Новые лиганды Cys-петельных рецепторов и путь к лекарствам
Необходимость детекции и воздействия на отдельные подтипы нАХР определяет необходимость поиска и создания новых высокоаффинных и селективных лигандов. Основой служат многочисленные известные соединения, взаимодействующие с разными участками нАХР: агонисты, конкурентные антагонисты, блокаторы ионного канала, а также разнообразные модуляторы холинергической активности.
Такое разнообразие, то есть способность связывать различные классы соединений, служит вероятным объяснением участия нАХР в самых различных физиологических процессах. Хорошо известно, что связывание агонистов с ортостерическими участками рецептора приводит к серии его конформационных изменений, которые в свою очередь вызывают открытие ионного канала. Конкурентные антагонисты (включая белковые и пептидные нейротоксины) тоже взаимодействуют с участком связывания агонистов, но при этом предотвращают активацию рецептора. На холинорецепторах выявлены и другие участки взаимодействия с рядом соединений, которые вызовут ослабление или усиление ионных токов. Последние, так называемые позитивные аллостерические модуляторы (PAM), в настоящее время представляют большой интерес как возможные лекарственные средства нового поколения. Перспективными мишенями для лекарственных препаратов могут оказаться нАХР, содержащие а7-субъединицы. Показано, что введение селективного агониста а7 нАХР оказывало нейропротекторный эффект у крыс с моделью паркинсонизма, индуцированной 6-гидроксидофамином (Suzuki et al., 2013). Новый класс соединений для лечения нейродегенеративных заболеваний могут представлять также аллостерические модуляторы, усиливающие ответы а7- и а4р2 подтипов нейрональных нАХР или частичные агонисты нАХР (Weltzin, Schulte, 2010; Henderson et al., 2011). К последним может быть отнесен варениклин - препарат, который сравнительно недавно стал использоваться в качестве средства, помогающего избавиться от никотиновой зависимости (Turner et al., 2010).Как уже отмечалось, ранее наиболее востребованными инструментами изучения структурно-функциональных свойств нАХР служили а-нейротоксины из ядов змей, благодаря своему исключительно высокому сродству к некоторым подтипам рецептора. Однако не для всех нАХР имеются избирательно действующие на них нейротоксины змей. Так называемые «короткие» a-нейротоксины эффективно взаимодействуют только с мышечными нАХР; а-нейротоксины «длинного» типа узнают кроме мышечных также инейрональные гомоолигомерные (а7, а8, а9) холинорецепторы (см. обзор Tsetlin, 1999). Еще один токсический полипептид из яда B. multicinctus - к-бунгаротоксин - специфичен к гетероолигомерному а3р2 нАХР. а-конотоксины, пептиды из яда морских улиток семейства Conus, являются конкурентными антагонистами нАХР и имеют высокую избирательность действия на различные подтипы холинорецепторов (см. обзоры: Terlau, Olivera, 2004; Han et al., 2008; Tsetlin, Hucho, 2009).
Хотя природа дала нам большое количество соединений, различающих тот или иной тип Cys-петельных рецепторов, задачи поиска и конструирования новых представителей, которые с большей избирательностью помогали бы идентифицировать, а также активировать или дезактивировать интересующий подтип определенного (например нАХР) рецептора, остаются по-прежнему актуальными. В связи с важной ролью а7 нАХР много усилий было потрачено на то, чтобы модифицировать природные а-конотоксины и получить аналоги, пригодные для идентификации этих рецепторов в тканях. Это недавно удалось американским исследователям: на основе природного а-конотоксина ArIB были получены его аналоги с несколькими заменами, несущими радиоактивные и флуоресцентные метки, вполне пригодные для надежного детектирования а7 нАХР (Hone et al., 2010). Нам удалось на основе а-конотоксина PnIA получить аналог, имеющий очень высокое (0,5 nM) сродство к АХСБ L. stagnalis, а его радиоиодированное производное очень удобно для скрининга соединений с возможной холинергической активностью (Kasheverov et al., 2011).
Выход на новые природные соединения, которые могли бы сыграть роль инструментов в изучении рецепторов, сегодня имеется благодаря протеомным исследованиям ядов.
Так, из яда кобры Naja kaouthia нами выделен димерный а-кобратоксин (Osipov et al., 2008), в котором две молекулы а-кобратоксина соединены двумя межмолекулярными дисульфидными связями, образовавшимися за счет межмолекулярной перегруппировки дисульфидных связей, имевшихся в исходном а-кобратоксине (рис. 2). Вывод о пространственной структуре этого нового нейротоксина и расположении дисульфидных связей был нами сделан на основании установления его кристаллической структуры (Osipov et al., 2012). Уже давно был известен к-бунгаротоксин, в котором две молекулы, сходные с а-бунгаротоксином, образуют прочно связанный димер, не содержащий каких-либо межмолекулярных ковалентных связей. к-Бунгаротоксин имеет высокое сродство к гетеромерному нейрональному а3р2 нАХР, тогда как а-бунгаротоксин или а-кобратоксин блокируют только мышечный и а7 нАХР. Обнаруженный нами димерный а-кобратоксин сохранил эти исходные активности, но и приобрел способность к блокированию а3р2 нАХР (Osipov et al., 2008, 2012).До сих пор мы рассматривали выделенные из ядов нейроактивные белки и пептиды, мишенью для которых служат различные нАХР. Чуть более 10 лет назад американскими исследователями в мозге мыши был обнаружен ген, кодирующий белок Lynxl, предположительно имеющий ту же трехпетельную структуру, что и а-нейротоксины змей (Miwa et al., 1999). Структура гена указывает, что эндогенный белок млекопитающих Lynx1 должен иметь и существенное отличие от нейротоксинов - дополнительный гликозил-фосфатидил-инозитольный (GPI) якорь. С помощью этого якоря Lynx1 прикреплен в мозге к мембране в непосредственной бли-
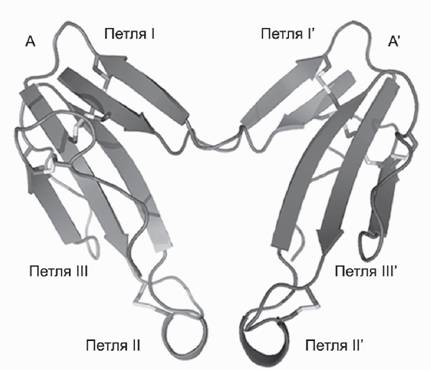
Рис 2. Кристаллическая структура димерного а-кобратоксина
Показаны исходные мономеры (А и А') а-кобратоксина. I-III - обозначения петель, имеющих Р-структурную конформацию. Две межмолекулярные дисульфидные связи образованы остатком Cys 3 и Cys 20 одного мономера с остатками Cys 20 и Cys 3 соседнего
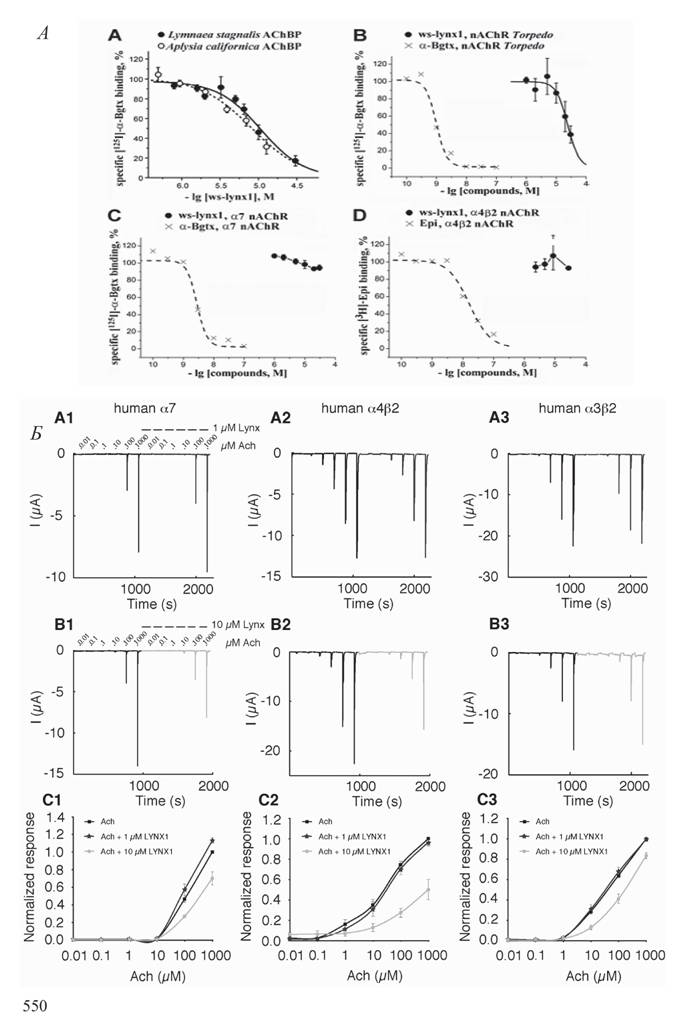
Рис.
3. Взаимодействие ws-lynx1 с никотиновым ацетилхолиновым рецептором А - конкуренция ws-lynx1 с иодированным а-бунгаротоксином за связывание с АХСБ и мембранно-связанным нАХР Torpedo californica и отсутствие конкуренции за связывание с а7 нАХР. Б - снижение амплитуды тока в а7 нАХР, гетерологически экмпрессированных в ооцитах Xenopus, при добавлении 10 цМ ws-lynx1зости от никотиновых рецепторов и модулирует их функциональную активность. До недавнего времени Lynxi был недоступен в форме индивидуального белка. Поэтому все важнейшие исследования его функциональной активности, опубликованные в таких ведущих журналах, как Neuron и Science (см., например, Miwa et al., 2006; Morishita et al., 2010), были выполнены исключительно с использованием суперэкспрессии или нокаута гена Lynx 1. Информации о том, каков механизм взаимодействия Lynxi с нАХР, не имелось, а прогресс в этой области наметился, когда нам в 2011 г., благодаря экспрессии в E. coli и последующей очистке, удалось получить в виде индивидуального водорастворимого белка полноразмерный Lynx 1, не имеющий GPI якоря (ws-lynx1) (Lyukmanova et al., 2011). В результате впервые были установлены концентрации, при которыхможно в опытах in vitro зарегистрировать эффекты ws-lynx1 на различные подтипы нАХР. При этом было впервые показано различие в характере связывания ws-lynx1 с двумя основными типами нАХР: ws-lynx1 конкурировал с радиоактивным а-бунгаротоксином за связывание с рецептором мышечного типа, но не конкурировал за связывание с нейрональным а7 нАХР (рис. 3а). Однако электрофизиологические опыты выявили ингибирование токов в а7 нАХР при добавлении 10 mM ws-lynx1 (рис. 3б).
4.
Еще по теме Новые лиганды Cys-петельных рецепторов и путь к лекарствам:
- 3. Нейролипины - лиганды каннабиноидных рецепторов
- Зрительный путь и путь зрачкового рефлекса
- 6.лиганд-рецепторн взаимод.вторич посред.
- Никотиновые ацетилхолиновые рецепторы: физиологические функции и роль в нейродегенеративных процессах
- Корково-спинальный (пирамидный) путь
- Путь к выздоровлению
- 2.1.1.3 Toll-рецепторы
- 3. Экстрапирамидный путь через мозжечок.
- 7.5. НОВЫЕ ТЕХНОЛОГИИ:
- 3. Путь биосинтеза рибофлавина и его регуляция
- 3.1.2. Путь в психологию
- АА — не единственный путь
- Тектоспинальный путь
- Вестибулоспинальный путь
- Ретикулоспинальный путь
- 1. Экстрапирамидный путь через Монаковский пучок.
- Метаботропные рецепторы наружной мембраны клетки
- 77. Развитие личности и её жизненный путь.
- 80. Самосознание личности и её жизненный путь.