3. Экстрапирамидный путь через мозжечок.
Он состоит из шести нейронов — одного периферического и пяти центральных.
Первый центральный нейрон берет начало в двух участках мозговой коры — в двигательной зоне и в лобных долях.
Оба эти пучка идут через внутреннюю капсулу, а дальше через ножку мозга в Варолиев мост. Оканчиваются они в серых массах Варолиева моста на той же стороне.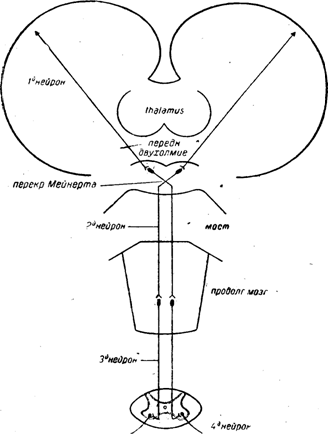
Рис. 5. Экстрапирамидный путь через переднее двухолмие.
Второй нейрон начинается в этих серых массах, переходит через среднюю линию (образует перекрест) и .в составе так называемой средней ножки .мозжечка (crus cerebelli ad pontem Varolii) идет в мозжечок, где оканчивается в коре и в flocculus главным образом на стороне, противоположной тому мозговому полушарию, откуда вышел первый нейрон.
Третий нейрон,весь лежит в пределах мозжечка. Он начинается в коре и в flocculus, а оканчивается в nucleus dentatus — все это в пределах одного и того же полушария.
Четвертый нейрон начинается в nucleus dentatus и через верхнюю мозжечковую ножку идет к красному ядру противоположной стороны, совершая таким образом второй перекрест.
Пятый нейрон — это уже известный пучок Монакова. Так как он делает так называемый перекрест Фореля, то вся система в целом в пределах пятого нейрона совершает третий перекрест.
В результате этих трех перекрестов двигательный импульс, пробегающий по этой системе, совершает полный винтовой оборот по длинной оси центральной нервной системы и в конечном счете иннервирует противоположную сторону. Шестой нейрон — периферический (рис, 6).
В то время как сведения наши о прямом кортико-мускулярном пути — в анатомической их части — представляют прочно сложившуюся систему, в учении об экстрапирамидных путях имеется много пробелов, и достоверность отдельных частей его далеко не одинакова. Одни звенья этих сложных и запутанных путей установлены вполне точно и даже изучены сравнительно-анатомически (как, например, пучок Монакова), другие представляют образования наполовину гипотетические.
Соединение этих звеньев в одну цепь различными авторами производится по-разному. Примером может служить экстрапирамидный путь через мозжечок: Левандовский (Lewandowsky) проводит второй нейрон в мозжечок, после чего следует дальнейшая цепь нейронов, обвивающая винтообразно длинную ось мозгового ствола; Бехтерев направляет импульс на отростки мостовых ядер прямо вниз через так называемый отвесный пучок моста в спинной мозг.
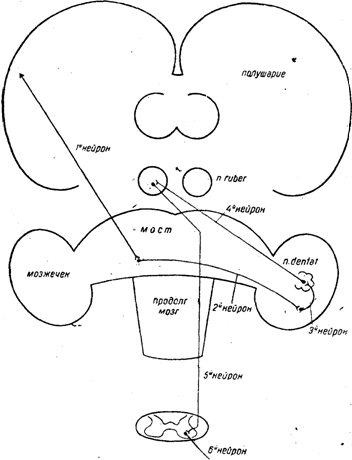
Рис. 6. Экстрапирамидный путь через мозжечок.
Соответственно этому отношение невропатологов к различным экстрапирамидным путям далеко не одинаково: одни готовы считать приведенное выше описание за прочно установленные анатомические факты, другие относятся ко всем этим концепциям с большой сдержанностью.
Непрочности анатомических основ вопроса соответствует и то смутное представление о физиологии экстрапирамидных путей, которым мы сейчас располагаем. Некоторые авторы, склонные приписывать экстрапирамидным путям важное функциональное значение, видят его в выполнении быстрых бессознательных движений защитного типа, — например, внезапная остановка на ходу перед препятствием — или в выполнении движений хотя и произвольных, но бессознательных, — как, например, ходьба, бег, играна музыкальных инструментах и т. п.
Мне лично представляется очень вероятным отношение экстрапирамидных путей к симпатической нервной системе. Есть основания думать, что эти прерывистые пути играют для периферического аппарата симпатической системы такую же роль, как пирамидный путь для периферического нейрона кортико-мускулярного пути. Более, подробную мотивировку этого мнения вы услышите в очерке строения симпатической системы.
Если непрочно установлены анатомические основания вопроса и смутны относящиеся сюда физиологические представления, то патология разбираемых систем может быть только гадательной.
И действительно, повреждению экстрапирамидных систем приписывались и параличи какого-то типа, промежуточного между центральными и периферическими, и различные насильственные движения, и многое другое.
Если бы вы спросили, как правильнее всего отнестись ко всем этим не вполне установленным и не приведенным в систему данным, то я бы формулировал свое мнение так:
Что кроме пирамидного пути существуют еще другие центробежные системы, — это сомнению подлежать не может. Они устанавливаются частью патологоанатомическими наблюдениями, частью опытами над животными. Но относительно функции их точно высказаться в настоящее время очень трудно: здесь возможны только более или менее правдоподобные предположения.
Последнее, на что я хочу обратить ваше внимание, — это вопрос об участии симпатической системы в иннервации поперечно-полосатых мышц.
Самый факт прохождения симпатических волоконец в мышцы был известен сравнительно давно, но раньше думали, что они идут к сосудам, что это волокна вазомоторной природы. И только за последнее десятилетие неожиданно выяснился их истинный характер благодаря остроумному сочетанию физиологического эксперимента и новейших гистологических методов.
У животных перерезались передние двигательные корешки, вследствие чего периферический двигательный нейрон перерождался вплоть до своего концевого аппарата в мышце, удалялись также и межпозвоночные ганглии, вследствие чего гибли чувствующие волокна. Тогда при исследовании соответствующих мышц можно было видеть наряду с перерождающимися нервными волокнами и их концевыми приборами — целые и неперерожденные безмякотные волокна также с концевыми приборами, оканчивающимися под сарколеммой мышечных волокон Смысл этих анатомических картин проверялся опытом. Когда перерезанные корешки успевали совершенно переродиться, пробовали раздражать их электрическим током и не получали никакого сокращения. Раздражение же соответствующих симпатических волокон давало мышечное сокращение очень своеобразного характера Выяснилось, между прочим, любопытное явление, что разные мышцы получают различное количество симпатических волокон. tàê, например, диафрагма и межреберные мышцы получают симпатических волокон даже значительно больше, чем соматические мышцы, заведуемые от n.
ulnans и п medianus, получают очень мало симпатических волокон, а разгибатели, снабжаемые от radialis, гораздо больше. Довольно много симпатических волокон получают мышцы едины и тазового пояса, т. е. невидимому именно те, которые создают вертикальное положение человеческого тела, делают, так сказать, из человека двуногое существо, а не четвероногое.Делались попытки выяснить физиологическую роль этих волокон. Так, например, думали присвоить им значение в поддержании мышечного тонуса. Но экспериментальная проверка не подтвердила этого предположения: удаление чуть ли не всей симпатической системы на одной стороне не создавало разницы в тонусе мышц на обеих, половинах тела
Есть попытки связать эти волокна с химизмом работающей мышцы. Пока еще определенных результатов получить не удалось.
Один только интересный результат получился из наблюдения над характером сокращения поперечно-полосатых мышц при раздражении симпатических волокон.
Так, у животных разрушался подъязычный нерв, снабжающий мыщцы языка. Когда этот нерв перерождался, то оставались целыми только симпатические двигательные волокна, которые идут к мышцам в составе чувствующего n. lingualis. Раздражение этого нерва, т. е. раздражение симпатических волокон, давало «своеобразное, медленное, долго длящееся тоническое сокращение парализованной половины языка».
Оказанным исчерпывается то немногое, что известно в настоящее время относительно физиологии этою иннервационного аппарата для поперечно-полосатых мышц.
Значение этих находок последних лет и чрезвычайно важно и вместе с тем чрезвычайно темно. Из них явствует, что так называемые «произвольные» мышечные волокна получают одновременно иннервацию из двух источников: цереброспинальной нервной системы, которой по ходячим воззрениям приписываются сознательно-произвольные функции, и симпатической, которой тот же источник присваивает непроизвольные функции. Следовательно механизм сокращения поперечно-полосатых мышц или, как принято говорить, «физиология произвольных движений», не покрывается механикой кортико-мускулярного пути: требуется еще какое-то дополнение.
И патология движения, которая в ее современном виде является почти исключительно патологией цереброспинальной, или, точнее, соматической двигательной сферы, также очевидно должна быть чем-то дополнена. С этой точки зрения всё учение о физиологии и патологии движения в том виде, как я его изложил в предыдущей лекции, приходится рассматривать в своеобразной исторической перспективе. Эта система постепенно создавалась в течение многих лет и, казалось, покрывала собой действительность. Но на наших глазах появились уже факты, которые не укладываются в ее рамки; очевидно эту систему придется скоро перестраивать. Начало перестройки происходит на наших глазах, а конец ее придется увидеть, вероятно, только вам. В этом вопросе мы присутствуем при глубоко интересном явлении, живем в переходный момент, когда еще не пришло новое, но уже наметился уход старого. Вас не должна смущать та ломка воззрений, которая неизбежно связана с такими периодами в истории науки. В этом случае следует помнить остроумный совет Шарко, который рекомендовал поступать с теориями так, как это делается с сорочками: мы пользуемся ими до тех пор, пока они свежи и новы; а когда они сомнутся, мы меняем их.
Еще по теме 3. Экстрапирамидный путь через мозжечок.:
- 1. Экстрапирамидный путь через Монаковский пучок.
- 2. Экстрапирамидный путь через переднее двухолмие.
- Зрительный путь и путь зрачкового рефлекса
- МОЗЖЕЧОК
- 1.4.4. Мозжечок
- 3.2. Экстрапирамидная система
- Характеристика двигательных реакций, наступающих по пирамидным и экстрапирамидным путям.
- МОЗЖЕЧОК
- КРОВОИЗЛИЯНИЕ В МОЗЖЕЧОК. HAEMORRHAGIA CEREBELLI. клиническая картина.
- 24.2. Пирамидные и экстрапирамидные дегенерации
- Ретикулоспинальный путь
- Синдромы поражения экстрапирамидной системы
- Тектоспинальный путь
- Корково-спинальный (пирамидный) путь
- Двигательные расстройства, связанные с нарушением функций пирамидной и экстрапирамидной систем.
- Путь к выздоровлению
- Использование компьютерной электромиографии для ранней и дифференциальной диагностики экстрапирамидных заболеваний
- 3. Путь биосинтеза рибофлавина и его регуляция
- 3.1.2. Путь в психологию
- АА — не единственный путь