синаптических контактов и уменьшение синапсов у нейронов (Singh et al., 2001).Это приводит к образованию такого большого количества фрагментов молекул Ар, при котором система сворачивания молекул Ар в физиологическую конформацию с помощью шаперонов не справляется со своей задачей. Часть молекул Ар минует встречу с шаперонами, оказывается в «свободном плавании», и сворачивание пептида происходит не по клеточному сценарию, а «диким» образом, по законам термодинамики и биохимии, в зависимости от состава окружающей среды, температуры и аминокислотного состава белковой молекулы.
Экспериментально показано, что в условиях обратимой денатурации белковая молекула в зависимости от состава окружающей среды (инкубационной среды) и температуры может принимать широкий спектр кратковременных и устойчивых конформационных состояний (Maltsev et al., 2011). Таким образом, образуются конформационно модифицированные «молекулы-частицы наноамилоиды» (нано-АР). В структуре у этих нано-Ар преобладают гидрофобные участки. В связи с этим они плохо растворимы в водных растворах и как следствие - обладают высокой агрессивностью к взаимодействию с другими молекулами, органеллами и мембранами, и предрасположены к образованию олигомеров нано-Ар (Maltsev et al., 2011) с разной архитектурой. Они не контролируются нейронами и организмом, и с полным основанием их можно считать частицами нано-Ар (Smyth et al., 1994). Находясь рядом с мембранами нейронов, нано-Ар подавляют их физиологические функции, ингибируют работу синапсов и мембранных каналов, угнетают активность митохондрий и других внутриклеточных систем, способствуют развитию окислительного стресса (Wilkinson, 2001). Эта ситуация может привести к амилоидозу нейрона, а при условии отсутствия периода покоя, в котором происходит восстановление функций всех клеточных систем, не исключена и гибель нейрона (Бехтерева, 1988). Плохо растворимые мономеры нано-Ар обладают высокой активностью к образованию олигомеров нано-Ар разной архитектуры (Maltsev et al., 2011). После образования олигомеров нано-Ар их агрессивность по отношению к окружающим молекулам, органеллам и клеткам пропадает. Олигомеры нано-Ар фибриллярной конфигурации (архитектуры) нерастворимы и образуют предфибриллярные образования, а затем из них формируются синильные бляшки и фибриллы. Олигомеры нано-Ар с округлой (шаровидной) архитектурой растворимы в водной среде и мигрируют в ликвор, а затем в кровь (Maltsev et al., 2011).Экспериментально показано, что разрушенные аксоны, особенно в острой стадии, могут служить источниками резко повышенного локально уровня АРР, что неминуемо сопровождается отщеплением из него Ар-пептида (Smith et al., 2003). Однако существуют данные, что АРР может играть нейропротекторную и нейротрофическую роль, а также защищать нейроны от эксайтотоксичности (Choi et al., 1995; Furukawa et al., 1996; Mucke et al., 1996).
8.
Еще по теме Образование конформационно модифицированных наноамилоидов с разной архитектурой:
- Какова логика построения ПК, его архитектура?
- АРХИТЕКТУРА И ПЛАНИРОВКА ЗДАНИЙ, БЛОКОВ, СЕКЦИЙ
- Основные компоненты архитектуры клетки